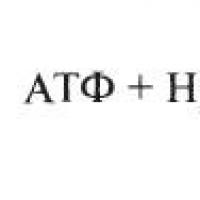Обмен и функции углеводов. Анаэробное и аэробное окисление глюкозы. Анаэробное окисление углеводов Технологические особенности проведения реакции окисления глюкозы
Анаэробное окисление углеводов происходит в клетках, органах и тканях без участия кислорода. Если процесс начинается с превращения глюкозы и заканчивается образованием молочной кислоты, то он называется анаэробным гликолизом, если начинается с превращения гликогена – гликогенолизом.
Гликолиз
На 1-й стадии гликолиза (рис. 7) происходит фосфорилирование глюкозы под действием фермента гексокиназы в присутствии АТФ и ионов Mg 2+ с образованием глюкозо-6-фосфата (Г-6-Ф), который на 2-й стадии превращается во фруктозо-6-фосфат (Ф-6-Ф). Эту реакцию катализирует глюкозофосфатизомераза . В ходе 3-й стадии осуществляется фосфорилирование фруктозо-6-фосфата, в результате чего образуется фруктозо-1,6-дифосфат (Ф-1,6-Ф). Катализ данной реакции обеспечивает в присутствии ионов Mg 2+ и АТФ фосфофруктокиназа .
На 4-й стадии Ф-1,6-Ф под действием альдолазы расщепляется на 2 фосфотриозы - глицеральдегид-3-фосфат (ГА-3-Ф) и дигидроксиацетонфосфат (ДАФ).
ДАФ под действием триозофосфатизомеразы превращается в ГА-3-Ф. Таким образом, из одной молекулы глюкозы мы получаем 2 молекулы ГА-3-Ф.
ГА-3-Ф в присутствии глицеральдегидфосфатдегидрогеназы , кофермента НАД + и фосфорной кислоты окисляется с образованием 1,3-дифосфоглицерата (1,3-ДФГ) и НАДН(Н +). Энергия, высвобождающаяся в этой реакции аккумулируется в макроэргической связи 1,3-ДФГ, который далее превращается в 3-фосфоглицерат под действием фосфоглицераткиназы . Этот процесс сопряжен с фосфорилированием АДФ (образуется АТФ на уровне субстратного фосфорилирования ).
3-ФГ при участии фосфоглицератмутазы превращается в 2-фосфоглицерат (2-ФГ), который подвергается дегидратации в присутствии енолазы и ионов Mg 2+ . В результате образуется фосфоенолпируват (ФЕП), содержащий макроэргическую связь. ФЕП под действием пируваткиназы при наличии ионов Mg 2+ превращается в енольную форму пирувата . Этот процесс сопряжен с синтезом АТФ на уровне субстратного фосфорилирования. Енольная форма пирувата таутомеризуется в кетонную форму.
На заключительной стадии гликолиза происходит восстановление пирувата до L -лактата в присутствии НАДН(Н +) и фермента лактатдегидрогеназы . Образующийся в этой реакции кофермент НАД + в дальнейшем используется в реакции окисления ГА-3-Ф.
Таким образом конечными продуктами гликолиза являются 2 молекулы L-лактата и 2 молекулы АТФ в расчете на 1 молекулу глюкозы.
Регуляция гликолиза осуществляется на уровне фосфофруктокиназной реакции. Фермент ингибируется высоким содержанием АТФ и цитрата. Аллостерическими активаторами фосфофруктокиназы являются АМФ, АДФ, Ф-6-Ф.
Гликогенолиз
Гликоген представляет собой разветвленный полисахарид, состоящий из остатков -D-глюкозы, связанных между собой в линейных участках молекулы -1,4-гликозидными связями, а в точках ветвления -1,6-гликозидными связями.
Под действием гликогенфосфорилазы происходит отщепление одного глюкозного остатка в линейных участках с переносом его на молекулу фосфорной кислоты, в результате чего образуется глюкозо-1-фосфат (Г-1-Ф). Гликогенфосфорилаза работает до тех пор, пока до ближайшей точки ветвления не останется 4 остатка глюкозы, затем в работу включается фермент олигосахаридтрансфераза , переносящий фрагмент из 3-х глюкозных остатков на соседнюю ветвь, таким образом, в точке ветвления остается один глюкозный остаток. Он отщепляется гидролитическим путем с помощью -1,6-гликозидазы в виде свободной молекулы глюкозы и вновь создаются условия для работы гликогенфосфорилазы в линейной цепи.
Продукт глигогенфосфорилазной реакции Г-1-Ф далее под действием фосфоглюкомутазы превращается в Г-6-Ф , который вовлекается в гликолитический путь (рис. 8).

Рис. 8. Схема распада гликогена в мышцах и печени.
Конечные продукты гликогенолиза – 2 молекулы L-лактата и 3 молекулы АТФ в расчете на 1 молекулу глюкозы.
100 р бонус за первый заказ
Выберите тип работы Дипломная работа Курсовая работа Реферат Магистерская диссертация Отчёт по практике Статья Доклад Рецензия Контрольная работа Монография Решение задач Бизнес-план Ответы на вопросы Творческая работа Эссе Чертёж Сочинения Перевод Презентации Набор текста Другое Повышение уникальности текста Кандидатская диссертация Лабораторная работа Помощь on-line
Узнать цену
Дыхание у микробов рассматривается как энергетический процесс или как совокупность различных химических реакций, окисления и расщепления веществ органического и неорганического происхождения. В результате этих химических реакций освобождается энергия, которая используется микробами для усвоения питательных веществ, синтеза бел кон их тела, движения, роста, размножения и других отправлений живого организма.
Примером высвобождения энергии может служить окисление глюкозы, которое можно выразить следующим у соединением:
С6 Н2О + 6 О 2 6 Н2О+ 6СО2 + 674 ккал.
Как видно из уравнений, в результате полного окисления одной молекулы глюкозы до конечных продуктов (воды и углекислого газа) выделяются 674 большие калории тепла.
Энергетический процесс дыхания у микробов протекает значительно сложнее и зависит от характера используемого питательного материала.
По типу дыхания микробов делят на аэробов и анаэробов, имеются микробы и с переходным типом дыхания.
Аэробы могут жить и развиваться при свободном доступе кислорода воздуха. Необходимую энергию для жизненных процессов они получают путем, поглощения кислорода и окисления питательных материалов.
Анаэробы способны развиваться без доступа кислорода. Свободный кислород воздуха на этих микробов оказывает вредное, губительное влияние. Строгие (облигатные) анаэробы (столбнячная палочка, возбудитель маслянокислого брожения) совсем не переносят кислорода. Необходимую энергию они получают путем расщепления органических веществ углеводов, белков, жиров, органических кислот, спиртов.
Факультативные анаэробы используют те же вещества, но применительно к условиям своего существования могут изменять анаэробный тип своего дыхания на аэробный. Так, дрожжи верхового брожения "при ограниченном притоке воздуха разлагают сахар на спирт и углекислоту; при обильной аэрации у них возникает аэробное дыхание с полным окислением сахара до углекислоты и воды. Молочнокислые бактерии в анаэробных условиях превращают глюкозу в молочную кислоту, при этом энергии освобождается несколько меньше, чем у аэробов. При недостатке кислорода денитрифицирующие бактерии пользуются для окисления органических соединений кислородом нитратов.
Приведенные примеры показывают разнообразие источников энергии и способов ее получения разными видами микробов; в силу этих причин у бактерий нет и быть но может единого механизма дыхания.
Большинство аэробных микроорганизмов окисляет органические питательные вещества в процессе дыхания до С02 и воды. Поскольку в молекуле СО 2 достигается высшая степень окисления углерода, в этом случае говорят о полном окислении и отличают этот тип дыхания от не полных окислений, при которых в качестве продуктов обмена выделяются частично окисленные органические соединения.
Под «полным окислением» имеется в виду лишь то, что не происходит вы деления каких-либо органических веществ; но это вовсе не означает, что окисляется весь поглощенный субстрат. В каждом случае значительная часть субстрата (40-70%) ассимилируется, т.е. превращается в вещества клеток.
Конечными продуктами «неполных окислений» могут быть уксусная, глюконовая, фумаровая, лимонная, молочная кислоты и ряд других соединений. Поскольку эти продукты сходны с теми, которые образуются при брожениях (пропионовая, масляная, янтарная, молочная кислоты и др.), а также в связи с тем, что при промышленных процессах брожения необходимы специальные технические устройства (ферментеры), неполные окисления называют также «окислительным брожением» или «аэробной ферментацией». Слова «брожение» и «ферментация» в этом случае отражают скорее технологический аспект.
Мы будем причислять к «неполным окислениям» также простое отщепление водорода от субстрата и использование микроорганизмов для катализа некоторых реакций, не имеющих для них какого-либо значения в обычных условиях. Ниже будут рассмотрены некоторые примеры таких окислений.
Дыхание бактерий
Дыхание является самой совершенной формой окислительного процесса и наиболее эффективным способом получения энергии. Главное преимущество дыхания состоит в том, что энергия окисляемого вещества - субстрата, на котором микроорганизм растет, используется наиболее полно. Поэтому в процессе дыхания перерабатывается гораздо меньше субстрата для получения определенного количества энергии, чем, например, при брожениях.
Рис.10. Роль пировнноградной кислоты в процессах дыхания и брожения.
Процесс дыхания заключается в том, что углеводы (или белки, жиры и другие запасные вещества клетки) разлагаются, окисляясь кислородом воздуха, до углекислого газа и воды. Выделяющаяся при этом энергия расходуется на поддержание жизнедеятельности организмов, рост и размножение. Бактерии вследствие ничтожно малых размеров своего тела не могут накапливать значительного количества запасных веществ. Поэтому они используют в основном питательные соединения среды.
В общем виде дыхание можно представить следующим уравнением:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2,87-106 дж.
глюкоза кислород углекислый газ вода энергия
За этой простой формулой скрывается сложная цепь химических реакций, каждая из которых катализируется специфическим ферментом.

Рис. 11. Схема гликолитического пути расщепления углеводов.
Ферментативные реакции, происходящие в процессе дыхания, в настоящее время хорошо изучены. Схема реакций оказалась универсальной, т. е. в принципе одинаковой у животных, растений и многих микроорганизмов, в том числе бактерий. Процесс дыхания при окислении глюкозы складывается из следующих основных этапов (рис. 10).
Сначала происходит образование фосфорных эфиров глюкозы - моноaктивированная глюкоза в форме дифосфата далее расщепляется на два триозофосфата (трехуглеродные соединения): фосфоглицериновый альдегид и диоксиацетон-фосфат, которые могут обратимо превращаться друг в друга.

Рис. 12. Цикл трикарбоновых кислот. Стрелками показано направление, а номерами - порядок реакций.
Далее в обмен вступает фосфоглицериновый альдегид, он окисляется в дифосфо-глицериновую кислоту. Назначение этого процесса заключается в отщеплении атомов водорода от окисляемого субстрата и переносе водорода с помощью специфических окислительных ферментов к кислороду воздуха (см. рис. 10, 11).
Водород от фосфоглицеринового альдегида присоединяется к ферменту - никотин-амиддинуклеотиду (НАД); при этом альдегид окисляется до кислоты и выделяется энергия. Часть этой энергии тратится на образование АТФ; при этом присоединяется фосфорная кислота к аденозиндифосфат у- АДФ. При гидролизе АТФ энергия освобождается и может быть затрачена на различные процессы синтеза белка и другие нужды клетки.
Фосфоглицериыовая кислота окисляется до пировиноградной кислоты. При этом также образуется АТФ, т. е. запасается энергия.
На этом завершается первая - анаэробная - стадия процесса дыхания, которая носит название гликолитического пути или пути Эмбдена - Мейергофа - Парнаса. Для осуществления этих реакций кислород не требуется. Образовавшаяся пировиноградная кислота (СН3СОСООН) является интереснейшим и очень важным соединением. Пути расщепления глюкозы в процессе дыхания и многих брожений, вплоть до образования пировиноградной кислоты, идут совершенно одинаково, что впервые было установлено русским биохимиком С. П. Костычевым. Пировиноградная кислота является тем центральным пунктом, от которого расходятся пути дыхания и брожений, откуда начинается специфическая для данного процесса цепь ферментативных превращений -специфическая цепь химических реакций (рис. 11).
В процессе дыхания пировиноградная кислота вступает в цикл трикарбоновых кислот (рис. 12). Это сложный замкнутый круг превращений, в результате которых образуются органические кислоты с 4, 5 и 6 атомами углерода (яблочная, молочная, фумаро-вая, а-кетоглутаровая и лимонная) и отщепля-«тся углекислота.
Прежде всего от пировиноградной кислоты, содержащей три атома углерода, отщепляется СО2 - образуется уксусная кислота, которая с коферментом А образует активное соединение - ацетилкоэнзим А. Он передает остаток уксусной кислоты (ацетил) на щавелевоуксусную кислоту {4 атома углерода), и образуется лимонная кислота (6 атомов углерода). Лимонная кислота претерпевает несколько превращений, в результате выделяется С02 и образуется пяти-углеродное соединение - а-кетоглутаровая кислота. От нее тоже отщепляется С02 (третья молекула углекислого газа), и образуется янтарная кислота (4 атома углерода), которая затем превращается в фумаровую, яблочную и, наконец, щавелевоуксусную кислоту.
На этом цикл замыкается. Щавелевоуксусная кислота снова может вступить в цикл.
Таким образом, в цикл вступает трехуглерод-ная пировиноградная кислота, и по ходу превращений выделяются 3 молекулы С02.
Водород пировиноградной кислоты, освобождающийся при дегидрировании в аэробных условиях, не остается свободным - он поступает в дыхательную цепь (так же, как водород глицеринового альдегида, отнятый при превращении его в глицериновую кислоту). Это - цепь окислительных ферментов.
Ферменты, которые первыми берут на себя водород от окисляемого субстрата, называются первичными дегидрогеназами.
В их состав входят ди- или трипиридин-нуклео-тиды: НАД или НАДФ и специфический белок. Механизм присоединения водорода - один и тот же:
Окисляемое вещество - Н2 + НАД -> окисленное вещество + НАД*Н2
Водород, полученный дегидрогеназой, затем присоединяется к следующей ферментной системе - флавиновым ферментам (ФМН или ФАД).
От флавиновых ферментов электроны попадают на цитохромы - железосодержащие протеиды (сложные белки). По цепи цитохро-мов передается не атом водорода, а только электроны. При этом происходит изменение валентности железа: Fe++ - e->Fe++
Заключительная реакция дыхания - это присоединение протона и электрона к кислороду воздуха и образование воды. Но прежде происходит активирование молекулы кислорода под действием фермента цитохромоксидазы. Активирование сводится к тому, что кислород приобретает отрицательный заряд за счет присоединения электрона окисляемого вещества. К активированному кислороду присоединяется водород (протон), образуя воду.
Кроме упомянутой цепи переносчиков электронов и водорода, известны и другие. Процесс этот гораздо более сложен, чем изложенная схема.
Биологический смысл этих превращений заключается в окислении веществ и образовании энергии, В результате окисления молекулы сахара (глюкозы) в АТФ запасается 12,6-1053ж энергии, в самой молекуле сахара содержится 28,6-106 дж, следовательно, полезно используется 44% энергии. Это очень высокий коэффициент полезного действия, если сравнить его с к. п. д. современных машин.
В процессе дыхания образуется огромное количество энергии. Если вся она выделилась бы сразу, то клетка перестала бы существовать. Но этого не происходит, потому что энергия выделяется не вся сразу, а ступенчато, небольшими порциями. Выделение энергии небольшими дозами обусловлено тем, что дыхание представляет собой многоступенчатый процесс, на отдельных этапах которого образуются различные промежуточные продукты (с разной длиной углеродной цепочки) и выделяется энергия. Выделяющаяся энергия не расходуется в виде тепла, а запасается в универсальном макроэр-гическом соединении - АТФ. При расщеплении АТФ энергия может использоваться в любых процессах, необходимых для поддержания жизнедеятельности организма: на синтез различных органических веществ, механическую работу, поддержание осмотического давления протоплазмы и т. д.
Дыхание является процессом, дающим энергию, однако его биологическое значение этим не ограничивается. В результате химических реакций, сопровождающих дыхание, образуется большое количество промежуточных соединений. Из этих соединений, имеющих различное количество углеродных атомов, могут синтезироваться самые разнообразные вещества клетки: аминокислоты, жирные кислоты, жиры, белки, витамины.
Поэтому обмен углеводов определяет остальные обмены веществ (белков, жиров). В этом его огромное значение.
С процессом дыхания, его химическими реакциями связано одно из удивительных свойств микробов - способность испускать видимый свет - люминесцировать.
Известно, что ряд живых организмов, в том числе бактерии, могут испускать видимый свет. Люминесценция, вызываемая микроорганизмами, известна уже в течение столетий. Скопление люминесцирующих бактерий, находящихся в симбиозе с мелкими морскими животными, иногда приводит к свечению моря; с люминесценцией встречались также при росте некоторых бактерий на мясе и т. д.
К основным компонентам, взаимодействие между которыми приводит к испусканию света, относятся восстановленные формы ФМН или НАД, молекулярный кислород, фермент люцифераза и окисляемое соединение - люциферин. Предполагается, что восстановленные НАД или ФМН реагируют с люциферазой, кислородом и люциферином, в результате чего электроны в некоторых молекулах переходят в возбужденное состояние и возвращение этих электронов на основной уровень сопровождается испусканием света. Люминесценцию у микробов рассматривают как «расточительный процесс», так как при этом энергетическая эффективность дыхания снижается.
Большинство организмов в биосфере находятся в аэробных условиях. В присутствии кислорода в организме происходит полное «сжигание» углеводов и других молекул «клеточного топлива» до конечных продуктов - С0 2 и Н 2 0.
Рис. 19.1. Схема полного окисления глюкозы до шести молекул С0 2 и энергетическая эффективность этого процесса (баланс АТФ); пути образования АТФ:
СФ - субстратное фосфорилирование; ОФ - окислительное фосфорилирование
Суммарный процесс полного окисления глюкозы в аэробных условиях описывается стехиометрическим уравнением
В этом сложном, многостадийном процессе окисления глюкозы можно выделить три этапа (рис. 19.1).
- На первом этапе протекают реакции аэробного гликолиза, в процессе которых глюкоза расщепляется на две молекулы пирувата. Этот этап составляет начальную фазу разложения углеводов, его называют «подготовительным».
- На втором этапе протекает цепь реакций окислительного декарбоксили- рования пирувата, в результате которых происходит образование одного из центральных метаболитов клетки ацетил-S-KoA и окисление одного атома углерода пирувата до С0 2 . Поскольку в расчете на одну молекулу глюкозы образуется две молекулы пирувата, на этом этапе уже происходит окисление двух атомов углерода глюкозы до С0 2 .
- Третий этап представляет собой крайне важный набор реакций полного окисления ацетильного остатка, который получил название цикла трикарбо- новых кислот (ЦТК).
Процесс аэробного окисления углеводов сопровождается освобождением большого количества энергии (2880 кДж/моль глюкозы). Если суммировать общий выход АТФ в этом процессе, то он составит 38 молекул (см. рис. 19.1). Как отмечалось ранее (гл. 15), на синтез одной макроэргической связи АТФ
необходимо 31 кДж, а на синтез 38 молекул АТФ расходуется 1178 кДж, т. е. более 40% свободной энергии полного окисления глюкозы запасается в молекулах АТФ. Это свидетельствует о высокой эффективности окислительных процессов, протекающих в аэробных условиях по сравнению с анаэробными. В процессе аэробного окисления метаболически доступная энергия кумулируется в молекулах восстановленных НАДН и ФАДН 2 , которые затем окисляются в ходе кислородзависимого процесса окислительного фосфорилирования , результатом которого является образование 34 молекул АТФ, и только 4 молекулы АТФ образуются путем субстратного фосфорилирования: 2АТФ в гликолизе (I этап) и 2АТФ - в ЦТК (2 оборота, III этап).
Следует отметить, что, если первый этап аэробного окисления углеводов - гликолиз является специфическим процессом катаболизма глюкозы, то два последующие - окислительное декарбоксилирование пирувата и ЦТК относятся к общим путям катаболизма (ОПК). После образования пирувата (С 3 -фрагмент) и ацетил-КоА (С 2 -фрагмент), образующихся при распаде не только глюкозы, но и липидов и аминокислот, пути окисления этих веществ до конечных продуктов происходят одинаково по механизму реакций ОПК.
Аэробное окисление углеводов - основной путь образования энергии для организма. Непрямой - дихотомический и прямой - апотомический.
Прямой путь распада глюкозы - пентозный цикл - приводит к образованию пентоз и накоплению НАДФН 2 . Пентозный цикл характеризуется последовательным отщеплением от молекул глюкозы каждого из ее 6 атомов углерода с образованием в течение одного цикла по 1 молекуле углекислого газа и воды. Распад всей молекулы глюкозы происходит в течение 6 повторяющихся циклов.
Значение пентозофосфатного цикла окисления углеводов в обмене веществ велико:
1. Он поставляет восстановленный НАДФ, необходимый для биосинтеза жирных кислот, холестерина и т.д. За счет пентозного цикла на 50% покрывается потребность организма в НАДФН 2 .
2. Поставка пентозофосфатов для синтеза нуклеиновых кислот и многих коферментов.
Реакции пентозного цикла протекают в цитоплазме клетки.
При ряде патологических состояний удельный вес пентозного пути окисления глюкозы возрастает.
Непрямой путь - распад глюкозы до углекислого газа и воды с образованием 36 молекул АТФ.
1. Распад глюкозы или гликогена до пировиноградной кислоты
2. Превращение пировиноградной кислоты в ацетил- КоА
Окисление ацетил-КоА в цикле Кребса до углекислого газа и воды
С 6 Н 12 О 6 + 6 О 2 6 СО 2 + 6 Н 2 О + 686 ккал
В случае аэробного превращения пировиноградная кислота подвергается окислительному декарбоксилированию с образованием ацетил- КоА, который затем окисляется до углекислого газа и воды.
Окисление пирувата до ацетил-КоА, катализируется пируватдегидрогеназной системой и протекает в несколько стадий. Суммарно реакция:
Пируват + НАДН + НS-КоА ацетил- КоА+ НАДН 2 + СО 2 реакция практически необратима
Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот или цикле Кребса. Этот процесс протекает в митохондриях.
Цикл состоит из 8 последовательных реакций:
В этом цикле, молекула, содержащая 2 атома углерода (уксусная кислота в форме ацетил-КоА) реагирует с молекулой щавелевоуксусной кислоты, в результате чего образуется соединение с 6 атомами углерода - лимонная кислота. В процессе дегидрирования, декарбоксилирования и подготовительной реакции лимонная кислота вновь превращается в щавелевоуксусную кислоту, которая легко соединяется с другой молекулой ацетил- КоА.
1) ацетил-КоА + оксалоацетат (ЩУК) лимонная кислота
цитратсинтаза
2) лимонная кислота изолимонная кислота
аконитатгидратаза
3)изолимонная к-та+НАД-кетоглутаровая к-та+НАДН 2 + СО 2
изоцитратдегидрогеназа
4)-кетоглутаровая к-та+НS-КоА+НАДсукцинилSКоА+НАДН 2 + СО 2
5) сукцинил-КоА+ГДФ+Фнянтарная кислота+ГТФ+НS-КоА
сукцинил КоА синтетаза
6) янтарная кислота+ФАДфумаровая кислота+ФАДН 2
сукцинатдегидрогеназа
7) фумаровая кислота+ Н 2 О L яблочная кислота
фумаратгидратаза
8) малат+ НАДоксалоацетат+ НАДН 2
малатдегидрогеназа
Итого при расщеплении в тканях молекулы глюкозы синтезируется 36 молекул АТФ. Несомненно, это в энергетическом отношении более эффективный процесс чем гликолиз.
Цикл Кребса - общий конечный путь, которым завершается обмен углеводов, жирных кислот и аминокислот. Все эти вещества включаются в цикл Кребса на том или другом этапе. Далее происходит биологическое окисление или тканевое дыхание, главной особенностью которого является то, что оно протекает постепенно, через многочисленные ферментативные стадии. Этот процесс происходит в митохондриях, клеточных органеллах, в которых сосредоточено большое количество ферментов. В процессе участвуют пиридинзависимые дегидрогеназы, флавинзависимые дегидрогеназы, цитохромы, коэнзим Q - убихинон, белки, содержащие негеминовое железо.
Интенсивность дыхания управляется соотношением АТФ/АДФ. Чем меньше это отношение, тем интенсивнее идет дыхание, обеспечивая выработку АТФ.
Также цикл лимонной кислоты является в клетке главным источником двуокиси углерода для реакций карбоксилирования, с которых начинается синтез жирных кислот и глюконеогенез. Та же двуокись углерода поставляет углерод для мочевины и некоторых звеньев пуриновых и пиримидиновых колец.
Взаимосвязь между процессами углеводного и азотистого обмена также достигаются посредством промежуточных продуктов цикла лимонной кислоты.
Существует несколько путей, по которым промежуточные продукты цикла лимонной кислоты включаются в процесс липогенеза. Расщепление цитрата приводит к образованию ацетил-КоА, играющего роль предшественника в биосинтезе жирных кислот.
Изоцитрат и малат обеспечивают образование НАДФ, который расходуется в последующих восстановительных этапах синтеза жиров.
Роль ключевого фактора, определяющего превращение НАДН играет состояние адениннуклеотидов. Высокое содержание АДФ и низкое АТФ свидетельствует о малом запасе энергии. При этом НАДН вовлекается в реакции дыхательной цепи, усиливая сопряженные с запасанием энергии процессы окислительного фосфорилирования. Обратное явление наблюдается при низком содержании АДФ и высоком АТФ. Ограничивая работу системы переноса электронов, они способствуют использованию НАДН в других восстановительных реакциях, таких как синтез глутамата и глюконеогенез.
Молекулярный кислород атмосферы, очевидно, имеет биогенное происхождение. Появление его связано с процессом фотосинтеза древнейших цианобактерий или их предков, впервые ставших использовать в процессе фотосинтеза донором водорода воду. Переход прокариот к анаэробному окислению оказался возможным только на определенном этапе
эволюции, когда в клетке была сформирована более или менее полная дыхательная цепь.
Большая часть аэробных прокариотных организмов потребляет в качестве источника энергии различные органические соединения, окисляя их до конечных продуктов СО 2 и Н 2 О. Аэробное окисление органических веществ в прокариотной клетке проходит подобно аэробному дыханию эукариот. В основе его лежит окисление пировиноградной кислоты по циклу трикарбоновых кислот (ЦТК – цикл Кребса).
Включению пировиноградной кислоты в цикл Кребса предшествует сложная реакция окисления ее до ацетил-К 0 А, катализируемая пируватдегидрогеназным комплексом:
СН 3 – СО – СООН + К 0 А – SН + НАД + →СН 3 – СО ~ К 0 А + НАД · Н 2 + СО 2
Собственно цикл Кребса начинается с реакции конденсации ацетил-К 0 А с молекулярной щавелевоуксусной кислоты при участии фермента цитрасинтазы.
Цикл Кребса выполняет две важные функции для клетки. В реакциях этого цикла осуществляется полное окисление органического субстрата с отщеплением водорода и переходом его на фермент, помимо этого идет снабжение клетки веществами-предшественниками.
Суммарно цикл Кребса можно выразить следующим уравнением:
СН 3 СОСООН + 2Н 2 О→ 3СО 2 + 8Н
Исходным субстратом для цикла трикарбоновых кислот служат не только углеводы, но и жирные кислоты и многие аминокислоты.
Цикл Кребса сопряжен с дыхательной цепью. Основная функция дыхательной цепи заключается в запасании энергии клеткой, освобождающейся в процессе переноса электронов путем трансформации ее в химическую энергию фосфатных связей в молекулах АТФ.
В состав дыхательной цепи аэробных прокариотных организмов входят: НАД-дегидрогеназы, ФАД-или ФМН-дегидрогеназы, убихинон и система цитохромов. НАД-дегидрогеназы катализируют отщепление водорода от окисляемого субстрата и передачу его на стартовые переносчики дыхательной цепи – НАД Н 2 -дегидрогеназы. С них водород передается в дыхательную цепь на ФАД-или ФМН-дегидрогеназы, затем на убихинон и далее на систему цитохромов. При передаче водорода по дыхательной цепи происходит расщепление его атомов на протоны и электроны. Протоны выделяются в среду, а электроны передаются далее по дыхательной цепи на терминальный переносчик – цитохромоксидазу. Последняя передает их на конечный акцептор – молекулярный кислород, который активизируется и соединяется с водородом.
Перенос электронов по дыхательной цепи на все более низкие энергетические уровни приводит к освобождению значительного количества свободной энергии, которая аккумулируется клеткой в фосфатных связях в
форме молекул АТФ. Поскольку реакции фосфорилирования идут сопряженно с реакциями окисления, этот процесс получил название окислительного фосфорилирования . В основе его лежит разность окислительно-восстановительного потенциала донора и акцептора электронов. Образование АТФ обычно происходит на участках дыхательной цепи с большой разностью потенциала. На примере митохондрий эукариот определены три участка в цепи дыхания. Первый участок связан с переносом водорода НАД Н 2 -дегидрогеназой на ФАД-или ФМН-дегидрогеназу. Второй участок сопряжен с активностью убихинона, осуществляющего перенос электронов от ФАД-или ФМН-дегидрогеназы на систетему цитохромов, Третий, последний участок связан с переносом электронов цитохромоксидазой на молекулярный кислород.
Об эффективности реакций окислительного фосфорилирования судят по отношению р/о (количество потребленных молекул неорганического фосфора на один атом поглощенного кислорода).У эокариот отношение р/о равно 3. У многих прокариот отношение р/о может быть меньше трех, что объясняется выпадением некоторых участков дыхательной цепи.
Среди прокариот-аэробов имеются микроорганизмы, способные получать энергию за счет неполного аэробного окисления некоторых органических веществ.
Уксуснокислые бактерии представлены палочками небольших размеров, в молодой культуре они подвижны. Все виды – облигатные аэробы, довольно требовательные к субстратам, особенно к витаминам и в первую очередь к пантотеновой кислоте. Наиболее характерна способность бактерий этой группы окислять этиловый спирт с образованием уксусной кислоты при участии НАД-зависимых дегидрогеназ.
Подчеркнем, что процессы неполного анаэробного окисления не имеют ничего общего с процессами брожения. Энергия для жизнедеятельности клетки образуется в реакциях окисленого фосфорирования, но в меньшем количестве, чем при полном аэробном окислении, так как часть ее сохраняется в недоокисленных конечных продуктах.
Различные виды уксуснокислых бактерий в качестве окисляемого субстрата способны использовать как одноатомные спирты, так и многоатомные спирты – производные сахаров. При окислении одноатомных спиртов образуются различные кислоты. Окисление многоатомных спиртов приводит к образованию альтоз и кетоз. В промышленности уксуснокислые бактерии применяются для получения столового уксуса, а также аскорбиновой кислоты.
В отличие от эукариот, осуществляющих дыхание толькл за счет окисления органических веществ, среди прокариот имеются группы хемолитотрофных микроорганизмов, способных в процессе катаболизма окислять неорганические вещества субстрата.
Дыхательная цепь хемолитотрофных микроорганизмов включает в основном те же ферменты-переносчики электронов, что и дыхательная цепь хемоорганотрофов. Специфика функционирования дыхательной цепи этой группы микроорганизмов заключается в том, что при окислении неорганических соединений, обладающих разным окислительно-восстановительным потенциалом, электроны с окисляемого субстрата включаются в дыхательную цепь на разных энергетических уровнях. Поэтому для обеспечения клетки энергией микроорганизмы вынуждены окислять огромное количество субстрата.
К хемолитотрофным микроорганизмам, получающим энергию за счет окисления неорганических веществ, относятся нитрифицирующие бактерии, железобактерии, тионовые бактерии и др.
С точки зрения лабильности метобализма особый интерес представляют карбоксибактерии. Эти микроорганизмы могут вести себя как автотрофы, потребляя в качестве единственного источника углерода и энергии угарный газ (СО), и как гетеротрофы, используя источником углерода и энергии органические вещества – спирты и органические кислоты.
Суммарно процессы катаболизма и анаболизма карбоксидобактерий-автотрофов можно представить следующим уравнением:
24СО + 11О 2 + Н 2 О→ 23СО 2 + (СН 2 О),
где (СН 2 О) – символ биомассы.
Из уравнения следует, что окисление СО – неэффективный способ получения энергии, поэтому микроорганизмы вынуждены окислять большое количество субстрата.
Выделение СО современным транспортом и промышленными предприятиями загрязняет атмосферу этим соединением. Единственный путь удаления СО из окружающей среды – утилизация его в обмене микробной клетки.
Анаэробное окисление: нитратное и сульфатное дыхание
Анаэробное окисление встречается только среди представителей царства прокариот. Оно присуще микроорганизмам, способным переходить от аэробного образа жизни к анаэробному, используя в качестве конечного акцептора электронов как молекулярный кислород, так и азот нитратов и серу сульфатов.
Типичным примером таких микроорганизмов являются денитрифицирующие бактерии.
Дыхательная цепь денитрифицирующих бактерий включает все основные ферменты-переносчики электронов, характерные для дыхательной цепи аэробов. Только конечное звено цитохромной системы – цитохромоксидаза замещена у них на нитратредуктазу, катализирующую перенос электронов на азот нитратов. Нитратредуктазы относятся к индуцибельным ферментам, синтезируемым клеткой только в анаэробных условиях при наличии нитратов в среде.
Процесс денитрификации состоит из 4 восстановительных стадий, каждая из которых катализируется соответствующей нитратредуктазой. На первой стадии происходит восстановление нитратов в нитриты:
азот+5 принимая 2 протона и 2 электрона восстанавливается в азот нитритов NО 2 - +3:
NО 3 - + 2e - + 2Н + →NО 2 - + Н 2 О.
NО 2 - + e - + Н + → NО + ОН -
2NО + 2e - + 2Н + → N 2 О + Н 2 О
N 2 О + 2e - + 2Н + →N 2 + Н 2 О
Использование азота в качестве акцептора электронов позволяет денитрифицирующим бактериям полностью окислять органические вещества субстрата до конечных продуктов СО 2 и Н 2 О. Поэтому энергетический выход нитратного дыхания практически приближается к обычному аэробному окислению.
Поскольку денитрофицирующие бактерии переключаются на нитратное дыхание, только попадая в анаэробные условия, приспособление их к анаэробному образу жизни следует считать эволюционно вторичным и рассматривать как возврат к анаэробиозу от типичного аэробного окисления.
К анаэробному окислению способны и сульфатвосстанавливающие бактерии, относящиеся к родам Desulfotomaculum, Desulfonema, Desulfovibrio и др. Пути получения энергии у сульфатвосстанавливающих бактерий могут быть разными. Это процесс брожения органических веществ, сопровождающиеся образованием АТФ в результате субстратного фосфорилирования, сульфатное дыхание, предусматривающее окисление органических веществ в анаэробных условиях с переносом электронов на серу сульфатов. Бактерии этой гетерогенной группы способны получать энергию также за счет окисления молекулярного водорода, сопряженного с востановлением сульфатов.
Способность сульфатвосстанавливающих бактерий использовать молекулярный водород для получения энергии позволяет отнести их к анаэробным хемолитотрофным микроорганизмам.
В процессе окисления молекулярного водорода получают энергию и метанообразующие бактерии, использующие в качестве акцептора электронов углекислый газ. Для бактерий этой группы СО 2 выступает одновременно источником углерода и акцептором электронов:
4Н 2 + СО 2 →СН 4 + 2Н 2 О
Изучение различных типов катаболизма прокариот дает возможность предположить, что именно совершенствование способов получения энергии клеткой лежит в основе эволюции представителей этого царства.
Наиболее древней группой прокариот являются анаэробные бактерии, добывающие энергию в процессах брожения за счет субстратного фосфорилирования.
Существенным этапом на пути эволюции прокариот следует считать появление фототрофных бактерий, использующих в качестве основного источника энергии солнечный свет и в качестве основного источника углерода СО 2 .
Развитие фотосинтетиков-аэробов, в первую очередь цианобактерий, привело к обогащению среды молекулярным кислородом. В клетке аэробных бактерий сложилась еще одна система электроного транспорта и сопряженный с ней механизм фосфорилирования – окислительное фосфорилирование.
В настоящее время в царстве прокариот мы встречаемся с поразительным разнообразием типов катаболизма. Однако доминирующим и эволюционно господствующим типом катаболизма, несомненно, являтся аэробное окисление со всем его многообразием доноров и акцепторов.