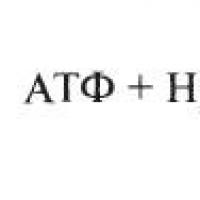Metabolismo at pag-andar ng carbohydrates. Anaerobic at aerobic oxidation ng glucose. Anaerobic oxidation ng carbohydrates Mga teknolohikal na katangian ng glucose oxidation reaction
Ang anaerobic oxidation ng carbohydrates ay nangyayari sa mga selula, organo at tisyu nang walang partisipasyon ng oxygen. Kung ang proseso ay nagsisimula sa conversion ng glucose at nagtatapos sa pagbuo ng lactic acid, kung gayon ito ay tinatawag na anaerobic glycolysis, kung ito ay nagsisimula sa conversion ng glycogen - glycogenolysis.
Glycolysis
Sa unang yugto ng glycolysis (Larawan 7), ang phosphorylation ng glucose ay nangyayari sa ilalim ng pagkilos ng enzyme. hexokinase sa pagkakaroon ng ATP at Mg 2+ ions na may pagbuo glucose-6-phosphate(G-6-F), na sa ika-2 yugto ay nagiging fructose 6-phosphate(F-6-F). Ang reaksyong ito ay catalyzed glucose phosphate isomerase . Sa ika-3 yugto, ang fructose-6-phosphate ay phosphorylated, na nagreresulta sa pagbuo fructose 1,6-bisphosphate(F-1,6-F). Ang catalysis ng reaksyong ito ay sinisiguro sa pagkakaroon ng Mg 2+ ions at ATPphosphofructokinase .
Sa ika-4 na yugto F-1,6-F sa ilalim ng impluwensya mga aldolase nahahati sa 2 phosphotriose - glyceraldehyde-3-phosphate(GA-3-F) at dihydroxyacetone phosphate(DAF).
DAP sa ilalim ng impluwensya triosephosphate isomerase nagiging GA-3-F. Kaya, mula sa isang molekula ng glucose ay nakakakuha tayo ng 2 molekula ng GA-3-P.
GA-3-F sa presensya glyceraldehyde phosphate dehydrogenase , coenzyme NAD + at phosphoric acid ay na-oxidized upang mabuo 1,3-diphosphoglycerate(1,3-DPG) at NADH(H+). Ang enerhiya na inilabas sa reaksyong ito ay naipon sa mataas na enerhiya na bono ng 1,3-DPG, na kung saan ay karagdagang na-convert sa 3-phosphoglycerate Sa ilalim ng impluwensiya phosphoglycerate kinase . Ang prosesong ito ay nauugnay sa phosphorylation ng ADP (formation ATP sa antas substrate phosphorylation).
3-FY kasama ang paglahok phosphoglycerate mutase nagiging 2-phosphoglycerate(2-PG), na sumasailalim sa dehydration sa presensya mga enolase at Mg 2+ ion. Ang resulta, phosphoenolpyruvate(PEP), na naglalaman ng high-energy bond. Ang PEP sa ilalim ng pagkilos ng pyruvate kinase sa presensya ng Mg 2+ ions ay na-convert sa enol form pyruvate. Ang prosesong ito ay nauugnay sa ATP synthesis sa antas ng substrate phosphorylation. Ang enol na anyo ng pyruvate tautomerizes sa ketone form.
Sa huling yugto ng glycolysis, ang pyruvate ay nabawasan sa L-lactate sa pagkakaroon ng NADH(H+) at enzyme lactate dehydrogenase . Ang coenzyme NAD + na nabuo sa reaksyong ito ay kasunod na ginagamit sa reaksyon ng oksihenasyon ng GA-3-P.
Kaya, ang mga huling produkto ng glycolysis ay 2 molekula ng L-lactate at 2 molekula ng ATP bawat 1 molekula ng glucose.
Regulasyon Ang glycolysis ay nangyayari sa antas phosphofructokinase mga reaksyon. Ang enzyme ay inhibited ng mataas na antas ng ATP at citrate. Ang mga allosteric activator ng phosphofructokinase ay AMP, ADP, P-6-P.
Glycogenolysis
Ang Glycogen ay isang branched polysaccharide na binubuo ng α-D-glucose residues na naka-link sa isa't isa sa mga linear na rehiyon ng molekula sa pamamagitan ng α-1,4-glycosidic bond, at sa mga branch point ng α-1,6-glycosidic bond.
Sa ilalim ng impluwensiya glycogen phosphorylase Ang cleavage ng isang glucose residue ay nangyayari sa mga linear na seksyon kasama ang paglipat nito sa isang phosphoric acid molecule, na nagreresulta sa pagbuo glucose-1-phosphate(G-1-F). Gumagana ang glycogen phosphorylase hanggang may 4 na residue ng glucose na natitira bago ang pinakamalapit na branch point, pagkatapos ay nagsimulang gumana ang enzyme. oligosaccharide transferase , naglilipat ng isang fragment ng 3 glucose residue sa katabing sangay, kaya nag-iiwan ng isang glucose residue sa branch point. Ito ay na-cleaved hydrolytically sa tulong ng -1,6-glycosidase sa anyo ng isang libreng molekula ng glucose, at ang mga kondisyon ay nilikha muli para sa glycogen phosphorylase upang gumana sa isang linear chain.
Produkto ng reaksyon ng glycogen phosphorylase G-1-F lalo pang nasa ilalim ng impluwensya phosphoglucomutase nagiging G-6-F, na kasangkot sa glycolytic landas (Larawan 8).

kanin. 8. Scheme ng pagkasira ng glycogen sa mga kalamnan at atay.
Ang mga huling produkto ng glycogenolysis ay 2 molekula ng L-lactate at 3 molekula ng ATP bawat 1 molekula ng glucose.
100 RUR bonus para sa unang order
Piliin ang uri ng trabaho Diploma work Course work Abstract Master's thesis Practice report Article Report Review Test work Monograph Problem solving Business plan Mga sagot sa mga tanong Creative work Essay Drawing Essays Translation Presentations Pagta-type Iba pa Pagpaparami ng uniqueness ng text Master's thesis Laboratory work On-line help
Alamin ang presyo
Ang paghinga sa mga mikrobyo ay itinuturing bilang isang proseso ng enerhiya o bilang isang hanay ng iba't ibang mga reaksiyong kemikal, oksihenasyon at pagkasira ng mga sangkap ng organic at inorganic na pinagmulan. Bilang resulta ng mga reaksiyong kemikal na ito, ang enerhiya ay inilabas, na ginagamit ng mga mikrobyo para sa pagsipsip ng mga sustansya, ang synthesis ng mga protina sa kanilang katawan, paggalaw, paglaki, pagpaparami at iba pang mga pag-andar ng buhay na organismo.
Ang isang halimbawa ng paglabas ng enerhiya ay ang oksihenasyon ng glucose, na maaaring ipahayag ng sumusunod na tambalan:
C6 H2O + 6 O 2 6 H2O+ 6CO2 + 674 kcal.
Tulad ng makikita mula sa mga equation, bilang resulta ng kumpletong oksihenasyon ng isang molekula ng glucose sa mga huling produkto (tubig at carbon dioxide), 674 malalaking calorie ng init ang pinakawalan.
Ang masiglang proseso ng paghinga sa mga mikrobyo ay mas kumplikado at depende sa likas na katangian ng materyal na nakapagpapalusog na ginamit.
Batay sa uri ng paghinga, ang mga mikrobyo ay nahahati sa aerobes at anaerobes; may mga mikrobyo na may transisyonal na uri ng paghinga.
Maaaring mabuhay at umunlad ang mga aerob na may libreng access sa oxygen. Nakukuha nila ang kinakailangang enerhiya para sa mga proseso ng buhay sa pamamagitan ng pagsipsip ng oxygen at pag-oxidize ng mga nutritional na materyales.
Ang mga anaerobes ay maaaring umunlad nang walang access sa oxygen. Ang libreng oxygen sa hangin ay may nakakapinsala, mapanirang epekto sa mga microbes na ito. Ang mahigpit (obligadong) anaerobes (tetanus bacillus, ang causative agent ng butyric acid fermentation) ay hindi pinahihintulutan ang oxygen sa lahat. Nakukuha nila ang kinakailangang enerhiya sa pamamagitan ng pagsira sa mga organikong sangkap ng carbohydrates, protina, taba, organikong acid, at alkohol.
Ang mga facultative anaerobes ay gumagamit ng parehong mga sangkap, ngunit may kaugnayan sa mga kondisyon ng kanilang pag-iral maaari nilang baguhin ang anaerobic na uri ng kanilang paghinga sa aerobic. Kaya, ang top-fermenting yeast "na may limitadong daloy ng hangin ay nabubulok ang asukal sa alkohol at carbon dioxide na may sapat na aeration, nakakaranas sila ng aerobic respiration na may kumpletong oksihenasyon ng asukal sa carbon dioxide at tubig sa ilalim ng anaerobic na mga kondisyon na nagpapalit ng glucose sa lactic acid, habang ang ilang enerhiya ay mas mababa kaysa sa aerobes Kapag may kakulangan ng oxygen, ang denitrifying bacteria ay gumagamit ng nitrate oxygen upang i-oxidize ang mga organic compound.
Ang mga halimbawang ibinigay ay nagpapakita ng pagkakaiba-iba ng mga pinagkukunan ng enerhiya at mga paraan ng pagkuha nito sa pamamagitan ng iba't ibang uri ng mikrobyo; Para sa mga kadahilanang ito, ang bakterya ay wala at maaaring magkaroon ng isang solong mekanismo ng paghinga.
Karamihan sa mga aerobic microorganism ay nag-oxidize ng mga organikong sustansya sa panahon ng paghinga sa CO2 at tubig. Dahil ang pinakamataas na antas ng oksihenasyon ng carbon ay nakamit sa molekula ng CO 2, sa kasong ito ay nagsasalita sila ng kumpletong oksihenasyon at nakikilala ang ganitong uri ng paghinga mula sa hindi kumpletong mga oksihenasyon, kung saan ang bahagyang na-oxidized na mga organikong compound ay inilabas bilang mga produktong metabolic.
Sa pamamagitan ng "kumpletong oksihenasyon" ang ibig naming sabihin ay walang pagpapalaya ng anumang mga organikong sangkap; ngunit hindi ito nangangahulugan na ang buong hinihigop na substrate ay na-oxidized. Sa bawat kaso, ang isang makabuluhang bahagi ng substrate (40-70%) ay assimilated, i.e. nagiging mga sangkap ng cell.
Ang mga huling produkto ng "hindi kumpletong oksihenasyon" ay maaaring acetic, gluconic, fumaric, citric, lactic acids at isang bilang ng iba pang mga compound. Dahil ang mga produktong ito ay katulad ng mga nabuo sa panahon ng mga fermentation (propionic, butyric, succinic, lactic acids, atbp.), At dahil din sa katotohanan na ang mga proseso ng pang-industriya na fermentation ay nangangailangan ng mga espesyal na teknikal na aparato (fermenters), ang mga hindi kumpletong oksihenasyon ay tinatawag ding " oxidative fermentation. " o "aerobic fermentation". Ang mga salitang "fermentation" at "fermentation" sa kasong ito ay nagpapakita ng higit sa isang teknolohikal na aspeto.
Uuriin din namin bilang "hindi kumpletong oksihenasyon" ang simpleng abstraction ng hydrogen mula sa substrate at ang paggamit ng mga microorganism upang ma-catalyze ang ilang mga reaksyon na walang kabuluhan para sa kanila sa ilalim ng normal na mga kondisyon. Ang ilang mga halimbawa ng naturang mga oksihenasyon ay tatalakayin sa ibaba.
Paghinga ng bakterya
Ang paghinga ay ang pinaka-advanced na anyo ng proseso ng oxidative at ang pinaka-epektibong paraan upang makakuha ng enerhiya. Ang pangunahing bentahe ng paghinga ay ang enerhiya ng oxidized substance - ang substrate kung saan lumalaki ang microorganism - ay ginagamit nang lubusan. Samakatuwid, sa panahon ng proseso ng paghinga, mas kaunting substrate ang naproseso upang makakuha ng isang tiyak na halaga ng enerhiya kaysa, halimbawa, sa panahon ng pagbuburo.
Larawan 10. Ang papel ng pyruvic acid sa mga proseso ng paghinga at pagbuburo.
Ang proseso ng paghinga ay binubuo sa katotohanan na ang mga carbohydrates (o mga protina, taba at iba pang reserbang sangkap ng cell) ay nabubulok, na-oxidized ng atmospheric oxygen, sa carbon dioxide at tubig. Ang enerhiya na inilabas sa kasong ito ay ginugugol sa pagpapanatili ng mahahalagang tungkulin ng mga organismo, paglaki at pagpaparami. Ang bakterya, dahil sa hindi gaanong sukat ng kanilang mga katawan, ay hindi makakaipon ng malaking halaga ng mga reserbang sangkap. Samakatuwid, pangunahing ginagamit nila ang mga nutrient compound ng kapaligiran.
Sa pangkalahatan, ang paghinga ay maaaring kinakatawan ng sumusunod na equation:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2.87-106 J.
glucose oxygen carbon dioxide tubig enerhiya
Sa likod ng simpleng pormula na ito ay namamalagi ang isang kumplikadong kadena ng mga reaksiyong kemikal, na ang bawat isa ay na-catalyzed ng isang partikular na enzyme.

kanin. 11. Scheme ng glycolytic pathway para sa pagkasira ng carbohydrates.
Ang mga reaksyong enzymatic na nangyayari sa panahon ng paghinga ay pinag-aralan nang mabuti. Ang scheme ng reaksyon ay naging unibersal, ibig sabihin, sa prinsipyo, pareho sa mga hayop, halaman at maraming microorganism, kabilang ang bakterya. Ang proseso ng paghinga sa panahon ng oksihenasyon ng glucose ay binubuo ng mga sumusunod na pangunahing yugto (Larawan 10).
Una, ang phosphorus esters ng glucose ay nabuo - ang monoactivated glucose sa anyo ng diphosphate ay higit na nahati sa dalawang triose phosphates (tatlong carbon compound): phosphoglyceraldehyde at dihydroxyacetone phosphate, na maaaring ma-reversibly convert sa isa't isa.

kanin. 12. Tricarboxylic acid cycle. Ang mga arrow ay nagpapahiwatig ng direksyon, at ang mga numero ay nagpapahiwatig ng pagkakasunud-sunod ng mga reaksyon.
Susunod, ang phosphoglyceraldehyde ay pumapasok sa palitan at na-oxidized sa diphosphoglyceric acid. Ang layunin ng prosesong ito ay ang abstraction ng hydrogen atoms mula sa oxidized substrate at ang paglipat ng hydrogen sa tulong ng mga tiyak na oxidative enzymes sa atmospheric oxygen (tingnan ang Fig. 10, 11).
Ang hydrogen mula sa phosphoglyceraldehyde ay nakakabit sa enzyme nicotine amide dinucleotide (NAD); sa kasong ito, ang aldehyde ay na-oxidized sa isang acid at ang enerhiya ay inilabas. Ang ilan sa enerhiyang ito ay ginugugol sa pagbuo ng ATP; sa kasong ito, ang phosphoric acid ay idinagdag sa adenosine diphosphate y-ADP. Kapag ang ATP ay na-hydrolyzed, ang enerhiya ay inilabas at maaaring gastusin sa iba't ibang mga proseso ng synthesis ng protina at iba pang mga pangangailangan ng cell.
Ang Phosphoglyceric acid ay na-oxidized sa pyruvic acid. Kasabay nito, nabuo din ang ATP, i.e. ang enerhiya ay nakaimbak.
Kinukumpleto nito ang unang - anaerobic - yugto ng proseso ng paghinga, na tinatawag na glycolytic pathway o ang Embden-Meyerhof-Parnas pathway. Hindi kailangan ng oxygen para mangyari ang mga reaksyong ito. Ang nagreresultang pyruvic acid (CH3COCOOH) ay isang kawili-wili at napakahalagang tambalan. Ang mga landas para sa pagkasira ng glucose sa panahon ng paghinga at maraming mga pagbuburo, hanggang sa pagbuo ng pyruvic acid, ay nagpapatuloy sa eksaktong parehong paraan, na unang itinatag ng Russian biochemist na si S.P. Kostychev. Ang Pyruvic acid ay ang sentrong punto kung saan nag-iiba ang mga landas ng paghinga at pagbuburo, kung saan nagsisimula ang isang kadena ng mga pagbabagong enzymatic na tiyak sa isang naibigay na proseso - isang tiyak na kadena ng mga reaksiyong kemikal (Fig. 11).
Sa panahon ng paghinga, ang pyruvic acid ay pumapasok sa tricarboxylic acid cycle (Larawan 12). Ito ay isang kumplikadong saradong bilog ng mga pagbabagong-anyo, bilang isang resulta kung saan ang mga organikong acid na may 4, 5 at 6 na mga carbon atom ay nabuo (malic, lactic, fumaric, a-ketoglutaric at citric) at ang carbon dioxide ay tinanggal.
Una sa lahat, ang CO2 ay nahahati mula sa pyruvic acid na naglalaman ng tatlong carbon atoms - nabuo ang acetic acid, na may coenzyme A ay bumubuo ng aktibong compound - acetyl coenzyme A. Inililipat nito ang natitirang bahagi ng acetic acid (acetyl) sa oxaloacetic acid (4 carbon). atoms), at ang citric acid ay nabuo (6 na carbon atoms). Ang citric acid ay sumasailalim sa ilang mga pagbabago, bilang isang resulta CO2 ay pinakawalan at isang limang-carbon compound ay nabuo - a-ketoglutaric acid. Ang CO2 (ang ikatlong molekula ng carbon dioxide) ay nahahati din mula dito, at ang succinic acid (4 na carbon atoms) ay nabuo, na pagkatapos ay nagiging fumaric, malic at, sa wakas, oxaloacetic acid.
Kinukumpleto nito ang cycle. Ang oxaloacetic acid ay maaaring pumasok muli sa cycle.
Kaya, ang tatlong-carbon pyruvic acid ay pumapasok sa cycle, at sa panahon ng pagbabagong-anyo, 3 CO2 molecule ang pinakawalan.
Ang hydrogen ng pyruvic acid, na inilabas sa panahon ng dehydrogenation sa ilalim ng aerobic na mga kondisyon, ay hindi nananatiling libre - ito ay pumapasok sa respiratory chain (tulad ng hydrogen ng glyceraldehyde, kinuha kapag ito ay na-convert sa glyceric acid). Ito ay isang chain ng oxidative enzymes.
Ang mga enzyme na unang kumukuha ng hydrogen mula sa substrate na na-oxidized ay tinatawag na primary dehydrogenases.
Naglalaman ang mga ito ng di- o tripyridine nucleotides: NAD o NADP at isang partikular na protina. Ang mekanismo ng pagdaragdag ng hydrogen ay pareho:
Na-oxidize na substance - H2 + NAD -> oxidized substance + NAD*H2
Ang hydrogen na ginawa ng dehydrogenase ay idinagdag sa susunod na sistema ng enzyme, flavin enzymes (FMN o FAD).
Mula sa flavin enzymes, ang mga electron ay pumupunta sa cytochromes - mga protina na naglalaman ng bakal (kumplikadong protina). Hindi ang hydrogen atom ang inililipat kasama ang cytochrome chain, ngunit ang mga electron lamang. Sa kasong ito, nagbabago ang valency ng iron: Fe++ - e->Fe++
Ang huling reaksyon ng paghinga ay ang pagdaragdag ng isang proton at isang elektron sa oxygen sa hangin at ang pagbuo ng tubig. Ngunit una, ang molekula ng oxygen ay isinaaktibo sa ilalim ng pagkilos ng enzyme cytochrome oxidase. Ang pag-activate ay bumaba sa katotohanan na ang oxygen ay nakakakuha ng negatibong singil dahil sa pagdaragdag ng isang electron mula sa oxidized substance. Ang hydrogen (proton) ay nakakabit sa activated oxygen, na bumubuo ng tubig.
Bilang karagdagan sa nabanggit na kadena ng mga carrier ng elektron at hydrogen, kilala ang iba. Ang prosesong ito ay mas kumplikado kaysa sa diagram na nakabalangkas.
Ang biological na kahulugan ng mga pagbabagong ito ay ang oksihenasyon ng mga sangkap at ang pagbuo ng enerhiya Bilang resulta ng oksihenasyon ng isang molekula ng asukal (glucose), 12.6-1053 J ng enerhiya ay nakaimbak sa ATP, ang molekula ng asukal mismo ay naglalaman ng 28.6-106. J, samakatuwid, 44% ng enerhiya ay kapaki-pakinabang na ginagamit. Ito ay isang napakataas na kadahilanan ng kahusayan kung ihahambing sa kahusayan ng mga modernong makina.
Ang proseso ng paghinga ay gumagawa ng isang malaking halaga ng enerhiya. Kung ang lahat ng ito ay inilabas nang sabay-sabay, ang cell ay hindi na umiiral. Ngunit hindi ito nangyayari, dahil ang enerhiya ay hindi inilabas nang sabay-sabay, ngunit sa mga yugto, sa maliliit na bahagi. Ang pagpapakawala ng enerhiya sa maliliit na dosis ay dahil sa ang katunayan na ang paghinga ay isang multi-stage na proseso, sa mga indibidwal na yugto kung saan ang iba't ibang mga intermediate na produkto ay nabuo (na may iba't ibang haba ng carbon chain) at ang enerhiya ay inilabas. Ang inilabas na enerhiya ay hindi natupok sa anyo ng init, ngunit naka-imbak sa unibersal na macroergic compound - ATP. Kapag ang ATP ay nasira, ang enerhiya ay maaaring gamitin sa anumang mga proseso na kinakailangan upang mapanatili ang mahahalagang pag-andar ng katawan: para sa synthesis ng iba't ibang mga organikong sangkap, mekanikal na gawain, pagpapanatili ng osmotic pressure ng protoplasm, atbp.
Ang paghinga ay isang proseso na nagbibigay ng enerhiya, ngunit ang biological na kahalagahan nito ay hindi limitado dito. Bilang resulta ng mga reaksiyong kemikal na kasama ng paghinga, nabuo ang isang malaking bilang ng mga intermediate compound. Mula sa mga compound na ito, na may iba't ibang bilang ng mga carbon atom, maraming iba't ibang mga sangkap ng cell ang maaaring synthesize: mga amino acid, fatty acid, taba, protina, bitamina.
Samakatuwid, tinutukoy ng metabolismo ng karbohidrat ang iba pang mga metabolismo (protina, taba). Ito ang malaking kahalagahan nito.
Ang isa sa mga kamangha-manghang katangian ng microbes ay nauugnay sa proseso ng paghinga at ang mga kemikal na reaksyon nito - ang kakayahang maglabas ng nakikitang liwanag - upang luminesce.
Ito ay kilala na ang isang bilang ng mga buhay na organismo, kabilang ang bakterya, ay maaaring maglabas ng nakikitang liwanag. Ang luminescence na dulot ng mga microorganism ay kilala sa loob ng maraming siglo. Ang akumulasyon ng luminescent bacteria sa symbiosis na may maliliit na hayop sa dagat kung minsan ay humahantong sa isang glow sa dagat; Ang luminescence ay nakatagpo din sa panahon ng paglaki ng ilang bakterya sa karne, atbp.
Ang mga pangunahing bahagi, ang pakikipag-ugnayan sa pagitan ng kung saan ay humahantong sa paglabas ng liwanag, ay kinabibilangan ng mga pinababang anyo ng FMN o NAD, molecular oxygen, enzyme luciferase at ang oxidizable compound - luciferin. Ipinapalagay na ang pinababang NAD o FMN ay tumutugon sa luciferase, oxygen at luciferin, bilang isang resulta kung saan ang mga electron sa ilang mga molekula ay napupunta sa isang nasasabik na estado at ang pagbabalik ng mga electron na ito sa antas ng lupa ay sinamahan ng paglabas ng liwanag. Ang luminescence sa microbes ay itinuturing na isang "masasayang proseso", dahil binabawasan nito ang kahusayan ng enerhiya ng paghinga.
Karamihan sa mga organismo sa biosphere ay nasa aerobic na kondisyon. Sa pagkakaroon ng oxygen sa katawan, ang kumpletong "pagsunog" ng mga karbohidrat at iba pang mga molekula ng "cellular fuel" ay nangyayari sa mga huling produkto - C0 2 at H 2 0.
kanin. 19.1. Scheme ng kumpletong oksihenasyon ng glucose sa anim na CO2 molecules at ang energy efficiency ng prosesong ito (ATP balance); Mga landas sa pagbuo ng ATP:
SF - substrate phosphorylation; OP - oxidative phosphorylation
Ang pangkalahatang proseso ng kumpletong oksihenasyon ng glucose sa ilalim ng mga kondisyon ng aerobic ay inilarawan ng stoichiometric equation
Sa kumplikadong ito, multi-stage na proseso ng glucose oxidation, tatlong yugto ang maaaring makilala (Larawan 19.1).
- Sa unang yugto, nagaganap ang mga reaksiyong aerobic glycolysis, kung saan ang glucose ay nahahati sa dalawang molekulang pyruvate. Ang yugtong ito ay bumubuo sa paunang yugto ng pagkabulok ng carbohydrate ito ay tinatawag na "paghahanda".
- Sa ikalawang yugto, ang isang kadena ng mga reaksyon ng oxidative decarboxylation ng pyruvate ay nangyayari, na nagreresulta sa pagbuo ng isa sa mga sentral na metabolite ng cell, acetyl-S-CoA, at ang oksihenasyon ng isang carbon atom ng pyruvate sa CO 2 . Dahil ang dalawang molekula ng pyruvate ay nabuo sa bawat molekula ng glucose, sa yugtong ito ang oksihenasyon ng dalawang carbon atoms ng glucose sa CO 2 ay nangyayari na.
- Ang ikatlong yugto ay isang napakahalagang hanay ng mga reaksyon ng kumpletong oksihenasyon ng acetyl residue, na tinatawag na tricarboxylic acid cycle (TCA cycle).
Ang proseso ng aerobic oxidation ng carbohydrates ay sinamahan ng pagpapalabas ng malaking halaga ng enerhiya (2880 kJ/mol glucose). Kung susumahin natin ang kabuuang ATP yield sa prosesong ito, ito ay magiging 38 molecules (tingnan ang Fig. 19.1). Gaya ng nabanggit kanina (Kabanata 15), ang synthesis ng isang high-energy ATP bond
Kinakailangan ang 31 kJ, at 1178 kJ ang ginagamit para sa synthesis ng 38 ATP molecule, ibig sabihin, higit sa 40% ng libreng enerhiya ng kumpletong oksihenasyon ng glucose ay nakaimbak sa mga molekula ng ATP. Ipinapahiwatig nito ang mataas na kahusayan ng mga proseso ng oxidative na nagaganap sa ilalim ng mga kondisyon ng aerobic kumpara sa mga anaerobic. Sa proseso ng aerobic oxidation, ang metabolically available energy ay naipon sa pinababang NADH at FADH 2 molecules, na pagkatapos ay na-oxidize sa isang prosesong umaasa sa oxygen. oxidative phosphorylation, ang resulta nito ay ang pagbuo ng 34 na molekula ng ATP, at 4 na mga molekula ng ATP lamang ang nabuo sa pamamagitan ng substrate phosphorylation: 2ATP sa glycolysis (yugto I) at 2ATP sa siklo ng TCA (2 pagliko, yugto III).
Dapat pansinin na kung ang unang yugto ng aerobic oxidation ng carbohydrates - glycolysis - ay isang tiyak na proseso ng glucose catabolism, kung gayon ang susunod na dalawa - oxidative decarboxylation ng pyruvate at ang TCA cycle ay nabibilang sa pangkalahatang mga landas ng catabolism (GCP). Matapos ang pagbuo ng pyruvate (C 3 fragment) at acetyl-CoA (C 2 fragment), na nabuo sa panahon ng pagkasira ng hindi lamang glucose, kundi pati na rin ang mga lipid at amino acid, ang mga daanan ng oksihenasyon ng mga sangkap na ito sa mga huling produkto ay nangyayari sa parehong paraan ayon sa mekanismo ng mga reaksyon ng OPC.
Ang aerobic oxidation ng carbohydrates ay ang pangunahing paraan ng paggawa ng enerhiya para sa katawan. Di-tuwiran - dichotomous at direktang - apotomic.
Ang direktang landas ng pagkasira ng glucose ay ikot ng pentose- humahantong sa pagbuo ng mga pentose at ang akumulasyon ng NADPH 2. Ang pentose cycle ay nailalarawan sa pamamagitan ng sunud-sunod na pag-aalis ng bawat isa sa 6 na carbon atoms nito mula sa mga molekula ng glucose na may pagbuo ng 1 molekula ng carbon dioxide at tubig sa isang cycle. Ang pagkasira ng buong molekula ng glucose ay nangyayari sa loob ng 6 na paulit-ulit na cycle.
Ang kahalagahan ng pentose phosphate cycle ng carbohydrate oxidation sa metabolismo ay mahusay:
1. Nagbibigay ito ng pinababang NADP, kinakailangan para sa biosynthesis ng mga fatty acid, kolesterol, atbp. Dahil sa pentose cycle, 50% ng pangangailangan ng katawan para sa NADPH 2 ay sakop.
2. Supply ng pentose phosphates para sa synthesis ng mga nucleic acid at maraming coenzymes.
Ang mga reaksyon ng pentose cycle ay nangyayari sa cytoplasm ng cell.
Sa isang bilang ng mga pathological na kondisyon, ang proporsyon ng pentose pathway ng glucose oxidation ay tumataas.
Hindi direktang landas- pagkasira ng glucose sa carbon dioxide at tubig na may pagbuo ng 36 na molekula ng ATP.
1. Pagkasira ng glucose o glycogen sa pyruvic acid
2. Pagbabago ng pyruvic acid sa acetyl-CoA
Oxidation ng acetyl-CoA sa Krebs cycle sa carbon dioxide at tubig
C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 686 kcal
Sa kaso ng aerobic conversion, ang pyruvic acid ay sumasailalim sa oxidative decarboxylation upang bumuo ng acetyl-CoA, na pagkatapos ay na-oxidize sa carbon dioxide at tubig.
Ang oksihenasyon ng pyruvate sa acetyl-CoA ay na-catalyzed ng pyruvate dehydrogenase system at nangyayari sa ilang mga yugto. Kabuuang reaksyon:
Ang reaksyon ng Pyruvate + NADH + NS-CoA acetyl-CoA + NADH 2 + CO 2 ay halos hindi maibabalik
Ang kumpletong oksihenasyon ng acetyl-CoA ay nangyayari sa tricarboxylic acid cycle o Krebs cycle. Ang prosesong ito ay nagaganap sa mitochondria.
Ang cycle ay binubuo ng 8 magkakasunod na reaksyon:
Sa siklo na ito, ang isang molekula na naglalaman ng 2 carbon atoms (acetic acid sa anyo ng acetyl-CoA) ay tumutugon sa isang molekula ng oxaloacetic acid, na nagreresulta sa pagbuo ng isang compound na may 6 na carbon atoms - citric acid. Sa panahon ng proseso ng dehydrogenation, decarboxylation at preparatory reaction, ang citric acid ay binago pabalik sa oxaloacetic acid, na madaling pinagsama sa isa pang molekula ng acetyl-CoA.
1) acetyl-CoA + oxaloacetate (OA) citric acid
citrate synthase
2) citric acid isocitric acid
aconitate hydratase
3) isocitric acid + NAD-ketoglutaric acid + NADH 2 + CO 2
isocitrate dehydrogenase
4)-ketoglutaric acid + NS-CoA + NADsuccinylSCoA + NADH 2 + CO 2
5) succinyl-CoA + GDP + Succinic acid + GTP + HS-CoA
succinyl CoA synthetase
6) succinic acid + FAD fumaric acid + FADN 2
succinate dehydrogenase
7) fumaric acid + H 2 O L malic acid
fumarate hydratase
8) malate + NADoxaloacetate + NADH 2
malate dehydrogenase
Sa kabuuan, kapag ang isang molekula ng glucose ay nasira sa mga tisyu, 36 na molekula ng ATP ang na-synthesize. Walang alinlangan, ito ay isang mas energetically mas mahusay na proseso kaysa sa glycolysis.
Ang Krebs cycle ay ang karaniwang panghuling landas kung saan ang metabolismo ng mga carbohydrates, fatty acids at amino acids ay nakumpleto. Ang lahat ng mga sangkap na ito ay kasama sa siklo ng Krebs sa isang yugto o iba pa. Susunod, ang biological oxidation o tissue respiration ay nangyayari, ang pangunahing tampok nito ay ang unti-unti itong nagpapatuloy sa maraming yugto ng enzymatic. Ang prosesong ito ay nangyayari sa mitochondria, cellular organelles kung saan ang isang malaking bilang ng mga enzyme ay puro. Ang proseso ay nagsasangkot ng pyridine-dependent dehydrogenases, flavin-dependent dehydrogenases, cytochromes, coenzyme Q - ubiquinone, mga protina na naglalaman ng non-heme iron.
Ang rate ng paghinga ay kinokontrol ng ATP/ADP ratio. Ang mas mababang ratio na ito, ang mas matinding paghinga ay nangyayari, na tinitiyak ang paggawa ng ATP.
Gayundin, ang siklo ng citric acid ay ang pangunahing pinagmumulan ng carbon dioxide sa cell para sa mga reaksyon ng carboxylation, na nagsisimula sa synthesis ng mga fatty acid at gluconeogenesis. Ang parehong carbon dioxide ay nagbibigay ng carbon para sa urea at ilang mga yunit ng purine at pyrimidine rings.
Ang relasyon sa pagitan ng mga proseso ng carbohydrate at nitrogen metabolism ay nakakamit din sa pamamagitan ng mga intermediate na produkto ng citric acid cycle.
Mayroong ilang mga landas kung saan ang mga intermediate ng siklo ng citric acid ay isinama sa proseso ng lipogenesis. Ang pagkasira ng citrate ay humahantong sa pagbuo ng acetyl-CoA, na gumaganap ng papel ng isang precursor sa biosynthesis ng mga fatty acid.
Isocitrate at malate ay nagbibigay ng pagbuo ng NADP, na kung saan ay natupok sa kasunod na reductive yugto ng taba synthesis.
Ang papel na ginagampanan ng pangunahing kadahilanan na tumutukoy sa conversion ng NADH ay nilalaro ng estado ng adenine nucleotides. Ang mataas na ADP at mababang ATP ay nagpapahiwatig ng mababang reserbang enerhiya. Sa kasong ito, ang NADH ay kasangkot sa mga reaksyon ng respiratory chain, na nagpapahusay sa mga proseso ng oxidative phosphorylation na nauugnay sa pag-iimbak ng enerhiya. Ang kabaligtaran na kababalaghan ay sinusunod sa mababang nilalaman ng ADP at mataas na nilalaman ng ATP. Sa pamamagitan ng paglilimita sa sistema ng transportasyon ng elektron, itinataguyod nila ang paggamit ng NADH sa iba pang pagbabawas ng mga reaksyon tulad ng glutamate synthesis at gluconeogenesis.
Ang molecular oxygen sa atmospera ay malinaw na biogenic na pinagmulan. Ang hitsura nito ay nauugnay sa proseso ng photosynthesis ng sinaunang cyanobacteria o kanilang mga ninuno, na unang nagsimulang gumamit ng tubig bilang isang hydrogen donor sa proseso ng photosynthesis. Ang paglipat ng mga prokaryote sa anaerobic oxidation ay naging posible lamang sa isang tiyak na yugto
ebolusyon, kapag ang isang higit pa o hindi gaanong kumpletong respiratory chain ay nabuo sa cell.
Karamihan sa mga aerobic prokaryotic na organismo ay kumokonsumo ng iba't ibang mga organikong compound bilang pinagmumulan ng enerhiya, na nag-o-oxidize sa mga ito sa mga huling produkto na CO 2 at H 2 O. Ang aerobic na oksihenasyon ng mga organikong sangkap sa isang prokaryotic cell ay nangyayari katulad ng aerobic respiration ng mga eukaryotes. Ito ay batay sa oksihenasyon ng pyruvic acid sa pamamagitan ng tricarboxylic acid cycle (TCA - Krebs cycle).
Ang pagsasama ng pyruvic acid sa Krebs cycle ay nauuna sa isang kumplikadong reaksyon ng oksihenasyon nito sa acetyl-K 0 A, na na-catalyzed ng pyruvate dehydrogenase complex:
CH 3 – CO – COOH + K 0 A – SH + NAD + →CH 3 – CO ~ K 0 A + NAD H 2 + CO 2
Ang Krebs cycle mismo ay nagsisimula sa condensation reaction ng acetyl-K 0 A na may molecular oxaloacetic acid na may partisipasyon ng enzyme citras synthase.
Ang Krebs cycle ay gumaganap ng dalawang mahalagang function para sa cell. Sa mga reaksyon ng siklo na ito, ang organikong substrate ay ganap na na-oxidized sa pag-aalis ng hydrogen at ang paglipat nito sa enzyme bilang karagdagan, ang cell ay binibigyan ng mga precursor substance;
Sa buod, ang Krebs cycle ay maaaring ipahayag sa pamamagitan ng sumusunod na equation:
CH 3 COCOOH + 2H 2 O → 3CO 2 + 8H
Ang panimulang substrate para sa tricarboxylic acid cycle ay hindi lamang carbohydrates, kundi pati na rin ang mga fatty acid at maraming amino acids.
Ang Krebs cycle ay nauugnay sa respiratory chain. Ang pangunahing pag-andar ng respiratory chain ay upang mag-imbak ng enerhiya sa cell, na inilabas sa panahon ng proseso ng paglilipat ng elektron sa pamamagitan ng pagbabago nito sa enerhiya ng kemikal ng mga phosphate bond sa mga molekula ng ATP.
Kasama sa respiratory chain ng aerobic prokaryotic organism ang: NAD dehydrogenases, FAD o FMN dehydrogenases, ubiquinone at ang cytochrome system. Ang NAD dehydrogenases ay nag-catalyze sa abstraction ng hydrogen mula sa oxidized substrate at ang paglipat nito sa mga panimulang carrier ng respiratory chain - NAD H 2 dehydrogenase. Mula sa kanila, ang hydrogen ay inililipat sa respiratory chain sa FAD o FMN dehydrogenases, pagkatapos ay sa ubiquinone at pagkatapos ay sa cytochrome system. Kapag ang hydrogen ay inilipat sa pamamagitan ng respiratory chain, ang mga atomo nito ay nahahati sa mga proton at electron. Ang mga proton ay inilabas sa medium, at ang mga electron ay inililipat pa sa kahabaan ng respiratory chain sa terminal carrier - cytochrome oxidase. Ang huli ay naglilipat sa kanila sa panghuling acceptor - molekular na oxygen, na isinaaktibo at pinagsama sa hydrogen.
Ang paglipat ng mga electron sa kahabaan ng respiratory chain sa mas mababang at mas mababang antas ng enerhiya ay humahantong sa pagpapalabas ng isang malaking halaga ng libreng enerhiya, na naipon ng cell sa mga phosphate bond sa
anyo ng mga molekulang ATP. Dahil ang mga reaksyon ng phosphorylation ay isinama sa mga reaksyon ng oksihenasyon, ang prosesong ito ay tinatawag oxidative phosphorylation. Ito ay batay sa pagkakaiba sa redox potensyal ng electron donor at acceptor. Karaniwang nangyayari ang produksyon ng ATP sa mga lugar ng respiratory chain na may malalaking potensyal na pagkakaiba. Gamit ang eukaryotic mitochondria bilang isang halimbawa, tatlong rehiyon sa respiratory chain ang natukoy. Ang unang site ay nauugnay sa paglipat ng hydrogen ng NAD H 2 dehydrogenase sa FAD o FMN dehydrogenase. Ang pangalawang seksyon ay nauugnay sa aktibidad ng ubiquinone, na naglilipat ng mga electron mula sa FAD o FMN dehydrogenase sa cytochrome system.
Ang kahusayan ng oxidative phosphorylation reactions ay hinuhusgahan ng p/o ratio (ang bilang ng mga natupok na molekula ng inorganic phosphorus sa bawat isang atom ng absorbed oxygen Sa mga eokaryote, ang p/o ratio ay 3. Sa maraming prokaryotes, ang p/o). ratio ay maaaring mas mababa sa tatlo, na kung saan ay ipinaliwanag sa pamamagitan ng pagkawala ng ilang mga seksyon respiratory chain.
Sa mga aerobic prokaryote, mayroong mga microorganism na maaaring makakuha ng enerhiya sa pamamagitan ng hindi kumpletong aerobic oxidation ng ilang mga organikong sangkap.
Ang bakterya ng acetic acid ay kinakatawan ng mga maliliit na tungkod; sa isang batang kultura sila ay mobile. Ang lahat ng mga species ay obligadong aerobes, medyo hinihingi sa mga substrate, lalo na ang mga bitamina at pangunahin ang pantothenic acid. Ang pinaka-katangian na kakayahan ng bakterya ng pangkat na ito ay ang pag-oxidize ng ethyl alcohol upang bumuo ng acetic acid na may partisipasyon ng NAD-dependent dehydrogenases.
Binibigyang-diin namin na ang mga proseso ng hindi kumpletong anaerobic oxidation ay walang pagkakatulad sa mga proseso ng fermentation. Ang enerhiya para sa buhay ng cell ay nabuo sa mga reaksyon ng oxidized phosphoration, ngunit sa mas maliit na dami kaysa sa kumpletong aerobic oxidation, dahil ang bahagi nito ay naka-imbak sa mga under-oxidized na mga end product.
Ang iba't ibang uri ng acetic acid bacteria ay may kakayahang gumamit ng parehong monohydric alcohol at polyhydric alcohol - mga sugar derivatives - bilang isang oxidizable substrate. Ang oksihenasyon ng mga monohydric na alkohol ay gumagawa ng iba't ibang mga acid. Ang oksihenasyon ng polyhydric alcohol ay humahantong sa pagbuo ng altose at ketosis. Sa industriya, ang acetic acid bacteria ay ginagamit upang makagawa ng table vinegar at ascorbic acid.
Hindi tulad ng mga eukaryote, na nagsasagawa lamang ng paghinga sa pamamagitan ng oksihenasyon ng mga organikong sangkap, sa mga prokaryote mayroong mga grupo ng mga chemolithotrophic microorganism na may kakayahang mag-oxidize ng mga inorganic na sangkap ng substrate sa pamamagitan ng proseso ng catabolism.
Ang respiratory chain ng chemolithotrophic microorganisms ay karaniwang kinabibilangan ng parehong electron transfer enzymes bilang respiratory chain ng chemoorganotrophs. Ang pagtitiyak ng paggana ng respiratory chain ng grupong ito ng mga microorganism ay na sa panahon ng oksihenasyon ng mga inorganic compound na may iba't ibang mga potensyal na redox, ang mga electron mula sa oxidized substrate ay kasama sa respiratory chain sa iba't ibang antas ng enerhiya. Samakatuwid, upang mabigyan ang cell ng enerhiya, ang mga microorganism ay pinipilit na mag-oxidize ng isang malaking halaga ng substrate.
Ang mga chemolithotrophic microorganism na nakakakuha ng enerhiya sa pamamagitan ng oksihenasyon ng mga inorganic na sangkap ay kinabibilangan ng nitrifying bacteria, iron bacteria, thionic bacteria, atbp.
Mula sa punto ng view ng metabolic lability, ang carboxybacteria ay partikular na interes. Ang mga mikroorganismo na ito ay maaaring kumilos bilang mga autotroph, kumokonsumo ng carbon monoxide (CO) bilang nag-iisang pinagmumulan ng carbon at enerhiya, at bilang mga heterotroph, gamit ang mga organikong sangkap - mga alkohol at mga organikong acid - bilang isang mapagkukunan ng carbon at enerhiya.
Ang kabuuang mga proseso ng catabolism at anabolism ng autotrophic carboxydobacteria ay maaaring katawanin ng sumusunod na equation:
24CO + 11O 2 + H 2 O → 23CO 2 + (CH 2 O),
kung saan (CH 2 O) ang simbolo ng biomass.
Ito ay sumusunod mula sa equation na ang CO oxidation ay isang hindi epektibong paraan upang makakuha ng enerhiya, kaya ang mga microorganism ay napipilitang mag-oxidize ng malaking halaga ng substrate.
Ang pagpapakawala ng CO sa pamamagitan ng modernong transportasyon at mga pang-industriya na negosyo ay nagpaparumi sa kapaligiran ng tambalang ito. Ang tanging paraan upang alisin ang CO mula sa kapaligiran ay ang paggamit nito sa metabolismo ng mga microbial cell.
Anaerobic oxidation: nitrate at sulfate respiration
Ang anaerobic oxidation ay nangyayari lamang sa mga kinatawan ng prokaryotic kingdom. Ito ay likas sa mga microorganism na may kakayahang lumipat mula sa isang aerobic na pamumuhay patungo sa isang anaerobic, gamit ang parehong molecular oxygen at nitrogen mula sa nitrates at sulfur mula sa sulfates bilang ang huling electron acceptor.
Ang isang tipikal na halimbawa ng naturang mga microorganism ay denitrifying bacteria.
Ang respiratory chain ng denitrifying bacteria ay kinabibilangan ng lahat ng pangunahing electron transfer enzyme na katangian ng aerobic respiratory chain. Tanging ang huling link ng sistema ng cytochrome, cytochrome oxidase, ay pinalitan ng nitrate reductase, na nag-catalyze sa paglipat ng mga electron sa nitrate nitrogen. Ang nitrate reductases ay mga inducible na enzyme na na-synthesize ng mga cell lamang sa ilalim ng anaerobic na kondisyon sa pagkakaroon ng mga nitrates sa kapaligiran.
Ang proseso ng denitrification ay binubuo ng 4 na mga yugto ng pagbabawas, ang bawat isa ay na-catalyzed ng isang kaukulang nitrate reductase. Sa unang yugto, ang mga nitrates ay nabawasan sa mga nitrite:
nitrogen +5 na kumukuha ng 2 proton at 2 electron ay nabawasan sa nitrite nitrogen NO 2 - +3:
HINDI 3 - + 2e - + 2H + →NO 2 - + H 2 O.
HINDI 2 - + e - + H + → HINDI + OH -
2NO + 2e - + 2H + → N 2 O + H 2 O
N 2 O + 2e - + 2H + →N 2 + H 2 O
Ang paggamit ng nitrogen bilang isang electron acceptor ay nagpapahintulot sa denitrifying bacteria na ganap na ma-oxidize ang mga organikong sangkap ng substrate sa mga huling produkto CO 2 at H 2 O. Samakatuwid, ang enerhiya na ani ng nitrate respiration ay halos malapit sa maginoo na aerobic oxidation.
Dahil ang denitrifying bacteria ay lumipat sa nitrate respiration lamang kapag nalantad sa anaerobic na kondisyon, ang kanilang adaptasyon sa isang anaerobic na pamumuhay ay dapat ituring na ebolusyonaryong pangalawa at ituring bilang isang pagbabalik sa anaerobiosis mula sa tipikal na aerobic oxidation.
Ang mga bacteria na nagpapababa ng sulfate na kabilang sa genera na Desulfotomaculum, Desulfonema, Desulfovibrio, atbp. ay may kakayahang anaerobic oxidation Ang mga paraan kung saan ang sulfate-reducing bacteria ay nakakakuha ng enerhiya. Ito ang proseso ng pagbuburo ng mga organikong sangkap, na sinamahan ng pagbuo ng ATP bilang isang resulta ng substrate phosphorylation, sulfate respiration, na nagsasangkot ng oksihenasyon ng mga organikong sangkap sa ilalim ng anaerobic na mga kondisyon na may paglipat ng mga electron sa sulfate sulfur. Ang mga bakterya ng heterogenous na grupong ito ay may kakayahang makakuha ng enerhiya sa pamamagitan ng oksihenasyon ng molecular hydrogen kasabay ng pagbabawas ng mga sulfate.
Ang kakayahan ng sulfate-reducing bacteria na gumamit ng molecular hydrogen upang makagawa ng enerhiya ay nagpapahintulot sa kanila na mauri bilang anaerobic chemolithotrophic microorganisms.
Sa proseso ng oksihenasyon ng molecular hydrogen, ang mga bacteria na gumagawa ng methane na gumagamit ng carbon dioxide bilang electron acceptor ay nakakakuha din ng enerhiya. Para sa bakterya ng pangkat na ito, ang CO 2 ay gumaganap bilang parehong pinagmumulan ng carbon at isang electron acceptor:
4H 2 + CO 2 →CH 4 + 2H 2 O
Ang pag-aaral ng iba't ibang uri ng prokaryotic catabolism ay ginagawang posible na ipalagay na ito ay ang pagpapabuti ng mga paraan kung saan ang cell ay nakakakuha ng enerhiya na pinagbabatayan ng ebolusyon ng mga kinatawan ng kaharian na ito.
Ang pinaka sinaunang grupo ng mga prokaryote ay anaerobic bacteria na gumagawa ng enerhiya sa mga proseso ng fermentation dahil sa substrate phosphorylation.
Ang isang makabuluhang yugto sa ebolusyon ng mga prokaryote ay dapat isaalang-alang ang paglitaw ng mga phototrophic bacteria na gumagamit ng sikat ng araw bilang pangunahing mapagkukunan ng enerhiya at CO 2 bilang pangunahing mapagkukunan ng carbon.
Ang pagbuo ng mga photosynthetic aerobes, pangunahin ang cyanobacteria, ay humantong sa pagpapayaman ng kapaligiran na may molekular na oxygen. Sa cell ng aerobic bacteria, isa pang electron transport system ang nabuo at isang nauugnay na mekanismo ng phosphorylation ay oxidative phosphorylation.
Sa kasalukuyan, sa kaharian ng mga prokaryote, nakatagpo tayo ng kamangha-manghang iba't ibang uri ng catabolism. Gayunpaman, ang nangingibabaw at evolutionarily dominanteng uri ng catabolism ay walang alinlangan na aerobic oxidation kasama ang lahat ng pagkakaiba-iba nito ng mga donor at acceptors.