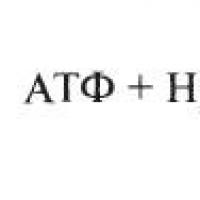Bilang resulta ng hydrolysis ng ATP, nabuo ang AmphADP. ATF: pera ng enerhiya. Oxidative decarboxylation - ang oksihenasyon ng pyruvate sa acetyl-CoA ay nangyayari sa pakikilahok ng isang bilang ng mga enzyme at coenzymes, pinagsama sa istruktura sa isang multienzyme system, makuha
ATP (adenosine triphosphate)– isang organikong tambalan mula sa pangkat ng mga nucleoside triphosphate, na gumaganap ng malaking papel sa ilang proseso ng biochemical, pangunahin sa pagbibigay ng enerhiya sa mga cell.
Pag-navigate sa artikulo
Istraktura at synthesis ng ATP
Ang adenosine triphosphate ay adenine kung saan nakakabit ang tatlong molekula ng orthophosphoric acid. Ang adenine ay bahagi ng maraming iba pang mga compound na laganap sa buhay na kalikasan, kabilang ang mga nucleic acid.
Ang paglabas ng enerhiya, na ginagamit ng katawan para sa iba't ibang layunin, ay nangyayari sa panahon ng hydrolysis ng ATP, na humahantong sa paglitaw ng isa o dalawang libreng molekula ng phosphoric acid. Sa unang kaso, ang adenosine triphosphate ay binago sa adenosine diphosphate (ADP), sa pangalawa, sa adenosine monophosphate (AMP).
Ang synthesis ng ATP, na nangyayari sa isang buhay na organismo dahil sa kumbinasyon ng adenosine diphosphate na may phosphoric acid, ay maaaring mangyari sa maraming paraan:
- Pangunahing: oxidative phosphorylation, na nangyayari sa intracellular organelles - mitochondria, sa panahon ng oksihenasyon ng mga organikong sangkap.
- Ang pangalawang landas: substrate phosphorylation, na nangyayari sa cytoplasm at gumaganap ng isang pangunahing papel sa anaerobic na mga proseso.
Mga function ng ATP
Ang adenosine triphosphate ay hindi gumaganap ng anumang makabuluhang papel sa pag-iimbak ng enerhiya, ngunit sa halip ay gumaganap ng mga function ng transportasyon sa cellular energy metabolism. Ang adenosine triphosphate ay na-synthesize mula sa ADP at sa lalong madaling panahon ay na-convert pabalik sa ADP, na naglalabas ng kapaki-pakinabang na enerhiya.
Kaugnay ng mga vertebrates at mga tao, ang pangunahing tungkulin ng ATP ay upang matiyak ang aktibidad ng motor ng mga fibers ng kalamnan.
Depende sa tagal ng pagsisikap, ito man ay panandaliang trabaho o pangmatagalang (cyclic) load, ang mga proseso ng enerhiya ay medyo naiiba. Ngunit sa lahat ng mga ito, ang adenosine triphosphate ay gumaganap ng isang mahalagang papel.
ATP structural formula:
Bilang karagdagan sa pag-andar ng enerhiya nito, ang adenosine triphosphate ay gumaganap ng isang mahalagang papel sa paghahatid ng signal sa pagitan ng mga selula ng nerbiyos at iba pang mga intercellular na pakikipag-ugnayan, sa regulasyon ng pagkilos ng mga enzyme at hormone. Ito ay isa sa mga panimulang produkto para sa synthesis ng protina.
Gaano karaming mga molekula ng ATP ang ginawa sa panahon ng glycolysis at oksihenasyon?
Ang buhay ng isang molekula ay karaniwang hindi hihigit sa isang minuto, kaya sa anumang sandali ang nilalaman ng sangkap na ito sa katawan ng isang may sapat na gulang ay mga 250 gramo. Sa kabila ng katotohanan na ang kabuuang halaga ng adenosine triphosphate na na-synthesize bawat araw ay karaniwang maihahambing sa sariling timbang ng katawan.
Ang proseso ng glycolysis ay nangyayari sa 3 yugto:
- Paghahanda.
Sa pasukan sa yugtong ito, ang mga molekula ng adenosine triphosphate ay hindi nabuo - Anaerobic.
2 ATP molecule ang nabuo. - Aerobic.
Sa panahon nito, nangyayari ang oksihenasyon ng PVA at pyruvic acid. 36 ATP molecules ay nabuo mula sa 1 glucose molecule.
Sa kabuuan, sa panahon ng glycolysis ng 1 glucose molecule, 38 ATP molecules ang nabuo: 2 sa panahon ng anaerobic stage ng glycolysis, 36 sa panahon ng oksihenasyon ng pyruvic acid.
Ang figure ay nagpapakita ng dalawang pamamaraan Mga imahe ng istraktura ng ATP. Ang adenosine monophosphate (AMP), adenosine diphosphate (ADP), at adenosine triphosphate (ATP) ay nabibilang sa isang klase ng mga compound na tinatawag na nucleotides. Ang nucleotide molecule ay binubuo ng isang limang-carbon na asukal, isang nitrogenous base at phosphoric acid. Sa molekula ng AMP, ang asukal ay kinakatawan ng ribose, at ang base ay adenine. Mayroong dalawang grupo ng pospeyt sa molekula ng ADP, at tatlo sa molekula ng ATP.
Halaga ng ATP
Kapag ang ATP ay nahati sa ADP at ang inorganikong phosphate (Pn) na enerhiya ay inilabas:
Ang reaksyon ay nangyayari sa pagsipsip ng tubig, ibig sabihin, ito ay kumakatawan sa hydrolysis (sa aming artikulo ay nakatagpo namin ang napaka-karaniwang uri ng biochemical na reaksyon ng maraming beses). Ang ikatlong pangkat ng pospeyt na nahati mula sa ATP ay nananatili sa selula sa anyo ng hindi organikong pospeyt (Pn). Ang libreng enerhiya na ani para sa reaksyong ito ay 30.6 kJ bawat 1 mol ng ATP.
Mula sa ADF at pospeyt, maaaring ma-synthesize muli ang ATP, ngunit nangangailangan ito ng paggastos ng 30.6 kJ ng enerhiya sa bawat 1 mole ng bagong nabuong ATP.
Sa ganitong reaksyon, na tinatawag na condensation reaction, ang tubig ay inilabas. Ang pagdaragdag ng pospeyt sa ADP ay tinatawag na reaksyon ng phosphorylation. Ang parehong mga equation sa itaas ay maaaring pagsamahin:

Ang nababaligtad na reaksyon na ito ay na-catalyze ng isang enzyme na tinatawag ATPase.
Ang lahat ng mga cell, tulad ng nabanggit na, ay nangangailangan ng enerhiya upang maisagawa ang kanilang trabaho, at para sa lahat ng mga cell ng anumang organismo ang pinagmumulan ng enerhiya na ito ay nagsisilbing ATP. Samakatuwid, ang ATP ay tinatawag na "universal energy carrier" o "energy currency" ng mga cell. Ang isang naaangkop na pagkakatulad ay mga de-kuryenteng baterya. Tandaan kung bakit hindi natin ginagamit ang mga ito. Sa kanilang tulong maaari kaming makatanggap ng liwanag sa isang kaso, tunog sa isa pa, minsan mekanikal na paggalaw, at kung minsan kailangan namin ng elektrikal na enerhiya mula sa kanila. Ang kaginhawahan ng mga baterya ay maaari nating gamitin ang parehong mapagkukunan ng enerhiya - isang baterya - para sa iba't ibang layunin, depende sa kung saan natin ito inilalagay. Ang ATP ay gumaganap ng parehong papel sa mga cell. Nagbibigay ito ng enerhiya para sa mga magkakaibang proseso tulad ng pag-urong ng kalamnan, paghahatid ng mga nerve impulses, aktibong transportasyon ng mga sangkap o synthesis ng protina, at lahat ng iba pang uri ng aktibidad ng cellular. Upang gawin ito, dapat itong "konektado" lamang sa kaukulang bahagi ng cell apparatus.
Maaaring ipagpatuloy ang pagkakatulad. Dapat munang gawin ang mga baterya, at ang ilan sa mga ito (mga rechargeable), tulad ng , ay maaaring ma-recharge. Kapag ang mga baterya ay ginawa sa isang pabrika, ang isang tiyak na halaga ng enerhiya ay dapat na naka-imbak sa kanila (at sa gayon ay natupok ng pabrika). Ang ATP synthesis ay nangangailangan din ng enerhiya; ang pinagmulan nito ay ang oksihenasyon ng mga organikong sangkap sa panahon ng paghinga. Dahil ang enerhiya ay inilabas sa panahon ng proseso ng oksihenasyon sa phosphorylate ADP, ang naturang phosphorylation ay tinatawag na oxidative phosphorylation. Sa panahon ng photosynthesis, ang ATP ay ginawa mula sa liwanag na enerhiya. Ang prosesong ito ay tinatawag na photophosphorylation (tingnan ang Seksyon 7.6.2). Mayroon ding mga "pabrika" sa cell na gumagawa ng karamihan sa ATP. Ito ay mitochondria; naglalaman ang mga ito ng kemikal na "mga linya ng pagpupulong" kung saan nabuo ang ATP sa panahon ng aerobic respiration. Sa wakas, ang mga pinalabas na "baterya" ay na-recharge din sa cell: pagkatapos ng ATP, na inilabas ang enerhiya na nilalaman nito, ay na-convert sa ADP at Fn, maaari itong mabilis na ma-synthesize muli mula sa ADP at Fn dahil sa enerhiya na natanggap sa proseso. ng paghinga mula sa oksihenasyon ng mga bagong bahagi ng organikong bagay.
halaga ng ATP sa cell sa anumang naibigay na sandali ay napakaliit. Samakatuwid, sa ATF dapat makita lamang ng isa ang carrier ng enerhiya, at hindi ang depot nito. Ang mga sangkap tulad ng taba o glycogen ay ginagamit para sa pangmatagalang pag-iimbak ng enerhiya. Ang mga cell ay napaka-sensitibo sa mga antas ng ATP. Habang tumataas ang rate ng paggamit nito, tumataas din ang rate ng proseso ng paghinga na nagpapanatili sa antas na ito.
Papel ng ATP bilang isang link sa pagitan ng cellular respiration at mga proseso na kinasasangkutan ng pagkonsumo ng enerhiya, ay makikita mula sa figure na ito ay mukhang simple, ngunit ito ay naglalarawan ng isang napakahalagang pattern.
Kaya naman masasabi na, sa pangkalahatan, ang tungkulin ng paghinga ay upang gumawa ng ATP.

Isa-isahin natin ang sinabi sa itaas.
1. Ang synthesis ng ATP mula sa ADP at inorganic phosphate ay nangangailangan ng 30.6 kJ ng enerhiya bawat 1 mole ng ATP.
2. Ang ATP ay naroroon sa lahat ng mga buhay na selula at samakatuwid ay isang unibersal na tagapagdala ng enerhiya. Walang ibang mga carrier ng enerhiya ang ginagamit. Pinapasimple nito ang bagay - ang kinakailangang cellular apparatus ay maaaring maging mas simple at gumana nang mas mahusay at matipid.
3. Ang ATP ay madaling naghahatid ng enerhiya sa anumang bahagi ng cell sa anumang proseso na nangangailangan ng enerhiya.
4. Mabilis na naglalabas ng enerhiya ang ATP. Ito ay nangangailangan lamang ng isang reaksyon - hydrolysis.
5. Ang rate ng produksyon ng ATP mula sa ADP at inorganic phosphate (respiration process rate) ay madaling nababagay ayon sa mga pangangailangan.
6. Ang ATP ay synthesize sa panahon ng paghinga dahil sa kemikal na enerhiya na inilabas sa panahon ng oksihenasyon ng mga organikong sangkap tulad ng glucose, at sa panahon ng photosynthesis dahil sa solar energy. Ang pagbuo ng ATP mula sa ADP at inorganic phosphate ay tinatawag na phosphorylation reaction. Kung ang enerhiya para sa phosphorylation ay ibinibigay ng oksihenasyon, pagkatapos ay nagsasalita tayo ng oxidative phosphorylation (ang prosesong ito ay nangyayari sa panahon ng paghinga), ngunit kung ang liwanag na enerhiya ay ginagamit para sa phosphorylation, kung gayon ang proseso ay tinatawag na photophosphorylation (ito ay nangyayari sa panahon ng photosynthesis).
Ipadala ang iyong mabuting gawa sa base ng kaalaman ay simple. Gamitin ang form sa ibaba
Ang mga mag-aaral, nagtapos na mga mag-aaral, mga batang siyentipiko na gumagamit ng base ng kaalaman sa kanilang pag-aaral at trabaho ay lubos na magpapasalamat sa iyo.
Nai-post sa http://www.allbest.ru/
- Panimula
- 1.1 Mga kemikal na katangian ng ATP
- 1.2 Mga pisikal na katangian ng ATP
- 2.1
- 3.1 Papel sa cell
- 3.2 Papel sa paggana ng enzyme
- 3.4 Iba pang mga function ng ATP
- Konklusyon
- Bibliograpiya
Listahan ng mga simbolo
ATP - adenosine triphosphate
ADP - adenosine diphosphate
AMP - adenosine monophosphate
RNA - ribonucleic acid
DNA - deoxyribonucleic acid
NAD - nicotinamide adenine dinucleotide
PVC - pyruvic acid
G-6-P - phosphoglucose isomerase
F-6-F - fructose-6-phosphate
TPP - thiamine pyrophosphate
FAD - phenyladenine dinucleotide
Fn - walang limitasyong pospeyt
G - entropy
RNR - ribonucleotide reductase
Panimula
Ang pangunahing pinagmumulan ng enerhiya para sa lahat ng nabubuhay na nilalang na naninirahan sa ating planeta ay ang enerhiya ng sikat ng araw, na direktang ginagamit lamang ng mga selula ng berdeng halaman, algae, berde at lila na bakterya. Sa mga selulang ito, ang mga organikong sangkap (carbohydrates, taba, protina, nucleic acid, atbp.) ay nabuo mula sa carbon dioxide at tubig sa panahon ng proseso ng photosynthesis. Sa pamamagitan ng pagkain ng mga halaman, ang mga hayop ay nakakakuha ng mga organikong sangkap sa tapos na anyo. Ang enerhiya na nakaimbak sa mga sangkap na ito ay pumasa sa kanila sa mga selula ng mga heterotrophic na organismo.
Sa mga selula ng mga organismo ng hayop, ang enerhiya ng mga organikong compound sa panahon ng kanilang oksihenasyon ay na-convert sa enerhiya ng ATP. (Ang carbon dioxide at tubig na inilabas sa kasong ito ay muling ginagamit ng mga autotrophic na organismo para sa mga proseso ng photosynthesis.) Ang lahat ng mahahalagang proseso ay isinasagawa gamit ang enerhiya ng ATP: biosynthesis ng mga organikong compound, paggalaw, paglaki, paghahati ng cell, atbp.
Ang paksa ng pagbuo at paggamit ng ATP sa katawan ay hindi bago sa loob ng mahabang panahon, ngunit bihira na makakahanap ka ng isang kumpletong talakayan ng pareho sa isang mapagkukunan at kahit na mas madalas ang isang pagsusuri ng parehong mga prosesong ito nang sabay-sabay at sa iba't ibang organismo.
Kaugnay nito, ang kaugnayan ng aming trabaho ay naging isang masusing pag-aaral ng pagbuo at paggamit ng ATP sa mga buhay na organismo, dahil ang paksang ito ay hindi pinag-aralan sa wastong antas sa popular na panitikan sa agham.
Ang layunin ng aming gawain ay:
· pag-aaral ng mga mekanismo ng pagbuo at mga paraan ng paggamit ng ATP sa katawan ng mga hayop at tao.
Binigyan kami ng mga sumusunod na gawain:
· Pag-aralan ang kemikal na katangian at katangian ng ATP;
· Pag-aralan ang mga landas ng pagbuo ng ATP sa mga buhay na organismo;
· Isaalang-alang ang mga paraan ng paggamit ng ATP sa mga buhay na organismo;
· Isaalang-alang ang kahalagahan ng ATP para sa katawan ng tao at mga hayop.
Kabanata 1. Kemikal na katangian at katangian ng ATP
1.1 Mga kemikal na katangian ng ATP
Ang adenosine triphosphate ay isang nucleotide na gumaganap ng isang napakahalagang papel sa metabolismo ng enerhiya at mga sangkap sa mga organismo; Una sa lahat, ang tambalan ay kilala bilang isang unibersal na pinagmumulan ng enerhiya para sa lahat ng mga prosesong biochemical na nagaganap sa mga buhay na sistema. Ang ATP ay natuklasan noong 1929 ni Karl Lohmann, at noong 1941 ay ipinakita ni Fritz Lipmann na ang ATP ang pangunahing tagapagdala ng enerhiya sa cell.
Systematic na pangalan ng ATP:
9-in-D-ribofuranosyladenine-5"-triphosphate, o
9-in-D-ribofuranosyl-6-amino-purine-5"-triphosphate.
Sa kemikal, ang ATP ay ang triphosphate ester ng adenosine, na isang derivative ng adenine at ribose.
Ang purine nitrogenous base - adenine - ay konektado sa pamamagitan ng isang N-glycosidic bond sa 1" carbon ng ribose. Tatlong molekula ng phosphoric acid ay sunud-sunod na nakakabit sa 5" carbon ng ribose, na itinalaga ayon sa pagkakabanggit ng mga titik: b, c at d .
Ang istraktura ng ATP ay katulad ng adenine nucleotide na bahagi ng RNA, tanging sa halip na isang phosphoric acid, ang ATP ay naglalaman ng tatlong phosphoric acid residues. Ang mga cell ay hindi maaaring maglaman ng mga acid sa kapansin-pansing dami, ngunit ang kanilang mga asin lamang. Samakatuwid, ang phosphoric acid ay pumapasok sa ATP bilang isang nalalabi (sa halip na ang pangkat ng OH ng acid ay mayroong negatibong sisingilin na oxygen atom).
Sa ilalim ng pagkilos ng mga enzyme, ang molekula ng ATP ay madaling sumasailalim sa hydrolysis, iyon ay, nakakabit ito ng isang molekula ng tubig at nasira upang bumuo ng adenosine diphosphoric acid (ADP):
ATP + H2O ADP + H3PO4.
Ang pag-aalis ng isa pang phosphoric acid residue ay nagpapalit ng ADP sa adenosine monophosphoric acid AMP:
ADP + H2O AMP + H3PO4.
Ang mga reaksyong ito ay nababaligtad, ibig sabihin, ang AMP ay maaaring maging ADP at pagkatapos ay maging ATP, na nag-iipon ng enerhiya. Ang pagsira sa isang regular na peptide bond ay naglalabas lamang ng 12 kJ/mol ng enerhiya. At ang mga bono na nakakabit sa mga residue ng phosphoric acid ay mataas na enerhiya (tinatawag din silang mataas na enerhiya): ang pagkasira ng bawat isa sa kanila ay naglalabas ng 40 kJ/mol ng enerhiya. Samakatuwid, ang ATP ay gumaganap ng isang pangunahing papel sa mga cell bilang isang unibersal na biological energy accumulator. Ang mga molekula ng ATP ay na-synthesize sa mitochondria at chloroplasts (maliit lamang na halaga ang na-synthesize sa cytoplasm), at pagkatapos ay pumapasok sa iba't ibang organelles ng cell, na nagbibigay ng enerhiya para sa lahat ng mahahalagang proseso.
Dahil sa enerhiya ng ATP, nangyayari ang paghahati ng cell, ang aktibong transportasyon ng mga sangkap sa mga lamad ng cell, ang pagpapanatili ng potensyal na elektrikal ng lamad sa panahon ng paghahatid ng mga nerve impulses, pati na rin ang biosynthesis ng mga high-molecular compound at pisikal na gawain.
Sa tumaas na pagkarga (halimbawa, sa maikling pagtakbo), ang mga kalamnan ay gumagana nang eksklusibo dahil sa supply ng ATP. Sa mga selula ng kalamnan, ang reserbang ito ay sapat na para sa ilang dosenang mga contraction, at pagkatapos ay dapat na mapunan ang halaga ng ATP. Ang synthesis ng ATP mula sa ADP at AMP ay nangyayari dahil sa enerhiya na inilabas sa panahon ng pagkasira ng carbohydrates, lipids at iba pang mga sangkap. Ang pagsasagawa ng mental na gawain ay nangangailangan din ng malaking halaga ng ATP. Para sa kadahilanang ito, ang mga taong may mental na trabaho ay nangangailangan ng mas mataas na halaga ng glucose, ang pagkasira nito ay nagsisiguro sa synthesis ng ATP.
1.2 Mga pisikal na katangian ng ATP
Ang ATP ay binubuo ng adenosine at ribose - at tatlong grupo ng pospeyt. Ang ATP ay lubos na natutunaw sa tubig at medyo matatag sa mga solusyon sa pH 6.8-7.4, ngunit mabilis na na-hydrolyzed sa matinding pH. Samakatuwid, ang ATP ay pinakamahusay na nakaimbak sa mga anhydrous salts.
Ang ATP ay isang hindi matatag na molekula. Sa unbuffered na tubig, ito ay hydrolyzes sa ADP at pospeyt. Ito ay dahil ang lakas ng mga bono sa pagitan ng mga grupo ng pospeyt sa ATP ay mas mababa kaysa sa lakas ng mga bono ng hydrogen (hydration bond) sa pagitan ng mga produkto nito (ADP + phosphate) at tubig. Kaya, kung ang ATP at ADP ay nasa chemical equilibrium sa tubig, halos lahat ng ATP ay tuluyang mako-convert sa ADP. Ang isang sistema na malayo sa ekwilibriyo ay naglalaman ng libreng enerhiya ng Gibbs, at may kakayahang gumawa ng trabaho. Ang mga nabubuhay na selula ay nagpapanatili ng ratio ng ATP sa ADP sa isang puntong sampung order ng magnitude mula sa equilibrium, na may konsentrasyon ng ATP na isang libong beses na mas mataas kaysa sa konsentrasyon ng ADP. Ang paglipat na ito mula sa posisyon ng balanse ay nangangahulugan na ang hydrolysis ng ATP sa cell ay naglalabas ng malaking halaga ng libreng enerhiya.
Ang dalawang high-energy phosphate bond (yaong nag-uugnay sa mga katabing phosphate) sa isang molekula ng ATP ay responsable para sa mataas na nilalaman ng enerhiya ng molekula na iyon. Ang enerhiya na nakaimbak sa ATP ay maaaring ilabas sa pamamagitan ng hydrolysis. Matatagpuan sa distal sa ribose sugar, ang pangkat ng g-phosphate ay may mas mataas na enerhiya ng hydrolysis kaysa sa b- o b-phosphate. Ang mga bono na nabuo pagkatapos ng hydrolysis o phosphorylation ng isang ATP residue ay mas mababa sa enerhiya kaysa sa iba pang mga ATP bond. Sa panahon ng enzyme-catalyzed ATP hydrolysis o ATP phosphorylation, ang magagamit na libreng enerhiya ay maaaring gamitin ng mga nabubuhay na sistema upang gumawa ng trabaho.
Anumang hindi matatag na sistema ng mga potensyal na reaktibong molekula ay maaaring magsilbing isang paraan upang mag-imbak ng libreng enerhiya kung ang mga selula ay napanatili ang kanilang konsentrasyon na malayo sa punto ng ekwilibriyo ng reaksyon. Gayunpaman, tulad ng karamihan sa mga polymeric biomolecules, ang pagkasira ng RNA, DNA at ATP sa mga simpleng monomer ay nagsasangkot ng parehong paglabas ng enerhiya at entropy, pagtaas ng mga pagsasaalang-alang sa parehong karaniwang konsentrasyon at mga konsentrasyon na matatagpuan sa cell.
Ang karaniwang dami ng enerhiya na inilabas bilang resulta ng ATP hydrolysis ay maaaring kalkulahin mula sa mga pagbabago sa enerhiya na hindi nauugnay sa natural (standard) na mga kondisyon, pagkatapos ay iwasto ang biological na konsentrasyon. Ang netong pagbabago sa thermal energy (enthalpy) sa karaniwang temperatura at presyon para sa decomposition ng ATP sa ADP at inorganic phosphates ay 20.5 kJ/mol, na may libreng pagbabago sa enerhiya na 3.4 kJ/mol. Ang enerhiya na inilabas ng pagkasira ng pospeyt o pyrophosphate mula sa ATP hanggang sa pamantayan ng estado na 1 M ay:
ATP + H 2 O > ADP + P i DG ? = - 30.5 kJ/mol (-7.3 kcal/mol)
ATP + H 2 O > AMP + PP at DG ? = - 45.6 kJ/mol (-10.9 kcal/mol)
Ang mga halagang ito ay maaaring gamitin upang kalkulahin ang mga pagbabago sa enerhiya sa ilalim ng mga kondisyong pisyolohikal at cellular ATP/ADP. Gayunpaman, mas madalas na gumagana ang isang mas kinakatawan na kahalagahan na tinatawag na singil ng enerhiya. Ang mga halaga ay ibinibigay para sa libreng enerhiya ng Gibbs. Ang mga reaksyong ito ay nakasalalay sa isang bilang ng mga kadahilanan, kabilang ang kabuuang lakas ng ionic at ang pagkakaroon ng mga alkaline earth metal tulad ng Mg 2+ at Ca 2+ ions. Sa normal na kondisyon, ang DG ay humigit-kumulang -57 kJ/mol (-14 kcal/mol).
protina biological na enerhiya ng baterya
Kabanata 2. Mga landas para sa pagbuo ng ATP
Sa katawan, ang ATP ay synthesize sa pamamagitan ng phosphorylation ng ADP:
ADP + H 3 PO 4 + enerhiya> ATP + H 2 O.
Ang Phosphorylation ng ADP ay posible sa dalawang paraan: substrate phosphorylation at oxidative phosphorylation (gamit ang enerhiya ng oxidizing substances). Ang bulk ng ATP ay nabuo sa mitochondrial membranes sa panahon ng oxidative phosphorylation ng H-dependent ATP synthase. Ang substrate phosphorylation ng ATP ay hindi nangangailangan ng pakikilahok ng mga enzyme ng lamad na nangyayari sa panahon ng glycolysis o sa pamamagitan ng paglipat ng isang grupo ng pospeyt mula sa iba pang mga compound na may mataas na enerhiya.
Ang mga reaksyon ng ADP phosphorylation at ang kasunod na paggamit ng ATP bilang isang mapagkukunan ng enerhiya ay bumubuo ng isang paikot na proseso na siyang kakanyahan ng metabolismo ng enerhiya.
Sa katawan, ang ATP ay isa sa mga pinaka-madalas na na-renew na mga sangkap. Kaya sa mga tao, ang lifespan ng isang molekula ng ATP ay mas mababa sa 1 minuto. Sa araw, ang isang molekula ng ATP ay dumadaan sa isang average ng 2000-3000 na mga siklo ng resynthesis (ang katawan ng tao ay nag-synthesize ng mga 40 kg ng ATP bawat araw), iyon ay, halos walang reserbang ATP na nilikha sa katawan, at para sa normal na buhay ito ay kinakailangan upang patuloy na mag-synthesize ng mga bagong molekula ng ATP.
Oxidative phosphorylation -
Gayunpaman, ang mga karbohidrat ay kadalasang ginagamit bilang isang substrate. Kaya, ang mga selula ng utak ay hindi maaaring gumamit ng anumang iba pang substrate para sa nutrisyon maliban sa carbohydrates.
Ang mga pre-complex na carbohydrates ay nahahati sa mga simple, na humahantong sa pagbuo ng glucose. Ang glucose ay isang unibersal na substrate sa proseso ng cellular respiration. Ang glucose oxidation ay nahahati sa 3 yugto:
1. glycolysis;
2. oxidative decarboxylation at ang Krebs cycle;
3. oxidative phosphorylation.
Sa kasong ito, ang glycolysis ay isang pangkaraniwang yugto para sa aerobic at anaerobic respiration.
2 .1.1 GlicoLiz- isang enzymatic na proseso ng sunud-sunod na pagkasira ng glucose sa mga cell, na sinamahan ng synthesis ng ATP. Ang glycolysis sa ilalim ng mga kondisyon ng aerobic ay humahantong sa pagbuo ng pyruvic acid (pyruvate), ang glycolysis sa ilalim ng mga kondisyon ng anaerobic ay humahantong sa pagbuo ng lactic acid (lactate). Ang Glycolysis ay ang pangunahing landas ng glucose catabolism sa mga hayop.
Ang glycolytic pathway ay binubuo ng 10 sequential reactions, bawat isa ay na-catalyzed ng isang hiwalay na enzyme.
Ang proseso ng glycolysis ay maaaring nahahati sa dalawang yugto. Ang unang yugto, na nangyayari sa pagkonsumo ng enerhiya ng 2 molekula ng ATP, ay binubuo ng paghahati ng isang molekula ng glucose sa 2 mga molekula ng glyceraldehyde-3-phosphate. Sa ikalawang yugto, ang NAD-dependent na oksihenasyon ng glyceraldehyde-3-phosphate ay nangyayari, na sinamahan ng synthesis ng ATP. Ang glycolysis mismo ay isang ganap na anaerobic na proseso, iyon ay, hindi ito nangangailangan ng pagkakaroon ng oxygen para mangyari ang mga reaksyon.
Ang Glycolysis ay isa sa mga pinakalumang proseso ng metabolic, na kilala sa halos lahat ng nabubuhay na organismo. Marahil, ang glycolysis ay lumitaw higit sa 3.5 bilyong taon na ang nakalilipas sa primordial prokaryotes.
Ang resulta ng glycolysis ay ang conversion ng isang molekula ng glucose sa dalawang molekula ng pyruvic acid (PVA) at ang pagbuo ng dalawang pagbabawas na katumbas sa anyo ng coenzyme NADH.
Ang kumpletong equation para sa glycolysis ay:
C 6 H 12 O 6 + 2NAD + + 2ADP + 2P n = 2NAD H + 2PVK + 2ATP + 2H 2 O + 2H +.
Sa kawalan o kakulangan ng oxygen sa cell, ang pyruvic acid ay sumasailalim sa pagbawas sa lactic acid, kung gayon ang pangkalahatang equation ng glycolysis ay ang mga sumusunod:
C 6 H 12 O 6 + 2ADP + 2P n = 2 lactate + 2ATP + 2H 2 O.
Kaya, sa panahon ng anaerobic breakdown ng isang glucose molecule, ang kabuuang net yield ng ATP ay dalawang molekula na nakuha sa mga reaksyon ng substrate phosphorylation ng ADP.
Sa mga aerobic na organismo, ang mga huling produkto ng glycolysis ay sumasailalim sa karagdagang pagbabago sa mga biochemical cycle na may kaugnayan sa cellular respiration. Bilang isang resulta, pagkatapos ng kumpletong oksihenasyon ng lahat ng mga metabolite ng isang molekula ng glucose sa huling yugto ng paghinga ng cellular - oxidative phosphorylation, na nangyayari sa mitochondrial respiratory chain sa pagkakaroon ng oxygen - isang karagdagang 34 o 36 na molekula ng ATP ay na-synthesize para sa bawat glucose molekula.
Ang unang reaksyon ng glycolysis ay ang phosphorylation ng isang molekula ng glucose, na nangyayari sa pakikilahok ng tissue-specific enzyme hexokinase na may paggasta ng enerhiya ng 1 molekula ng ATP; Ang aktibong anyo ng glucose ay nabuo - glucose-6-phosphate (G-6-F):
Upang maganap ang reaksyon, ang pagkakaroon ng mga Mg 2+ ions sa daluyan ay kinakailangan, kung saan ang molekula ng ATP ay kumplikadong nakagapos. Ang reaksyong ito ay hindi maibabalik at ito ang una susi reaksyon glycolysis.
Ang phosphorylation ng glucose ay may dalawang layunin: una, dahil sa ang katunayan na ang plasma membrane, na natatagusan sa neutral na molekula ng glucose, ay hindi pinapayagan ang negatibong sisingilin na mga molekula ng G-6-P, ang phosphorylated glucose ay naka-lock sa loob ng cell. Pangalawa, sa panahon ng phosphorylation, ang glucose ay na-convert sa isang aktibong anyo na maaaring lumahok sa mga biochemical reaksyon at maisama sa mga metabolic cycle.
Ang hepatic isoenzyme ng hexokinase, glucokinase, ay mahalaga sa pag-regulate ng mga antas ng glucose sa dugo.
Sa susunod na reaksyon ( 2 ) sa pamamagitan ng enzyme phosphoglucoisomerase G-6-P ay na-convert sa fructose 6-phosphate (F-6-F):
Walang kinakailangang enerhiya para sa reaksyong ito at ang reaksyon ay ganap na mababaligtad. Sa yugtong ito, ang fructose ay maaari ding isama sa proseso ng glycolysis sa pamamagitan ng phosphorylation.
Pagkatapos, ang dalawang reaksyon ay sumusunod halos kaagad nang sunud-sunod: hindi maibabalik na phosphorylation ng fructose-6-phosphate ( 3 ) at reversible aldol cleavage ng resulta fructose 1,6-biphosphate (F-1.6-bF) sa dalawang triose ( 4 ).
Ang Phosphorylation ng P-6-P ay isinasagawa ng phosphofructokinase na may paggasta ng enerhiya ng isa pang molekula ng ATP; ito ang pangalawa susi reaksyon glycolysis, tinutukoy ng regulasyon nito ang intensity ng glycolysis sa kabuuan.
cleavage ni Aldol F-1.6-bF nangyayari sa ilalim ng pagkilos ng fructose-1,6-bisphosphate aldolase:
Bilang resulta ng ikaapat na reaksyon, dihydroxyacetone phosphate At glyceraldehyde-3-phosphate, at ang una ay halos agad na nasa ilalim ng impluwensya phosphotriose isomerase papunta sa pangalawa ( 5 ), na nakikilahok sa mga karagdagang pagbabago:
Ang bawat molekula ng glyceraldehyde phosphate ay na-oxidize ng NAD+ sa pagkakaroon ng dehydrogenases glyceraldehyde phosphate dati 1,3- dat phosphoglyce- rata (6 ):
Kasunod ng 1,3-diphosphoglycerate na naglalaman ng isang mataas na enerhiya na bono sa posisyon 1, ang enzyme phosphoglycerate kinase ay naglilipat ng phosphoric acid residue sa ADP molecule (reaksyon 7 ) - isang molekula ng ATP ay nabuo:
Ito ang unang reaksyon ng substrate phosphorylation. Mula sa sandaling ito, ang proseso ng pagkasira ng glucose ay tumigil na hindi kumikita sa mga tuntunin ng enerhiya, dahil ang mga gastos sa enerhiya sa unang yugto ay nabayaran: 2 ATP molecule ay synthesized (isa para sa bawat 1,3-diphosphoglycerate) sa halip na ang dalawang ginugol sa ang mga reaksyon 1 At 3 . Para mangyari ang reaksyong ito, kinakailangan ang pagkakaroon ng ADP sa cytosol, iyon ay, kapag may labis na ATP sa cell (at kakulangan ng ADP), bumababa ang bilis nito. Dahil ang ATP, na hindi na-metabolize, ay hindi idineposito sa cell ngunit nawasak lamang, ang reaksyong ito ay isang mahalagang regulator ng glycolysis.
Pagkatapos ay sunud-sunod: phosphoglycerol mutase forms 2-phospho- glycerate (8 ):
Mga anyo ng enolase phosphoenolpyruvate (9 ):
Sa wakas, ang pangalawang reaksyon ng substrate phosphorylation ng ADP ay nangyayari sa pagbuo ng enol form ng pyruvate at ATP ( 10 ):
Ang reaksyon ay nangyayari sa ilalim ng pagkilos ng pyruvate kinase. Ito ang huling pangunahing reaksyon ng glycolysis. Ang isomerization ng enol form ng pyruvate hanggang pyruvate ay nangyayari nang hindi enzymatically.
Mula nang mabuo ito F-1.6-bF Ang mga reaksyon lamang na naglalabas ng enerhiya ay nangyayari 7 At 10 , kung saan nangyayari ang substrate phosphorylation ng ADP.
Regulasyon glycolysis
Mayroong lokal at pangkalahatang regulasyon.
Ang lokal na regulasyon ay isinasagawa sa pamamagitan ng pagbabago ng aktibidad ng mga enzyme sa ilalim ng impluwensya ng iba't ibang mga metabolite sa loob ng cell.
Ang regulasyon ng glycolysis sa kabuuan, kaagad para sa buong organismo, ay nangyayari sa ilalim ng impluwensya ng mga hormone, na, na nakakaimpluwensya sa pamamagitan ng mga molekula ng pangalawang messenger, nagbabago ng intracellular metabolism.
Ang insulin ay may mahalagang papel sa pagpapasigla ng glycolysis. Ang glucagon at adrenaline ay ang pinaka makabuluhang hormonal inhibitors ng glycolysis.
Pinasisigla ng insulin ang glycolysis sa pamamagitan ng:
· pag-activate ng reaksyon ng hexokinase;
· pagpapasigla ng phosphofructokinase;
· pagpapasigla ng pyruvate kinase.
Ang iba pang mga hormone ay nakakaimpluwensya rin sa glycolysis. Halimbawa, pinipigilan ng somatotropin ang glycolytic enzymes, at ang mga thyroid hormone ay mga stimulant.
Ang Glycolysis ay kinokontrol sa pamamagitan ng ilang mahahalagang hakbang. Mga reaksyon na na-catalyze ng hexokinase ( 1 ), phosphofructokinase ( 3 ) at pyruvate kinase ( 10 ) ay nailalarawan sa pamamagitan ng isang makabuluhang pagbaba sa libreng enerhiya at halos hindi maibabalik, na nagpapahintulot sa kanila na maging epektibong mga punto ng regulasyon ng glycolysis.
Ang Glycolysis ay isang catabolic pathway na may pambihirang kahalagahan. Nagbibigay ito ng enerhiya para sa mga cellular reaction, kabilang ang synthesis ng protina. Ang mga intermediate na produkto ng glycolysis ay ginagamit sa synthesis ng mga taba. Ang pyruvate ay maaari ding gamitin upang synthesize ang alanine, aspartate at iba pang mga compound. Salamat sa glycolysis, ang pagganap ng mitochondrial at pagkakaroon ng oxygen ay hindi nililimitahan ang lakas ng kalamnan sa panahon ng panandaliang matinding pagkarga.
2.1.2 Oxidative decarboxylation - ang oksihenasyon ng pyruvate sa acetyl-CoA ay nangyayari sa partisipasyon ng isang bilang ng mga enzyme at coenzymes, na istrukturang pinagsama sa isang multienzyme system na tinatawag na pyruvate dehydrogenase complex.
Sa yugto I ng prosesong ito, ang pyruvate ay nawawala ang carboxyl group nito bilang resulta ng pakikipag-ugnayan sa thiamine pyrophosphate (TPP) sa aktibong sentro ng enzyme pyruvate dehydrogenase (E 1). Sa stage II, ang oxyethyl group ng E 1 -TPP-CHOH-CH 3 complex ay na-oxidize upang bumuo ng isang acetyl group, na sabay-sabay na inililipat sa lipoic acid amide (coenzyme) na nauugnay sa enzyme dihydrolipoylacetyltransferase (E 2). Ang enzyme na ito ay nag-catalyze sa stage III - ang paglipat ng acetyl group sa coenzyme CoA (HS-KoA) na may pagbuo ng panghuling produkto na acetyl-CoA, na isang high-energy (macroergic) compound.
Sa stage IV, ang oxidized form ng lipoamide ay muling nabuo mula sa pinababang dihydrolipoamide-E 2 complex. Sa pakikilahok ng enzyme dihydrolipoyl dehydrogenase (E 3), ang hydrogen ay inililipat mula sa pinababang sulfhydryl na mga grupo ng dihydrolipoamide sa FAD, na kumikilos bilang isang prosthetic na grupo ng enzyme na ito at mahigpit na nakagapos dito. Sa yugto V, ang pinababang FADH 2 dihydro-lipoyl dehydrogenase ay naglilipat ng hydrogen sa coenzyme NAD upang mabuo ang NADH + H +.
Ang proseso ng oxidative decarboxylation ng pyruvate ay nangyayari sa mitochondrial matrix. Kabilang dito (bilang bahagi ng isang kumplikadong multienzyme complex) 3 enzymes (pyruvate dehydrogenase, dihydrolipoyl acetyltransferase, dihydrolipoyl dehydrogenase) at 5 coenzymes (TPF, lipoic acid amide, coenzyme A, FAD at NAD), kung saan ang tatlo ay medyo matatag na nauugnay sa mga enzymes (TPF-E 1, lipoamide-E 2 at FAD-E 3), at ang dalawa ay madaling maghiwalay (HS-KoA at NAD).
kanin. 1 Mekanismo ng pagkilos ng pyruvate dehydrogenase complex
E 1 - pyruvate dehydrogenase; E 2 - di-hydrolipoylacetyltransferase; E 3 - dihydrolipoyl dehydrogenase; Ang mga numero sa mga bilog ay nagpapahiwatig ng mga yugto ng proseso.
Ang lahat ng mga enzyme na ito, na may istraktura ng subunit, at mga coenzyme ay isinaayos sa isang solong kumplikado. Samakatuwid, ang mga intermediate na produkto ay mabilis na nakikipag-ugnayan sa isa't isa. Ipinakita na ang mga polypeptide chain ng mga subunit ng dihydrolipoyl acetyltransferase na bumubuo sa complex ay bumubuo sa core ng complex, kung saan matatagpuan ang pyruvate dehydrogenase at dihydrolipoyl dehydrogenase. Karaniwang tinatanggap na ang katutubong enzyme complex ay nabuo sa pamamagitan ng self-assembly.
Ang pangkalahatang reaksyon na na-catalyze ng pyruvate dehydrogenase complex ay maaaring kinakatawan bilang mga sumusunod:
Pyruvate + NAD + + HS-CoA - > Acetyl-CoA + NADH + H + + CO 2.
Ang reaksyon ay sinamahan ng isang makabuluhang pagbaba sa karaniwang libreng enerhiya at halos hindi maibabalik.
Ang acetyl-CoA na nabuo sa panahon ng oxidative decarboxylation ay sumasailalim sa karagdagang oksihenasyon sa pagbuo ng CO 2 at H 2 O. Ang kumpletong oksihenasyon ng acetyl-CoA ay nangyayari sa tricarboxylic acid cycle (Krebs cycle). Ang prosesong ito, pati na rin ang oxidative decarboxylation ng pyruvate, ay nangyayari sa mitochondria ng mga selula.
2 .1.3 IkottricarbonicmaasimT (ikot Crebsa, kudyapimakapal ikot) ay ang gitnang bahagi ng pangkalahatang landas ng catabolism, isang cyclic biochemical aerobic na proseso kung saan ang conversion ng dalawa at tatlong carbon compound ay nabuo bilang mga intermediate na produkto sa mga buhay na organismo sa panahon ng pagkasira ng carbohydrates, fats at proteins ay nangyayari sa CO 2. Sa kasong ito, ang inilabas na hydrogen ay ipinadala sa tissue respiration chain, kung saan ito ay higit na na-oxidized sa tubig, direktang nakikilahok sa synthesis ng unibersal na mapagkukunan ng enerhiya - ATP.
Ang Krebs cycle ay isang mahalagang hakbang sa paghinga ng lahat ng mga cell na gumagamit ng oxygen, ang intersection ng maraming metabolic pathway sa katawan. Bilang karagdagan sa makabuluhang papel ng enerhiya, ang cycle ay mayroon ding makabuluhang plastic function, iyon ay, ito ay isang mahalagang pinagmumulan ng mga precursor molecule, kung saan, sa panahon ng iba pang biochemical transformations, ang mga compound na mahalaga para sa buhay ng cell ay na-synthesize, tulad ng amino acids, carbohydrates, fatty acids, atbp.
Ikot ng pagbabago limonmga acid sa mga buhay na selula ay natuklasan at pinag-aralan ng German biochemist na si Sir Hans Krebs, para sa gawaing ito siya (kasama si F. Lipman) ay iginawad sa Nobel Prize (1953).
Sa mga eukaryote, ang lahat ng mga reaksyon ng siklo ng Krebs ay nangyayari sa loob ng mitochondria, at ang mga enzyme na nag-catalyze sa kanila, maliban sa isa, ay nasa isang libreng estado sa mitochondrial matrix, maliban sa succinate dehydrogenase, na naisalokal sa panloob na mitochondrial membrane, na naka-embed sa ang lipid bilayer. Sa prokaryotes, ang mga reaksyon ng cycle ay nagaganap sa cytoplasm.
Ang pangkalahatang equation para sa isang rebolusyon ng Krebs cycle ay:
Acetyl-CoA > 2CO 2 + CoA + 8e?
Regulasyon ikotA:
Ang siklo ng Krebs ay kinokontrol "sa pamamagitan ng isang negatibong mekanismo ng feedback" sa pagkakaroon ng isang malaking halaga ng mga substrate (acetyl-CoA, oxaloacetate), ang cycle ay aktibong gumagana, at kapag mayroong labis na mga produkto ng reaksyon (NAD, ATP), ito ay inhibited. Ang regulasyon ay isinasagawa din sa tulong ng mga hormone; ang pangunahing mapagkukunan ng acetyl-CoA ay glucose, samakatuwid ang mga hormone na nagtataguyod ng aerobic breakdown ng glucose ay nakakatulong sa paggana ng Krebs cycle. Ang mga hormone na ito ay:
· insulin;
· adrenaline.
Pinasisigla ng glucagon ang glucose synthesis at pinipigilan ang mga reaksyon sa siklo ng Krebs.
Bilang isang patakaran, ang gawain ng siklo ng Krebs ay hindi nagambala dahil sa mga reaksyon ng anaplerotic na muling pinupunan ang ikot ng mga substrate:
Pyruvate + CO 2 + ATP = Oxaloacetate (Krebs Cycle substrate) + ADP + Fn.
Trabaho ATP synthase
Ang proseso ng oxidative phosphorylation ay isinasagawa ng ikalimang complex ng mitochondrial respiratory chain - Proton ATP synthase, na binubuo ng 9 na mga subunit ng 5 uri:
Ang 3 subunits (d,e,f) ay nag-aambag sa integridad ng ATP synthase
· Ang isang subunit ay ang pangunahing functional unit. Mayroon itong 3 conformation:
· L-conformation - nakakabit ng ADP at Phosphate (ipasok ang mitochondrion mula sa cytoplasm gamit ang mga espesyal na carrier)
T-conformation - ang pospeyt ay sumasali sa ADP at nabuo ang ATP
· O-conformation - Nahati ang ATP mula sa b-subunit at inilipat sa b-subunit.
· Upang mabago ng isang subunit ang conform nito, kinakailangan ang isang hydrogen proton, dahil ang conformation ay nagbabago ng 3 beses, 3 hydrogen proton ang kinakailangan. Ang mga proton ay pumped mula sa intermembrane space ng mitochondria sa ilalim ng impluwensya ng electrochemical potential.
· ang b-subunit ay nagdadala ng ATP sa transporter ng lamad, na "nagtatapon" ng ATP sa cytoplasm. Bilang kapalit, ang parehong transporter ay nagdadala ng ADP mula sa cytoplasm. Ang panloob na lamad ng mitochondria ay naglalaman din ng isang Phosphate transporter mula sa cytoplasm patungo sa mitochondrion, ngunit isang hydrogen proton ay kinakailangan para sa operasyon nito. Ang ganitong mga transporter ay tinatawag na translocases.
Kabuuan labasan
Upang ma-synthesize ang 1 molekula ng ATP, kinakailangan ang 3 proton.
Inhibitor oxidative phosphorylation
Hinaharang ng mga inhibitor ang V complex:
· Oligomycin - harangan ang mga proton channel ng ATP synthase.
· Atractyloside, cyclophylline - block translocases.
Mga disconnector oxidative phosphorylation
Mga disconnector- mga lipophilic na sangkap na maaaring tumanggap ng mga proton at ilipat ang mga ito sa pamamagitan ng panloob na lamad ng mitochondria, na lumalampas sa V complex (proton channel nito). Mga Disconnector:
· Natural- mga produkto ng lipid peroxidation, long-chain fatty acids; malalaking dosis ng mga thyroid hormone.
· Artipisyal- dinitrophenol, eter, bitamina K derivatives, anesthetics.
2.2 Substrate phosphorylation
Substrate A tumpakposporil At gumagala ( biochemical), synthesis ng mga compound ng phosphorus na mayaman sa enerhiya dahil sa enerhiya ng redox reactions ng glycolysis (catalyzed ng phosphoglyceraldehyde dehydrogenase at enolase) at sa panahon ng oksihenasyon ng a-ketoglutaric acid sa tricarboxylic acid cycle (sa ilalim ng pagkilos ng a-ketoglutarate dehydrogenase at succinate thiokinase). Ang mga kaso ng S. f. sa panahon ng oksihenasyon ng pyruvic acid.C. f., sa kaibahan sa phosphorylation sa electron transport chain, ay hindi hinahadlangan ng mga "uncoupling" na lason (halimbawa, dinitrophenol) at hindi nauugnay sa pag-aayos ng mga enzyme sa mitochondrial membranes. Kontribusyon ng S. f. ang kontribusyon sa cellular ATP pool sa ilalim ng aerobic na mga kondisyon ay makabuluhang mas mababa kaysa sa kontribusyon ng phosphorylation sa electron transport chain.
Kabanata 3. Mga paraan ng paggamit ng ATP
3.1 Papel sa cell
Ang pangunahing papel ng ATP sa katawan ay nauugnay sa pagbibigay ng enerhiya para sa maraming biochemical reactions. Bilang carrier ng dalawang high-energy bond, ang ATP ay nagsisilbing direktang pinagkukunan ng enerhiya para sa maraming prosesong biochemical at pisyolohikal na umuubos ng enerhiya. Ang lahat ng ito ay mga reaksyon ng synthesis ng mga kumplikadong sangkap sa katawan: ang pagpapatupad ng aktibong paglipat ng mga molekula sa pamamagitan ng mga biological na lamad, kabilang ang paglikha ng isang potensyal na elektrikal na transmembrane; pagpapatupad ng pag-urong ng kalamnan.
Tulad ng nalalaman sa bioenergy ng mga nabubuhay na organismo, dalawang pangunahing punto ang mahalaga:
a) ang enerhiya ng kemikal ay nakaimbak sa pamamagitan ng pagbuo ng ATP kasama ng mga exergonic catabolic na reaksyon ng oksihenasyon ng mga organikong substrate;
b) ang enerhiya ng kemikal ay ginagamit sa pamamagitan ng pagkasira ng ATP, kasama ng mga endergonic na reaksyon ng anabolismo at iba pang mga proseso na nangangailangan ng enerhiya.
Ang tanong ay lumitaw kung bakit tinutupad ng molekula ng ATP ang pangunahing papel nito sa bioenergetics. Upang malutas ito, isaalang-alang ang istraktura ng ATP Istruktura ATP - (sa pH 7,0 tetracharge anion) .
Ang ATP ay isang thermodynamically unstable compound. Ang kawalang-tatag ng ATP ay tinutukoy, una, sa pamamagitan ng electrostatic repulsion sa rehiyon ng isang kumpol ng mga negatibong singil ng parehong pangalan, na humahantong sa pag-igting sa buong molekula, ngunit ang bono ay pinakamalakas - P - O - P, at pangalawa, sa pamamagitan ng isang tiyak na resonance. Alinsunod sa huling kadahilanan, mayroong kumpetisyon sa pagitan ng mga atomo ng phosphorus para sa hindi nakabahaging mga mobile na elektron ng atom ng oxygen na matatagpuan sa pagitan nila, dahil ang bawat atom ng posporus ay may bahagyang positibong singil dahil sa makabuluhang impluwensya ng electron-acceptor ng P=O at P - O- mga pangkat. Kaya, ang posibilidad ng pagkakaroon ng ATP ay tinutukoy ng pagkakaroon ng sapat na dami ng enerhiya ng kemikal sa molekula upang mabayaran ang mga physicochemical stress na ito. Ang molekula ng ATP ay naglalaman ng dalawang phosphoanhydride (pyrophosphate) na mga bono, ang hydrolysis na kung saan ay sinamahan ng isang makabuluhang pagbaba sa libreng enerhiya (sa pH 7.0 at 37 o C).
ATP + H 2 O = ADP + H 3 PO 4 G0I = - 31.0 KJ/mol.
ADP + H 2 O = AMP + H 3 PO 4 G0I = - 31.9 KJ/mol.
Ang isa sa mga pangunahing problema ng bioenergy ay ang biosynthesis ng ATP, na sa buhay na kalikasan ay nangyayari sa pamamagitan ng phosphorylation ng ADP.
Ang Phosphorylation ng ADP ay isang endergonic na proseso at nangangailangan ng mapagkukunan ng enerhiya. Tulad ng nabanggit kanina, dalawang naturang mapagkukunan ng enerhiya ang nangingibabaw sa kalikasan - solar energy at ang kemikal na enerhiya ng mga pinababang organic compound. Ang mga berdeng halaman at ilang mikroorganismo ay may kakayahang baguhin ang enerhiya ng hinihigop na light quanta sa enerhiya ng kemikal, na ginugugol sa phosphorylation ng ADP sa magaan na yugto ng photosynthesis. Ang prosesong ito ng pagbabagong-buhay ng ATP ay tinatawag na photosynthetic phosphorylation. Ang pagbabagong-anyo ng enerhiya ng oksihenasyon ng mga organikong compound sa macroenergetic bond ng ATP sa ilalim ng aerobic na mga kondisyon ay nangyayari pangunahin sa pamamagitan ng oxidative phosphorylation. Ang libreng enerhiya na kinakailangan para sa pagbuo ng ATP ay nabuo sa respiratory oxidative chain ng mitochondria.
Ang isa pang uri ng ATP synthesis ay kilala, na tinatawag na substrate phosphorylation. Sa kaibahan sa oxidative phosphorylation na nauugnay sa paglipat ng elektron, ang donor ng activated phosphoryl group (- PO3 H2), na kinakailangan para sa pagbabagong-buhay ng ATP, ay mga intermediate ng mga proseso ng glycolysis at ang tricarboxylic acid cycle. Sa lahat ng mga kasong ito, ang mga proseso ng oxidative ay humahantong sa pagbuo ng mga high-energy compound: 1,3-diphosphoglycerate (glycolysis), succinyl-CoA (tricarboxylic acid cycle), na, kasama ang pakikilahok ng naaangkop na mga enzyme, ay may kakayahang folylating ADP at bumubuo ng ATP. Ang pagbabagong-anyo ng enerhiya sa antas ng substrate ay ang tanging paraan ng synthesis ng ATP sa mga anaerobic na organismo. Ang prosesong ito ng ATP synthesis ay nagpapahintulot sa iyo na mapanatili ang matinding trabaho ng mga kalamnan ng kalansay sa mga panahon ng gutom sa oxygen. Dapat tandaan na ito ang tanging landas para sa synthesis ng ATP sa mga mature na pulang selula ng dugo na walang mitochondria.
Ang isang partikular na mahalagang papel sa bioenergetics ng cell ay nilalaro ng adenyl nucleotide, kung saan ang dalawang phosphoric acid residues ay nakakabit. Ang sangkap na ito ay tinatawag na adenosine triphosphoric acid (ATP). Ang enerhiya ay nakaimbak sa mga bono ng kemikal sa pagitan ng mga residue ng phosphoric acid ng molekula ng ATP, na inilalabas kapag ang organikong phosphorite ay pinaghiwalay:
ATP= ADP+P+E,
kung saan ang F ay isang enzyme, ang E ay nagpapalaya ng enerhiya. Sa reaksyong ito, nabuo ang adenosine phosphoric acid (ADP) - ang natitira sa molekula ng ATP at organic phosphate. Ang lahat ng mga cell ay gumagamit ng enerhiya ng ATP para sa mga proseso ng biosynthesis, paggalaw, paggawa ng init, mga nerve impulses, luminescence (halimbawa, luminescent bacteria), iyon ay, para sa lahat ng mahahalagang proseso.
Ang ATP ay isang unibersal na biological energy accumulator. Ang liwanag na enerhiya na nakapaloob sa pagkain na natupok ay nakaimbak sa mga molekula ng ATP.
Ang supply ng ATP sa cell ay maliit. Kaya, ang reserbang ATP sa kalamnan ay sapat na para sa 20 - 30 contraction. Sa matinding, ngunit panandaliang trabaho, ang mga kalamnan ay gumagana nang eksklusibo dahil sa pagkasira ng ATP na nakapaloob sa kanila. Pagkatapos ng trabaho, ang isang tao ay humihinga nang mabigat - sa panahong ito, ang mga karbohidrat at iba pang mga sangkap ay nasira (naipon ang enerhiya) at ang supply ng ATP sa mga selula ay naibalik.
Ang papel ng ATP bilang isang transmitter sa mga synapses ay kilala rin.
3.2 Papel sa paggana ng enzyme
Ang buhay na selula ay isang sistemang kemikal na malayo sa ekwilibriyo: pagkatapos ng lahat, ang paglapit ng isang buhay na sistema sa ekwilibriyo ay nangangahulugan ng pagkawatak-watak at kamatayan nito. Ang produkto ng bawat enzyme ay kadalasang mabilis na nauubos dahil ginagamit ito bilang substrate ng isa pang enzyme sa metabolic pathway. Higit sa lahat, ang isang malaking bilang ng mga reaksyong enzymatic ay kinabibilangan ng pagkasira ng ATP sa ADP at hindi organikong pospeyt. Upang ito ay maging posible, ang ATP pool naman ay dapat mapanatili sa isang antas na malayo sa ekwilibriyo, upang ang ratio ng konsentrasyon ng ATP sa konsentrasyon ng mga produktong hydrolysis nito ay mataas. Kaya, ang ATP pool ay gumaganap ng papel ng isang "baterya" na nagpapanatili ng patuloy na paglipat ng enerhiya at mga atomo sa cell kasama ang mga metabolic pathway na tinutukoy ng pagkakaroon ng mga enzyme.
Kaya, isaalang-alang natin ang proseso ng ATP hydrolysis at ang epekto nito sa paggana ng mga enzyme. Isipin natin ang isang tipikal na proseso ng biosynthetic kung saan ang dalawang monomer - A at B - ay dapat pagsamahin sa isa't isa sa isang reaksyon ng pag-aalis ng tubig (tinatawag ding condensation), na sinamahan ng paglabas ng tubig:
A - N + B - OH - AB + H2O
Ang kabaligtaran na reaksyon, na tinatawag na hydrolysis, kung saan ang isang molekula ng tubig ay bumabagsak sa isang covalently bonded compound A - B, ay halos palaging magiging masiglang pabor. Ito ay nangyayari, halimbawa, sa panahon ng hydrolytic breakdown ng mga protina, nucleic acid at polysaccharides sa mga subunit.
Ang pangkalahatang diskarte kung saan nabuo ang mga cell A - B na may A - H at B - OH ay may kasamang multi-stage na pagkakasunud-sunod ng mga reaksyon, bilang isang resulta kung saan n Ang pag-uugnay ng energetically unfavorable synthesis ng mga kinakailangang compound na may balanseng kapaki-pakinabang na reaksyon ay nangyayari.
Ang ATP hydrolysis ba ay tumutugma sa isang malaking negatibong halaga? G, samakatuwid, ang ATP hydrolysis ay madalas na gumaganap ng papel ng isang masiglang kanais-nais na reaksyon, dahil sa kung saan ang mga reaksyon ng intracellular biosynthesis ay isinasagawa.
Sa landas mula sa A - H at B - OH - A - B, na nauugnay sa ATP hydrolysis, ang enerhiya ng hydrolysis ay unang nag-convert ng B - OH sa isang high-energy intermediate, na pagkatapos ay direktang tumutugon sa A - H, na bumubuo ng A - B .
1) B - OH + ATP - B - B - P + ADP
2) A - N + B - O - R - A - B + R
Dahil ang intermediate compound B - O - P na nabuo sa panahon ng reaksyon ay nawasak muli, ang pangkalahatang mga reaksyon ay maaaring ilarawan gamit ang mga sumusunod na equation:
3) A-N + B - OH - A - B at ATP - ADP + P
Ang una, energetically hindi kanais-nais na reaksyon ay lumalabas na posible dahil ito ay nauugnay sa pangalawang, energetically paborable na reaksyon (ATP hydrolysis). Ang isang halimbawa ng pinagsamang biosynthetic na reaksyon ng ganitong uri ay ang synthesis ng amino acid glutamine.
Ang halaga ng G ng ATP hydrolysis sa ADP at inorganic phosphate ay nakasalalay sa konsentrasyon ng lahat ng tumutugon na sangkap at kadalasan para sa mga kondisyon ng cell ay nasa saklaw mula - 11 hanggang - 13 kcal / mol. Ang reaksyon ng hydrolysis ng ATP ay maaaring magamit sa wakas upang magsagawa ng isang thermodynamically hindi kanais-nais na reaksyon na may halaga ng G na humigit-kumulang +10 kcal/mol, siyempre sa pagkakaroon ng naaangkop na pagkakasunud-sunod ng reaksyon. Gayunpaman, para sa maraming mga reaksyon ng biosynthesis kahit na ito ay hindi sapat? G = - 13 kcal/mol. Sa mga ito at iba pang mga kaso, ang ATP hydrolysis pathway ay binago upang ang AMP at PP (pyrophosphate) ay mabuo muna. Sa susunod na yugto, ang pyrophosphate ay sumasailalim din sa hydrolysis; ang kabuuang libreng pagbabago ng enerhiya ng buong proseso ay humigit-kumulang - 26 kcal/mol.
Paano ginagamit ang enerhiya mula sa hydrolysis ng pyrophosphate sa mga biosynthetic na reaksyon? Ang isa sa mga paraan ay maaaring maipakita sa pamamagitan ng halimbawa ng synthesis sa itaas ng tambalang A - B na may A - H at B - OH. Sa tulong ng naaangkop na enzyme, ang B - OH ay maaaring tumugon sa ATP at maging high-energy compound B - O - P - P. Ngayon ang reaksyon ay binubuo ng tatlong yugto:
1) B - OH + ATP - B - B - P - P + AMP
2) A - N + B - O - R - R - A - B + RR
3) PP + H2O - 2P
Ang kabuuang reaksyon ay maaaring ilarawan bilang mga sumusunod:
A - H + B - OH - A - B at ATP + H2O - AMP + 2P
Dahil ang enzyme ay palaging nagpapabilis sa catalyzed na reaksyon sa parehong pasulong at pabalik na direksyon, ang tambalang A - B ay maaaring mabulok sa pamamagitan ng pagtugon sa pyrophosphate (reaksyon, ang kabaligtaran ng hakbang 2). Gayunpaman, ang masiglang pabor na reaksyon ng pyrophosphate hydrolysis (hakbang 3) ay nakakatulong na mapanatili ang katatagan ng A-B compound sa pamamagitan ng pagpapanatiling napakababa ng konsentrasyon ng pyrophosphate (pinipigilan nitong mangyari ang reverse reaction mula sa hakbang 2). Kaya, ang enerhiya ng pyrophosphate hydrolysis ay nagsisiguro na ang reaksyon ay nagpapatuloy sa pasulong na direksyon. Ang isang halimbawa ng isang mahalagang biosynthetic reaksyon ng ganitong uri ay ang synthesis ng polynucleotides.
3.3 Tungkulin sa synthesis ng DNA at RNA at mga protina
Sa lahat ng kilalang organismo, ang mga deoxyribonucleotides na bumubuo sa DNA ay na-synthesize sa pamamagitan ng pagkilos ng ribonucleotide reductase (RNR) enzymes sa kaukulang ribonucleotides. Binabawasan ng mga enzyme na ito ang residue ng asukal na otribose sa deoxyribose sa pamamagitan ng pag-alis ng oxygen mula sa 2" hydroxyl group, mga substrate na ribonucleoside diphosphate at mga produktong deoxyribonucleoside diphosphate. sa panahon ng reaksyon Ang PHP enzyme ay pinoproseso sa pamamagitan ng reaksyon sa thioredoxin o glutaredoxin.
Ang regulasyon ng RHP at mga kaugnay na enzyme ay nagpapanatili ng balanse sa isa't isa. Ang napakababang konsentrasyon ay pumipigil sa synthesis ng DNA at pag-aayos ng DNA at nakamamatay sa cell, habang ang abnormal na ratio ay mutagenic dahil sa tumaas na posibilidad ng pagsasama ng DNA polymerase sa panahon ng DNA synthesis.
Sa panahon ng synthesis ng RNA nucleic acid, ang adenosine na nagmula sa ATP ay isa sa apat na nucleotides na direktang isinama sa mga molekula ng RNA ng RNA polymerase. Enerhiya, ang polymerization na ito ay nangyayari sa pag-aalis ng pyrophosphate (dalawang grupo ng pospeyt). Ang prosesong ito ay katulad sa DNA biosynthesis, maliban na ang ATP ay nabawasan sa deoxyribonucleotide dATP, bago isama sa DNA.
SA synthesis ardilya. Gumagamit ang Aminoacyl-tRNA synthetases ng mga enzyme ng ATP bilang pinagmumulan ng enerhiya upang ikabit ang isang molekula ng tRNA sa tiyak nitong amino acid, na bumubuo ng isang aminoacyl-tRNA, na handang ilipat sa mga ribosom. Ang enerhiya ay nagiging available sa pamamagitan ng hydrolysis ng ATP ng adenosine monophosphate (AMP), na nag-aalis ng dalawang grupo ng pospeyt.
Ang ATP ay ginagamit para sa maraming cellular function, kabilang ang transport work ng mga gumagalaw na substance sa mga cell membrane. Ginagamit din ito para sa mekanikal na trabaho, na nagbibigay ng enerhiya na kailangan para sa pag-urong ng kalamnan. Nagbibigay ito ng enerhiya hindi lamang sa kalamnan ng puso (para sa sirkulasyon ng dugo) at mga kalamnan ng kalansay (halimbawa, para sa kabuuang paggalaw ng katawan), kundi pati na rin sa mga chromosome at flagella upang maisagawa nila ang kanilang maraming tungkulin. Ang pangunahing papel ng ATP ay sa gawaing kemikal, na nagbibigay ng kinakailangang enerhiya para sa synthesis ng ilang libong uri ng macromolecules na dapat umiral ang isang cell.
Ginagamit din ang ATP bilang on-off switch para makontrol ang mga reaksiyong kemikal at magpadala ng impormasyon. Ang hugis ng mga kadena ng protina na gumagawa ng mga bloke ng gusali at iba pang mga istruktura na ginagamit sa buhay ay pangunahing tinutukoy ng mahinang mga bono ng kemikal na madaling mawala at muling ayusin. Ang mga circuit na ito ay maaaring paikliin, pahabain, at baguhin ang hugis bilang tugon sa input o output ng enerhiya. Ang mga pagbabago sa mga kadena ay nagbabago sa hugis ng protina at maaari ring baguhin ang paggana nito o maging sanhi ng pagiging aktibo o hindi aktibo.
Ang mga molekula ng ATP ay maaaring magbigkis sa isang bahagi ng isang molekula ng protina, na nagiging sanhi ng isa pang bahagi ng parehong molekula na mag-slide o bahagyang gumalaw na nagiging sanhi ng pagbabago nito sa conformation, at hindi aktibo ang molekula. Kapag naalis na, nagiging sanhi ng ATP na bumalik ang protina sa orihinal nitong anyo, at sa gayon ito ay gumaganang muli.
Maaaring ulitin ang cycle hanggang sa bumalik ang molekula, na epektibong kumikilos bilang parehong on/off switch. Parehong ang pagdaragdag ng phosphorus (phosphorylation) at ang pag-alis ng phosphorus mula sa isang protina (dephosphorylation) ay maaaring magsilbi bilang alinman sa isang on o off switch.
3.4 Iba pang mga function ng ATP
Tungkulin V metabolismo, synthesis At aktibo transportasyon
Kaya, ang ATP ay naglilipat ng enerhiya sa pagitan ng spatially separated metabolic reactions. Ang ATP ay ang pangunahing pinagkukunan ng enerhiya para sa karamihan ng mga cellular function. Kabilang dito ang synthesis ng macromolecules, kabilang ang DNA at RNA, at mga protina. Ang ATP ay gumaganap din ng isang mahalagang papel sa transportasyon ng mga macromolecule sa mga lamad ng cell, tulad ng exocytosis at endocytosis.
Tungkulin V istraktura mga selula At paggalaw
Ang ATP ay kasangkot sa pagpapanatili ng cellular na istraktura sa pamamagitan ng pagpapadali sa pagpupulong at pag-disassembly ng mga elemento ng cytoskeletal. Dahil sa prosesong ito, kinakailangan ang ATP para sa contraction ng actin filament at kailangan ang myosin para sa muscle contraction. Ang huling prosesong ito ay isa sa mga pangunahing pangangailangan ng enerhiya ng mga hayop at mahalaga para sa paggalaw at paghinga.
Tungkulin V hudyat mga sistema
Saextracellularhudyatmga sistema
Ang ATP ay isa ring molekula ng pagbibigay ng senyas. Ang ATP, ADP, o adenosine ay kinikilala bilang mga purinergic receptor. Ang mga purinoreceptor ay maaaring ang pinaka-masaganang mga receptor sa mga tisyu ng mammalian.
Sa mga tao, ang papel na ito ng pagbibigay ng senyas ay mahalaga sa parehong central at peripheral nervous system. Ang aktibidad ay nakasalalay sa pagpapalabas ng ATP mula sa mga synapses, axon at glia sa pamamagitan ng purinergic activation ng mga receptor ng lamad.
Saintracellularhudyatmga sistema
Ang ATP ay kritikal sa mga proseso ng signal transduction. Ginagamit ito ng mga kinase bilang pinagmumulan ng mga grupo ng pospeyt sa kanilang reaksyon sa paglilipat ng pospeyt. Ang kinase sa mga suporta tulad ng mga protina ng lamad o lipid ay isang karaniwang anyo ng signal. Ang phosphorylation ng protina sa pamamagitan ng kinases ay maaaring i-activate ang cascade na ito, tulad ng mitogen-activated protein kinase cascade.
Ang ATP ay ginagamit din ng adenylate cyclase at na-convert sa pangalawang messenger molecule na tinatawag na AMP, na kasangkot sa pag-trigger ng mga signal ng calcium upang palabasin ang calcium mula sa mga intracellular na tindahan. [38] Ang anyo ng signal na ito ay partikular na mahalaga sa paggana ng utak, bagama't ito ay kasangkot sa regulasyon ng maraming iba pang mga proseso ng selula.
Konklusyon
1. Adenosine triphosphate - isang nucleotide, ay gumaganap ng isang napakahalagang papel sa pagpapalitan ng enerhiya at mga sangkap sa mga organismo; Una sa lahat, ang tambalan ay kilala bilang isang unibersal na pinagmumulan ng enerhiya para sa lahat ng mga prosesong biochemical na nagaganap sa mga buhay na sistema. Sa kemikal, ang ATP ay ang triphosphate ester ng adenosine, na isang derivative ng adenine at ribose. Ang istraktura ng ATP ay katulad ng adenine nucleotide na bahagi ng RNA, tanging sa halip na isang phosphoric acid, ang ATP ay naglalaman ng tatlong phosphoric acid residues. Ang mga cell ay hindi maaaring maglaman ng mga acid sa kapansin-pansing dami, ngunit ang kanilang mga asin lamang. Samakatuwid, ang phosphoric acid ay pumapasok sa ATP bilang isang nalalabi (sa halip na ang pangkat ng OH ng acid ay mayroong negatibong sisingilin na oxygen atom).
2. Sa katawan, ang ATP ay synthesize sa pamamagitan ng phosphorylation ng ADP:
ADP + H 3 PO 4 + enerhiya> ATP + H 2 O.
Ang Phosphorylation ng ADP ay posible sa dalawang paraan: substrate phosphorylation at oxidative phosphorylation (gamit ang enerhiya ng oxidizing substances).
Oxidative phosphorylation - isa sa pinakamahalagang bahagi ng cellular respiration, na humahantong sa paggawa ng enerhiya sa anyo ng ATP. Ang mga substrate para sa oxidative phosphorylation ay ang mga produkto ng pagkasira ng mga organikong compound - mga protina, taba at carbohydrates. Ang proseso ng oxidative phosphorylation ay nagaganap sa cristae ng mitochondria.
Substrate A tumpakposporil At gumagala ( biochemical), synthesis ng mga compound na posporus na mayaman sa enerhiya dahil sa enerhiya ng redox reactions ng glycolysis at sa panahon ng oksihenasyon ng a-ketoglutaric acid sa tricarboxylic acid cycle.
3. Ang pangunahing papel ng ATP sa katawan ay nauugnay sa pagbibigay ng enerhiya para sa maraming biochemical reactions. Bilang carrier ng dalawang high-energy bond, ang ATP ay nagsisilbing direktang pinagkukunan ng enerhiya para sa maraming prosesong biochemical at pisyolohikal na umuubos ng enerhiya. Sa bioenergetics ng mga buhay na organismo, ang mga sumusunod ay mahalaga: ang enerhiya ng kemikal ay iniimbak sa pamamagitan ng pagbuo ng ATP, kasama ng mga exergonic catabolic na reaksyon ng oksihenasyon ng mga organikong substrate; Ang enerhiya ng kemikal ay ginagamit sa pamamagitan ng pagkasira ng ATP, kasama ng mga endergonic na reaksyon ng anabolismo at iba pang mga proseso na nangangailangan ng enerhiya.
4. Sa pagtaas ng load (halimbawa, sa short-distance running), ang mga kalamnan ay gumagana nang eksklusibo dahil sa supply ng ATP. Sa mga selula ng kalamnan, ang reserbang ito ay sapat na para sa ilang dosenang mga contraction, at pagkatapos ay dapat na mapunan ang halaga ng ATP. Ang synthesis ng ATP mula sa ADP at AMP ay nangyayari dahil sa enerhiya na inilabas sa panahon ng pagkasira ng carbohydrates, lipids at iba pang mga sangkap. Ang pagsasagawa ng mental na gawain ay nangangailangan din ng malaking halaga ng ATP. Para sa kadahilanang ito, ang mga taong may mental na trabaho ay nangangailangan ng mas mataas na halaga ng glucose, ang pagkasira nito ay nagsisiguro sa synthesis ng ATP.
Bilang karagdagan sa enerhiya, ang ATP ay gumaganap ng maraming iba pang pantay na mahalagang pag-andar sa katawan:
· Kasama ng iba pang nucleoside triphosphate, ang ATP ang panimulang produkto sa synthesis ng mga nucleic acid.
· Bilang karagdagan, ang ATP ay gumaganap ng isang mahalagang papel sa regulasyon ng maraming mga biochemical na proseso. Ang pagiging isang allosteric effector ng isang bilang ng mga enzymes, ATP, pagsali sa kanilang mga regulatory centers, pinahuhusay o pinipigilan ang kanilang aktibidad.
· Ang ATP ay isa ring direktang precursor para sa synthesis ng cyclic adenosine monophosphate, isang pangalawang messenger ng hormonal signal transmission sa cell.
Ang papel ng ATP bilang isang transmitter sa mga synapses ay kilala rin.
Bibliograpiya
1. Lemeza, N.A. Isang manwal sa biology para sa mga aplikante sa mga unibersidad / L.V. Kamlyuk N.D. Lisov. - Mn.: Unipress, 2011 - 624 p.
2. Lodish, H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. Molecular Cell Biology, 5th ed. - New York: W. H. Freeman, 2004.
3. Romanovsky, Yu.M. Molecular energy converters ng mga buhay na selula. Proton ATP synthase - isang umiikot na molecular motor / Yu.M. Romanovsky A.N. Tikhonov // UFN. - 2010. - T.180. - P.931 - 956.
4. Voet D, Voet JG. Biochemistry Vol 1 3rd ed. - Wiley: Hoboken, NJ. - N-Y: W. H. Freeman and Company, 2002. - 487 rub.
5. Pangkalahatang kimika. Biophysical chemistry. Chemistry ng mga biogenic na elemento. M.: Higher School, 1993
6. Vershubsky, A.V. Biophysics. / A.V. Vershubsky, V.I. Priklonsky, A.N. Tikhonov. - M: 471-481.
7. Alberts B. Molecular biology ng mga cell sa 3 volume. / Alberts B., Bray D., Lewis J. et al M.: Mir, 1994.1558 p.
8. Nikolaev A.Ya. Biological chemistry - M.: Medical Information Agency LLC, 1998.
9. Berg, J. M. Biochemistry, internasyonal na edisyon. / Berg, J. M, Tymoczko, J. L, Stryer, L. - New York: WH Freeman, 2011; p287.
10. Knorre D.G. Biyolohikal na kimika: Teksbuk. para sa kimika, biol. At honey. espesyalista. mga unibersidad - 3rd ed., rev. / Knorre D.G., Mysina S.D. - M.: Mas mataas. paaralan, 2000. - 479 p.: may sakit.
11. Eliot, V. Biochemistry at molecular biology / V. Eliot, D. Eliot. - M.: Publishing House ng Research Institute of Biomedical Chemistry ng Russian Academy of Medical Sciences, LLC "Materik-alpha", 1999, - 372 p.
12. Shina CL, K., 7 Areieh, W. Sa Energetics ng ATP Hydrolysis sa Solusyon. Journal Of Physical Chemistry B, 113 (47), (2009).
13. Berg, J. M. Biochemistry / J. M. Berg: J. L. Tymoczko, L. Stryer. - N-Y: W. H. Freeman and Company, 2002. - 1514 p.
...Mga katulad na dokumento
Mga organikong compound sa katawan ng tao. Istraktura, pag-andar at pag-uuri ng mga protina. Mga nucleic acid (polynucleotides), mga tampok na istruktura at katangian ng RNA at DNA. Carbohydrates sa kalikasan at sa katawan ng tao. Ang mga lipid ay mga taba at mga sangkap na tulad ng taba.
abstract, idinagdag 09/06/2009
Ang proseso ng synthesis ng protina at ang kanilang papel sa buhay ng mga nabubuhay na organismo. Mga pag-andar at kemikal na katangian ng mga amino acid. Mga dahilan para sa kanilang kakulangan sa katawan ng tao. Mga uri ng pagkain na naglalaman ng mga mahahalagang acid. Ang mga amino acid ay na-synthesize sa atay.
pagtatanghal, idinagdag noong 10/23/2014
Enerhiya, pag-iimbak at mga function ng pagbuo ng suporta ng carbohydrates. Mga katangian ng monosaccharides bilang pangunahing pinagkukunan ng enerhiya sa katawan ng tao; glucose. Ang mga pangunahing kinatawan ng disaccharides; sucrose. Polysaccharides, pagbuo ng almirol, metabolismo ng karbohidrat.
ulat, idinagdag noong 04/30/2010
Metabolic functions sa katawan: pagbibigay ng mga organo at sistema ng enerhiya na nabuo sa panahon ng pagkasira ng mga sustansya; ginagawang mga bloke ng gusali ang mga molekula ng pagkain; pagbuo ng mga nucleic acid, lipid, carbohydrates at iba pang mga bahagi.
abstract, idinagdag 01/20/2009
Ang papel at kahalagahan ng mga protina, taba at carbohydrates para sa normal na kurso ng lahat ng mahahalagang proseso. Komposisyon, istraktura at mga pangunahing katangian ng mga protina, taba at carbohydrates, ang kanilang pinakamahalagang gawain at tungkulin sa katawan. Pangunahing pinagmumulan ng mga sustansyang ito.
pagtatanghal, idinagdag noong 04/11/2013
Mga katangian ng istraktura ng mga molekula ng kolesterol bilang isang mahalagang bahagi ng lamad ng cell. Pag-aaral ng mga mekanismo ng regulasyon ng metabolismo ng kolesterol sa katawan ng tao. Pagsusuri ng mga tampok ng paglitaw ng labis na low-density na lipoprotein sa daluyan ng dugo.
abstract, idinagdag 06/17/2012
Metabolismo ng mga protina, lipid at carbohydrates. Mga uri ng nutrisyon ng tao: omnivorous, hiwalay at low-carbohydrate na nutrisyon, vegetarianism, hilaw na pagkain na pagkain. Ang papel ng mga protina sa metabolismo. Kakulangan ng taba sa katawan. Mga pagbabago sa katawan bilang resulta ng mga pagbabago sa uri ng diyeta.
course work, idinagdag 02/02/2014
Isinasaalang-alang ang pakikilahok ng bakal sa mga proseso ng oxidative at sa synthesis ng collagen. Pagkilala sa kahalagahan ng hemoglobin sa mga proseso ng pagbuo ng dugo. Pagkahilo, igsi ng paghinga at metabolic disorder bilang resulta ng kakulangan sa iron sa katawan ng tao.
pagtatanghal, idinagdag 02/08/2012
Mga katangian ng fluorine at iron. Pang-araw-araw na pangangailangan ng katawan. Mga function ng fluorine sa katawan, impluwensya, nakamamatay na dosis, pakikipag-ugnayan sa iba pang mga sangkap. Iron sa katawan ng tao, ang mga pinagmumulan nito. Mga kahihinatnan ng kakulangan sa iron para sa katawan at labis nito.
pagtatanghal, idinagdag noong 02/14/2017
Ang mga protina bilang pinagmumulan ng pagkain, ang kanilang mga pangunahing tungkulin. Ang mga amino acid ay kasangkot sa paglikha ng mga protina. Istraktura ng polypeptide chain. Mga pagbabagong-anyo ng mga protina sa katawan. Kumpleto at hindi kumpleto ang mga protina. Istraktura ng protina, mga katangian ng kemikal, mga reaksyon ng husay.
Milyun-milyong biochemical reaction ang nagaganap sa alinmang cell ng ating katawan. Ang mga ito ay na-catalyze ng iba't ibang mga enzyme, na kadalasang nangangailangan ng enerhiya. Saan nakukuha ng cell? Ang tanong na ito ay masasagot kung isasaalang-alang natin ang istraktura ng molekula ng ATP - isa sa mga pangunahing mapagkukunan ng enerhiya.
Ang ATP ay isang unibersal na mapagkukunan ng enerhiya
Ang ATP ay nangangahulugang adenosine triphosphate, o adenosine triphosphate. Ang substansiya ay isa sa dalawang pinakamahalagang mapagkukunan ng enerhiya sa anumang cell. Ang istraktura ng ATP at ang biological na papel nito ay malapit na nauugnay. Karamihan sa mga biochemical reaksyon ay maaaring mangyari lamang sa paglahok ng mga molekula ng isang sangkap, ito ay totoo lalo na, gayunpaman, ang ATP ay bihirang direktang kasangkot sa reaksyon: para sa anumang proseso na mangyari, ang enerhiya na tiyak na nilalaman sa adenosine triphosphate ay kinakailangan.
Ang istraktura ng mga molekula ng sangkap ay tulad na ang mga bono na nabuo sa pagitan ng mga grupo ng pospeyt ay nagdadala ng isang malaking halaga ng enerhiya. Samakatuwid, ang mga naturang bono ay tinatawag ding macroergic, o macroenergetic (macro=many, large amount). Ang termino ay unang ipinakilala ng siyentipiko na si F. Lipman, at iminungkahi din niya ang paggamit ng simbolong ̴ upang italaga ang mga ito.
Napakahalaga para sa cell na mapanatili ang isang pare-parehong antas ng adenosine triphosphate. Ito ay totoo lalo na para sa mga selula ng kalamnan at nerve fibers, dahil sila ang pinaka-nakadepende sa enerhiya at nangangailangan ng mataas na nilalaman ng adenosine triphosphate upang maisagawa ang kanilang mga function.
Ang istraktura ng molekula ng ATP
Ang adenosine triphosphate ay binubuo ng tatlong elemento: ribose, adenine at residues

Ribose- isang carbohydrate na kabilang sa pangkat ng pentose. Nangangahulugan ito na ang ribose ay naglalaman ng 5 carbon atoms, na nakapaloob sa isang cycle. Ang Ribose ay kumokonekta sa adenine sa pamamagitan ng isang β-N-glycosidic bond sa 1st carbon atom. Ang mga residue ng phosphoric acid sa 5th carbon atom ay idinagdag din sa pentose.
Ang Adenine ay isang nitrogenous base. Depende sa kung aling nitrogenous base ang nakakabit sa ribose, ang GTP (guanosine triphosphate), TTP (thymidine triphosphate), CTP (cytidine triphosphate) at UTP (uridine triphosphate) ay nakikilala din. Ang lahat ng mga sangkap na ito ay katulad sa istraktura sa adenosine triphosphate at gumaganap ng humigit-kumulang sa parehong mga function, ngunit ang mga ito ay mas karaniwan sa cell.
Mga residu ng phosphoric acid. Ang maximum na tatlong phosphoric acid residues ay maaaring ikabit sa ribose. Kung mayroong dalawa o isa lamang, kung gayon ang sangkap ay tinatawag na ADP (diphosphate) o AMP (monophosphate). Nasa pagitan ng mga residu ng phosphorus na ang mga macroenergetic bond ay natapos, pagkatapos ng pagkalagot kung saan ang 40 hanggang 60 kJ ng enerhiya ay pinakawalan. Kung ang dalawang bono ay nasira, 80, mas madalas - 120 kJ ng enerhiya ang pinakawalan. Kapag nasira ang bond sa pagitan ng ribose at phosphorus residue, 13.8 kJ lang ang inilalabas, kaya dalawa lang ang high-energy bond sa triphosphate molecule (P ̴ P ̴ P), at sa ADP molecule mayroong isa (P ̴ P).
Ito ang mga tampok na istruktura ng ATP. Dahil sa ang katunayan na ang isang macroenergetic bond ay nabuo sa pagitan ng phosphoric acid residues, ang istraktura at pag-andar ng ATP ay magkakaugnay.
Ang istraktura ng ATP at ang biological na papel ng molekula. Mga karagdagang pag-andar ng adenosine triphosphate
Bilang karagdagan sa enerhiya, ang ATP ay maaaring magsagawa ng maraming iba pang mga function sa cell. Kasama ng iba pang mga nucleotide triphosphate, ang triphosphate ay kasangkot sa pagbuo ng mga nucleic acid. Sa kasong ito, ang ATP, GTP, TTP, CTP at UTP ay mga supplier ng nitrogenous base. Ginagamit ang property na ito sa mga proseso at transkripsyon.
Ang ATP ay kinakailangan din para sa paggana ng mga channel ng ion. Halimbawa, ang Na-K channel ay nagbobomba ng 3 sodium molecule palabas ng cell at nagbomba ng 2 potassium molecules sa cell. Ang kasalukuyang ion na ito ay kinakailangan upang mapanatili ang isang positibong singil sa panlabas na ibabaw ng lamad, at sa tulong lamang ng adenosine triphosphate maaaring gumana ang channel. Ang parehong naaangkop sa proton at calcium channels.
Ang ATP ay ang pasimula ng pangalawang messenger cAMP (cyclic adenosine monophosphate) - ang cAMP ay hindi lamang nagpapadala ng signal na natanggap ng mga cell membrane receptor, ngunit ito rin ay isang allosteric effector. Ang mga allosteric effector ay mga sangkap na nagpapabilis o nagpapabagal sa mga reaksyon ng enzymatic. Kaya, pinipigilan ng cyclic adenosine triphosphate ang synthesis ng isang enzyme na nagpapagana sa pagkasira ng lactose sa mga selulang bacterial.
Ang mismong molekula ng adenosine triphosphate ay maaari ding isang allosteric effector. Bukod dito, sa ganitong mga proseso, ang ADP ay kumikilos bilang isang antagonist sa ATP: kung pinabilis ng triphosphate ang reaksyon, pinipigilan ito ng diphosphate, at kabaliktaran. Ito ang mga function at istraktura ng ATP.

Paano nabuo ang ATP sa isang cell?
Ang mga pag-andar at istraktura ng ATP ay tulad na ang mga molekula ng sangkap ay mabilis na ginagamit at nawasak. Samakatuwid, ang triphosphate synthesis ay isang mahalagang proseso sa pagbuo ng enerhiya sa cell.
Mayroong tatlong pinakamahalagang pamamaraan para sa synthesis ng adenosine triphosphate:
1. Substrate phosphorylation.
2. Oxidative phosphorylation.
3. Photophosphorylation.
Ang substrate phosphorylation ay batay sa maraming reaksyon na nagaganap sa cell cytoplasm. Ang mga reaksyong ito ay tinatawag na glycolysis - anaerobic stage Bilang resulta ng 1 cycle ng glycolysis, mula sa 1 molekula ng glucose dalawang molekula ang na-synthesize, na pagkatapos ay ginagamit upang makagawa ng enerhiya, at dalawang ATP ay na-synthesize din.
- C 6 H 12 O 6 + 2ADP + 2Pn --> 2C 3 H 4 O 3 + 2ATP + 4H.
Paghinga ng cell
Ang Oxidative phosphorylation ay ang pagbuo ng adenosine triphosphate sa pamamagitan ng paglilipat ng mga electron sa kahabaan ng chain ng transport electron ng lamad. Bilang resulta ng paglipat na ito, ang isang proton gradient ay nabuo sa isang gilid ng lamad at, sa tulong ng protina integral set ng ATP synthase, ang mga molekula ay binuo. Ang proseso ay nagaganap sa mitochondrial membrane.

Ang pagkakasunud-sunod ng mga yugto ng glycolysis at oxidative phosphorylation sa mitochondria ay bumubuo ng isang karaniwang proseso na tinatawag na respiration. Pagkatapos ng kumpletong cycle, 36 ATP molecules ang nabuo mula sa 1 glucose molecule sa cell.
Photophosphorylation
Ang proseso ng photophosphorylation ay kapareho ng oxidative phosphorylation na may isang pagkakaiba lamang: ang mga reaksyon ng photophosphorylation ay nangyayari sa mga chloroplast ng cell sa ilalim ng impluwensya ng liwanag. Ang ATP ay ginawa sa panahon ng magaan na yugto ng photosynthesis, ang pangunahing proseso ng paggawa ng enerhiya sa mga berdeng halaman, algae at ilang bakterya.
Sa panahon ng photosynthesis, ang mga electron ay dumadaan sa parehong electron transport chain, na nagreresulta sa pagbuo ng isang proton gradient. Ang konsentrasyon ng mga proton sa isang gilid ng lamad ay ang pinagmulan ng ATP synthesis. Ang pagpupulong ng mga molekula ay isinasagawa ng enzyme ATP synthase.

Ang average na cell ay naglalaman ng 0.04% adenosine triphosphate ayon sa timbang. Gayunpaman, ang pinakamataas na halaga ay sinusunod sa mga selula ng kalamnan: 0.2-0.5%.
Mayroong humigit-kumulang 1 bilyong molekula ng ATP sa isang cell.
Ang bawat molekula ay nabubuhay nang hindi hihigit sa 1 minuto.
Ang isang molekula ng adenosine triphosphate ay na-renew 2000-3000 beses sa isang araw.
Sa kabuuan, ang katawan ng tao ay nag-synthesize ng 40 kg ng adenosine triphosphate bawat araw, at sa anumang oras ang reserba ng ATP ay 250 g.

Konklusyon
Ang istraktura ng ATP at ang biological na papel ng mga molekula nito ay malapit na nauugnay. Ang sangkap ay gumaganap ng isang mahalagang papel sa mga proseso ng buhay, dahil ang mataas na enerhiya na mga bono sa pagitan ng mga residu ng pospeyt ay naglalaman ng malaking halaga ng enerhiya. Ang adenosine triphosphate ay gumaganap ng maraming mga function sa cell, at samakatuwid ito ay mahalaga upang mapanatili ang isang pare-pareho ang konsentrasyon ng sangkap. Ang pagkabulok at synthesis ay nangyayari sa mataas na bilis, dahil ang enerhiya ng mga bono ay patuloy na ginagamit sa mga biochemical na reaksyon. Ito ay isang mahalagang sangkap para sa anumang cell sa katawan. Iyon lang marahil ang masasabi tungkol sa istruktura ng ATP.