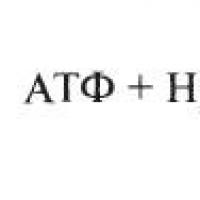Karbonhidratların metabolizması ve fonksiyonları. Glikozun anaerobik ve aerobik oksidasyonu. Karbonhidratların anaerobik oksidasyonu Glikoz oksidasyon reaksiyonunun teknolojik özellikleri
Karbonhidratların anaerobik oksidasyonu, oksijenin katılımı olmadan hücrelerde, organlarda ve dokularda meydana gelir. Süreç glikozun dönüşümüyle başlıyorsa ve laktik asit oluşumuyla bitiyorsa, glikojen - glikojenolizin dönüşümüyle başlıyorsa buna anaerobik glikoliz denir.
Glikoliz
Glikolizin 1. aşamasında (Şekil 7), enzimin etkisi altında glikozun fosforilasyonu meydana gelir heksokinaz oluşumu ile ATP ve Mg 2+ iyonlarının varlığında glikoz-6-fosfat(G-6-F), 2. aşamada dönüşüyor fruktoz 6-fosfat(F-6-F). Bu reaksiyon katalize edilir glukoz fosfat izomeraz . 3. aşamada fruktoz-6-fosfat fosforile edilir ve bunun sonucunda formasyon oluşur. fruktoz 1,6-bisfosfat(F-1,6-F). Bu reaksiyonun katalizi Mg2+ iyonlarının varlığında sağlanır ve ATPfosfofruktokinaz .
4. aşamada F-1,6-F'nin etkisi altında aldolazlar 2 fosfotrioza ayrılır gliseraldehit-3-fosfat(GA-3-F) ve dihidroksiaseton fosfat(DAF).
DAP etki altında triosefosfat izomeraz GA-3-F'ye dönüşür. Böylece bir glikoz molekülünden 2 molekül GA-3-P elde ederiz.
GA-3-F huzurunda gliseraldehit fosfat dehidrojenaz , koenzim NAD + ve fosforik asit oluşturmak üzere oksitlenir 1,3-difosfogliserat(1,3-DPG) ve NADH(H+). Bu reaksiyonda açığa çıkan enerji, 1,3-DPG'nin yüksek enerjili bağında birikir ve bu bağ daha sonra dönüştürülür. 3-fosfogliserat Etkisi altında fosfogliserat kinaz . Bu süreç ADP'nin fosforilasyonuyla ilişkilidir (oluşum ATP düzeyinde substrat fosforilasyonu).
3-MY katılımıyla fosfogliserat mutaz dönüşür 2-fosfogliserat(2-PG), varlığında dehidrasyona uğrar enolazlar ve Mg2+ iyonları. Sonuç olarak, fosfoenolpiruvat(PEP), yüksek enerjili bir bağ içerir. PEP, Mg2+ iyonlarının varlığında piruvat kinazın etkisi altında enol formuna dönüştürülür piruvat. Bu işlem, substrat fosforilasyonu seviyesinde ATP sentezi ile ilişkilidir. Piruvatın enol formu, keton formuna totomerleşir.
Glikolizin son aşamasında piruvat indirgenir. L-laktat NADH(H+) ve enzim varlığında laktat dehidrogenaz . Bu reaksiyonda oluşan koenzim NAD+ daha sonra GA-3-P'nin oksidasyon reaksiyonunda kullanılır.
Böylece, glikolizin son ürünleri, 1 molekül glikoz başına 2 molekül L-laktat ve 2 molekül ATP'dir.
Düzenleme seviyede glikoliz meydana gelir fosfofruktokinaz reaksiyonlar. Enzim, yüksek düzeyde ATP ve sitrat tarafından inhibe edilir. Fosfofruktokinazın allosterik aktivatörleri AMP, ADP, P-6-P'dir.
Glikojenoliz
Glikojen, molekülün doğrusal bölgelerinde a-1,4-glikosidik bağlarla ve dallanma noktalarında a-1,6-glikosidik bağlarla birbirine bağlanan a-D-glikoz kalıntılarından oluşan dallanmış bir polisakkarittir.
Etkisi altında glikojen fosforilaz Bir glikoz kalıntısının bölünmesi, bir fosforik asit molekülüne aktarılmasıyla doğrusal kesitlerde meydana gelir ve bunun sonucunda formasyon oluşur. glikoz-1-fosfat(G-1-F). Glikojen fosforilaz, en yakın dallanma noktasından önce 4 glikoz kalıntısı kalana kadar çalışır, ardından enzim çalışmaya başlar. oligosakarit transferaz 3 glikoz kalıntısının bir fragmanının bitişik dala aktarılması, böylece dal noktasında bir glikoz kalıntısının bırakılması. Serbest glikoz molekülü formundaki -1,6-glikosidaz yardımıyla hidrolitik olarak parçalanır ve glikojen fosforilazın doğrusal bir zincirde çalışması için yeniden koşullar yaratılır.
Glikojen fosforilaz reaksiyonunun ürünü G-1-F daha da etkisi altında fosfoğlukomutaz dönüşür G-6-F, dahil olan glikolitik yol (Şek. 8).

Pirinç. 8. Kaslarda ve karaciğerde glikojenin parçalanma şeması.
Glikojenolizin son ürünleri, 1 molekül glikoz başına 2 molekül L-laktat ve 3 molekül ATP'dir.
100 rupi ilk siparişe bonus
İşin türünü seçin Diploma çalışması Ders çalışması Özet Yüksek lisans tezi Uygulama raporu Makale Raporu İnceleme Test çalışması Monografi Problem çözme İş planı Soru cevapları Yaratıcı çalışma Deneme Çizim Denemeler Çeviri Sunumlar Yazma Diğer Metnin benzersizliğini arttırma Yüksek lisans tezi Laboratuvar çalışması Çevrimiçi yardım
Fiyatı öğren
Mikroplarda solunum, bir enerji süreci veya çeşitli kimyasal reaksiyonlar, oksidasyon ve organik ve inorganik kökenli maddelerin parçalanmasının bir kümesi olarak kabul edilir. Bu kimyasal reaksiyonların bir sonucu olarak, mikroplar tarafından besinlerin emilimi, vücutlarındaki proteinlerin sentezi, hareket, büyüme, üreme ve canlı organizmanın diğer işlevleri için kullanılan enerji açığa çıkar.
Enerji salınımının bir örneği, aşağıdaki bileşikle ifade edilebilen glikozun oksidasyonudur:
C6 H2O + 6 O2 6 H2O+ 6CO2 + 674 kcal.
Denklemlerden görülebileceği gibi, bir glikoz molekülünün nihai ürünlere (su ve karbondioksit) tamamen oksidasyonu sonucunda 674 büyük kalorilik ısı açığa çıkar.
Mikroplardaki enerjik solunum süreci çok daha karmaşıktır ve kullanılan besin maddesinin doğasına bağlıdır.
Solunum türüne göre mikroplar aeroblara ve anaeroblara ayrılır; geçiş tipi solunuma sahip mikroplar vardır.
Aeroblar oksijene serbest erişimle yaşayabilir ve gelişebilirler. Yaşam süreçleri için gerekli enerjiyi oksijeni emerek ve besin maddelerini oksitleyerek elde ederler.
Anaeroblar oksijene erişim olmadan gelişebilirler. Havadaki serbest oksijenin bu mikroplar üzerinde zararlı, yıkıcı etkisi vardır. Katı (zorunlu) anaeroblar (tetanoz basili, bütirik asit fermantasyonunun etken maddesi) oksijeni hiç tolere etmez. Gerekli enerjiyi karbonhidrat, protein, yağ, organik asit ve alkol gibi organik maddeleri parçalayarak elde ederler.
Fakültatif anaeroblar aynı maddeleri kullanırlar, ancak varoluş koşullarına bağlı olarak solunumlarının anaerobik tipini aerobik olarak değiştirebilirler. Böylece, üst fermente eden maya "sınırlı bir hava kaynağıyla şekeri alkol ve karbondioksite ayrıştırır; bol havalandırmayla, şekerin karbondioksit ve suya tamamen oksidasyonuyla aerobik solunum yaşarlar. Laktik asit bakterileri, anaerobik koşullar altında glikozu laktik asit haline dönüştürür. asit ve bir miktar enerji aeroblara göre daha az salınır. Oksijen eksikliği olduğunda, denitrifikasyon bakterileri organik bileşikleri oksitlemek için nitrat oksijeni kullanır.
Verilen örnekler, enerji kaynaklarının çeşitliliğini ve onu farklı mikrop türleri tarafından elde etme yöntemlerini göstermektedir; Bu nedenlerden dolayı bakterilerin tek bir solunum mekanizması yoktur ve olabilir.
Çoğu aerobik mikroorganizma, solunum sırasında organik besinleri CO2 ve suya oksitler. Karbonun en yüksek oksidasyon derecesi CO2 molekülünde elde edildiğinden, bu durumda tam oksidasyondan söz ederler ve bu tür solunumu, kısmen oksitlenmiş organik bileşiklerin metabolik ürünler olarak salındığı eksik oksidasyonlardan ayırırlar.
"Tam oksidasyon" ile yalnızca herhangi bir organik maddenin serbest kalmadığını kastediyoruz; ancak bu, emilen substratın tamamının oksitlendiği anlamına gelmez. Her durumda, substratın önemli bir kısmı (%40-70) asimile edilir; hücre maddelerine dönüşür.
"Eksik oksidasyonların" son ürünleri asetik, glukonik, fumarik, sitrik, laktik asitler ve bir dizi başka bileşik olabilir. Bu ürünler fermantasyon sırasında oluşanlara (propiyonik, bütirik, süksinik, laktik asitler vb.) benzediğinden ve ayrıca endüstriyel fermantasyon işlemlerinin özel teknik cihazlar (fermentörler) gerektirmesi nedeniyle, tamamlanmamış oksidasyonlara "oksidatif fermantasyon" da denir. " veya "aerobik fermantasyon". Bu durumda "fermantasyon" ve "fermantasyon" kelimeleri daha çok teknolojik bir yönü yansıtmaktadır.
Ayrıca, hidrojenin substrattan basit bir şekilde çıkarılmasını ve normal koşullar altında kendileri için hiçbir önemi olmayan belirli reaksiyonları katalize etmek için mikroorganizmaların kullanılmasını da "eksik oksidasyonlar" olarak sınıflandıracağız. Bu tür oksidasyonların bazı örnekleri aşağıda tartışılacaktır.
Bakterilerin solunumu
Solunum, oksidatif sürecin en gelişmiş şeklidir ve enerji elde etmenin en etkili yoludur. Solunumun temel avantajı, mikroorganizmanın üzerinde büyüdüğü substrat olan oksitlenmiş maddenin enerjisinin en iyi şekilde kullanılmasıdır. Bu nedenle, solunum işlemi sırasında, belirli bir miktarda enerji elde etmek için, örneğin fermantasyon sırasında olduğundan çok daha az substrat işlenir.
Şekil 10. Pirüvik asidin solunum ve fermantasyon süreçlerindeki rolü.
Solunum süreci, karbonhidratların (veya proteinlerin, yağların ve hücrenin diğer rezerv maddelerinin) atmosferik oksijen tarafından oksitlenerek karbondioksit ve suya ayrışması gerçeğinden oluşur. Bu durumda açığa çıkan enerji, organizmaların hayati işlevlerinin sürdürülmesine, büyümesine ve üremesine harcanır. Bakteriler, vücutlarının ihmal edilebilecek kadar küçük olması nedeniyle önemli miktarda rezerv madde biriktiremezler. Bu nedenle esas olarak çevredeki besin bileşiklerini kullanırlar.
Genel olarak nefes alma aşağıdaki denklemle temsil edilebilir:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2,87-106 J.
glikoz oksijen karbondioksit su enerji
Bu basit formülün arkasında, her biri belirli bir enzim tarafından katalize edilen karmaşık bir kimyasal reaksiyonlar zinciri yatmaktadır.

Pirinç. 11. Karbonhidratların parçalanması için glikolitik yolun şeması.
Solunum sırasında meydana gelen enzimatik reaksiyonlar artık iyi bir şekilde incelenmiştir. Reaksiyon şemasının evrensel olduğu, yani prensip olarak hayvanlarda, bitkilerde ve bakteriler dahil birçok mikroorganizmada aynı olduğu ortaya çıktı. Glikoz oksidasyonu sırasındaki solunum süreci aşağıdaki ana aşamalardan oluşur (Şekil 10).
İlk olarak, glikozun fosfor esterleri oluşturulur - difosfat formundaki monoaktif glikoz ayrıca iki trioz fosfata (üç karbonlu bileşikler) bölünür: fosfogliseraldehit ve dihidroksiaseton fosfat, bunlar geri dönüşümlü olarak birbirine dönüştürülebilir.

Pirinç. 12. Trikarboksilik asit döngüsü. Oklar yönü, sayılar ise reaksiyonların sırasını gösterir.
Daha sonra fosfogliseraldehit değişime girer ve difosfogliserik asite oksitlenir. Bu işlemin amacı, oksitlenmiş substrattan hidrojen atomlarının çıkarılması ve hidrojenin spesifik oksidatif enzimlerin yardımıyla atmosferik oksijene aktarılmasıdır (bkz. Şekil 10, 11).
Fosfogliseraldehidden gelen hidrojen, nikotin amid dinükleotid (NAD) enzimine bağlanır; bu durumda aldehit bir aside oksitlenir ve enerji açığa çıkar. Bu enerjinin bir kısmı ATP oluşumuna harcanır; bu durumda adenozin difosfat y-ADP'ye fosforik asit eklenir. ATP hidrolize edildiğinde enerji açığa çıkar ve protein sentezinin çeşitli süreçlerine ve diğer hücre ihtiyaçlarına harcanabilir.
Fosfogliserik asit pirüvik asite oksitlenir. Aynı zamanda ATP de oluşur, yani enerji depolanır.
Bu, glikolitik yol veya Embden-Meyerhof-Parnas yolu olarak adlandırılan solunum sürecinin ilk anaerobik aşamasını tamamlar. Bu reaksiyonların gerçekleşmesi için oksijene gerek yoktur. Ortaya çıkan piruvik asit (CH3COCOOH) ilginç ve çok önemli bir bileşiktir. Solunum sırasında glikozun parçalanması ve pirüvik asit oluşumuna kadar birçok fermantasyon yolları, ilk olarak Rus biyokimyacı S.P. Kostychev tarafından belirlenen şekilde tamamen aynı şekilde ilerlemektedir. Piruvik asit, solunum ve fermantasyon yollarının ayrıldığı, belirli bir sürece özgü bir enzimatik dönüşüm zincirinin - belirli bir kimyasal reaksiyon zincirinin - başladığı merkezi noktadır (Şekil 11).
Solunum sırasında pirüvik asit, trikarboksilik asit döngüsüne girer (Şekil 12). Bu, 4, 5 ve 6 karbon atomlu organik asitlerin (malik, laktik, fumarik, a-ketoglutarik ve sitrik) oluştuğu ve karbondioksitin elimine edildiği karmaşık bir kapalı dönüşüm çemberidir.
Her şeyden önce, CO2, üç karbon atomu içeren pirüvik asitten ayrılır - koenzim A ile aktif bir bileşik oluşturan asetik asit oluşur - asetil koenzim A. Geri kalan asetik asidin (asetil) oksaloasetik asite (4 karbon) aktarılmasını sağlar. atom) ve sitrik asit oluşur (6 karbon atomu). Sitrik asit çeşitli dönüşümlere uğrar, bunun sonucunda CO2 açığa çıkar ve beş karbonlu bir bileşik oluşur - a-ketoglutarik asit. CO2 (üçüncü karbondioksit molekülü) de ondan ayrılır ve süksinik asit (4 karbon atomu) oluşur, bu daha sonra fumarik, malik ve son olarak oksaloasetik asite dönüşür.
Bu döngüyü tamamlar. Oksaloasetik asit tekrar döngüye girebilir.
Böylece döngüye üç karbonlu pirüvik asit girer ve dönüşüm sırasında 3 CO2 molekülü açığa çıkar.
Aerobik koşullar altında dehidrojenasyon sırasında açığa çıkan pirüvik asit hidrojeni serbest kalmaz - solunum zincirine girer (tıpkı gliseraldehitin hidrojeni gibi, gliserik aside dönüştürüldüğünde alınır). Bu bir oksidatif enzim zinciridir.
Oksitlenen substrattan hidrojeni ilk alan enzimlere birincil dehidrojenazlar denir.
Di- veya tripiridin nükleotidlerini içerirler: NAD veya NADP ve spesifik bir protein. Hidrojen ilavesinin mekanizması aynıdır:
Oksitlenebilir madde - H2 + NAD -> oksitlenmiş madde + NAD*H2
Dehidrojenaz tarafından üretilen hidrojen daha sonra bir sonraki enzim sistemi olan flavin enzimlerine (FMN veya FAD) eklenir.
Flavin enzimlerinden elektronlar sitokromlara - demir içeren proteinlere (kompleks proteinler) gider. Sitokrom zinciri boyunca aktarılan hidrojen atomu değil, yalnızca elektronlardır. Bu durumda demirin değerliği değişir: Fe++ - e->Fe++
Solunumun son reaksiyonu, havadaki oksijene bir proton ve bir elektronun eklenmesi ve suyun oluşmasıdır. Ancak önce oksijen molekülü, sitokrom oksidaz enziminin etkisi altında aktive edilir. Aktivasyon, oksitlenmiş maddeden bir elektronun eklenmesi nedeniyle oksijenin negatif bir yük kazanması gerçeğine iner. Hidrojen (proton) aktif oksijene bağlanarak su oluşturur.
Yukarıda bahsedilen elektron ve hidrojen taşıyıcıları zincirine ek olarak başkaları da bilinmektedir. Bu süreç, özetlenen diyagramdan çok daha karmaşıktır.
Bu dönüşümlerin biyolojik anlamı, maddelerin oksidasyonu ve enerji oluşumudur. Bir şeker molekülünün (glikoz) oksidasyonu sonucu ATP'de 12.6-1053 J enerji depolanır, şeker molekülünün kendisi 28.6-106 J içerir. J dolayısıyla enerjinin %44'ü faydalı bir şekilde kullanılır. Bu, modern makinelerin verimliliğiyle karşılaştırıldığında çok yüksek bir verimlilik faktörüdür.
Nefes alma süreci büyük miktarda enerji üretir. Eğer hepsi bir anda salınsaydı hücrenin varlığı sona ererdi. Ancak bu gerçekleşmez çünkü enerji bir anda değil, aşamalar halinde, küçük porsiyonlar halinde salınır. Enerjinin küçük dozlarda salınması, solunumun, bireysel aşamalarda çeşitli ara ürünlerin oluştuğu (farklı karbon zinciri uzunluklarında) ve enerjinin açığa çıktığı çok aşamalı bir süreç olmasından kaynaklanmaktadır. Açığa çıkan enerji, ısı biçiminde tüketilmez, ancak evrensel makroerjik bileşik olan ATP'de depolanır. ATP parçalandığında enerji, vücudun hayati işlevlerini sürdürmek için gerekli olan herhangi bir işlemde kullanılabilir: çeşitli organik maddelerin sentezi, mekanik çalışma, protoplazmanın ozmotik basıncının korunması vb. için.
Nefes almak enerji sağlayan bir süreçtir ancak biyolojik önemi bununla sınırlı değildir. Solunuma eşlik eden kimyasal reaksiyonlar sonucunda çok sayıda ara bileşik oluşur. Farklı sayıda karbon atomuna sahip olan bu bileşiklerden çok çeşitli hücre maddeleri sentezlenebilir: amino asitler, yağ asitleri, yağlar, proteinler, vitaminler.
Dolayısıyla karbonhidrat metabolizması diğer metabolizmaları (proteinler, yağlar) belirler. Bu onun büyük önemidir.
Mikropların şaşırtıcı özelliklerinden biri, solunum süreci ve onun kimyasal reaksiyonları (görünür ışık yayma yeteneği) ile ışıldama ile ilişkilidir.
Bakteriler de dahil olmak üzere birçok canlı organizmanın görünür ışık yayabildiği bilinmektedir. Mikroorganizmaların neden olduğu lüminesans yüzyıllardır bilinmektedir. Küçük deniz hayvanları ile simbiyoz halinde ışıldayan bakterilerin birikmesi bazen denizde bir parıltıya yol açar; Et vb. üzerinde belirli bakterilerin büyümesi sırasında da lüminesansla karşılaşıldı.
Etkileşimi ışık emisyonuna yol açan ana bileşenler, FMN veya NAD'nin indirgenmiş formlarını, moleküler oksijeni, lusiferaz enzimini ve oksitlenebilir bileşik - lusiferini içerir. İndirgenmiş NAD veya FMN'nin lusiferaz, oksijen ve lusiferin ile reaksiyona girdiği, bunun sonucunda bazı moleküllerdeki elektronların uyarılmış duruma geçtiği ve bu elektronların zemin seviyesine dönüşüne ışık emisyonunun eşlik ettiği varsayılmaktadır. Mikroplardaki lüminesans, solunumun enerji verimliliğini azalttığı için “israflı bir süreç” olarak kabul edilir.
Biyosferdeki organizmaların çoğu aerobik koşullardadır. Vücutta oksijen varlığında, karbonhidratların ve diğer "hücresel yakıt" moleküllerinin tamamen "yanması", nihai ürünlere - C0 2 ve H 2 0 - dönüşür.
Pirinç. 19.1. Glikozun altı CO2 molekülüne tamamen oksidasyonunun şeması ve bu sürecin enerji verimliliği (ATP dengesi); ATP oluşum yolları:
SF - substrat fosforilasyonu; OP - oksidatif fosforilasyon
Aerobik koşullar altında glikozun tamamen oksidasyonunun genel süreci stokiyometrik denklem ile tanımlanır.
Bu karmaşık, çok aşamalı glikoz oksidasyonu sürecinde üç aşama ayırt edilebilir (Şekil 19.1).
- İlk aşamada, glikozun iki piruvat molekülüne bölündüğü aerobik glikoliz reaksiyonları meydana gelir. Bu aşama karbonhidratların parçalanmasının başlangıç aşamasını oluşturur; buna “hazırlık” adı verilir.
- İkinci aşamada, piruvatın oksidatif dekarboksilasyonunun bir reaksiyon zinciri meydana gelir, bu da hücrenin merkezi metabolitlerinden biri olan asetil-S-CoA'nın oluşmasına ve piruvatın bir karbon atomunun C02'ye oksidasyonuna neden olur. Bir glikoz molekülü başına iki molekül piruvat oluştuğundan, bu aşamada glikozun iki karbon atomunun C02'ye oksidasyonu zaten meydana gelir.
- Üçüncü aşama, trikarboksilik asit döngüsü (TCA döngüsü) adı verilen, asetil kalıntısının tamamen oksidasyonuna ilişkin son derece önemli bir dizi reaksiyondur.
Karbonhidratların aerobik oksidasyon işlemine büyük miktarda enerjinin (2880 kJ/mol glikoz) salınması eşlik eder. Bu süreçte toplam ATP verimini toplarsak 38 molekül olacaktır (bkz. Şekil 19.1). Daha önce belirtildiği gibi (Bölüm 15), bir yüksek enerjili ATP bağının sentezi
38 ATP molekülünün sentezi için 31 kJ gereklidir ve 1178 kJ tüketilir, yani glikozun tam oksidasyonundan kaynaklanan serbest enerjinin %40'ından fazlası ATP moleküllerinde depolanır. Bu, anaerobik koşullarla karşılaştırıldığında aerobik koşullar altında meydana gelen oksidatif süreçlerin yüksek verimliliğini gösterir. Aerobik oksidasyon sürecinde, metabolik olarak mevcut enerji, indirgenmiş NADH ve FADH2 moleküllerinde birikir ve bunlar daha sonra oksijene bağımlı bir süreçte oksitlenir. oksidatif fosforilasyon bunun sonucu 34 ATP molekülünün oluşmasıdır ve substrat fosforilasyonuyla yalnızca 4 ATP molekülü oluşturulur: glikolizde 2ATP (aşama I) ve TCA döngüsünde 2ATP (2 dönüş, aşama III).
Karbonhidratların aerobik oksidasyonunun ilk aşaması - glikoliz - spesifik bir glikoz katabolizması süreci ise, sonraki iki - piruvatın oksidatif dekarboksilasyonu ve TCA döngüsünün genel katabolizma yollarına (GCP) ait olduğu belirtilmelidir. Sadece glikozun değil aynı zamanda lipitlerin ve amino asitlerin parçalanması sırasında oluşan piruvat (C3 fragmanı) ve asetil-CoA (C2 fragmanı) oluşumundan sonra, bu maddelerin nihai ürünlere oksidasyon yolları aynı şekilde meydana gelir. OPC reaksiyonlarının mekanizmasına göre.
Karbonhidratların aerobik oksidasyonu vücut için enerji üretiminin ana yoludur. Dolaylı - ikili ve doğrudan - aotomik.
Glikozun parçalanmasının doğrudan yolu pentoz döngüsü- pentoz oluşumuna ve NADPH 2 birikimine yol açar. Pentoz döngüsü, bir döngü sırasında 1 molekül karbondioksit ve suyun oluşmasıyla 6 karbon atomunun her birinin glikoz moleküllerinden sırayla elimine edilmesiyle karakterize edilir. Glikoz molekülünün tamamının parçalanması 6 tekrarlanan döngüden oluşur.
Karbonhidrat oksidasyonunun pentoz fosfat döngüsünün metabolizmadaki önemi büyüktür:
1. Yağ asitleri, kolesterol vb. biyosentezi için gerekli olan azaltılmış NADP'yi sağlar. Pentoz döngüsü nedeniyle vücudun NADPH 2 ihtiyacının %50'si karşılanır.
2. Nükleik asitlerin ve birçok koenzimin sentezi için pentoz fosfatların sağlanması.
Pentoz döngüsünün reaksiyonları hücrenin sitoplazmasında meydana gelir.
Bir dizi patolojik durumda, glikoz oksidasyonunun pentoz yolunun oranı artar.
Dolaylı yol- 36 molekül ATP'nin oluşmasıyla glikozun karbondioksit ve suya parçalanması.
1. Glikoz veya glikojenin pirüvik asite parçalanması
2. Pirüvik asidin asetil-CoA'ya dönüşümü
Krebs döngüsünde asetil-CoA'nın karbondioksit ve suya oksidasyonu
C 6 H 12 O 6 + 6 O 2 6 CO 2 + 6 H 2 O + 686 kcal
Aerobik dönüşüm durumunda piruvik asit, asetil-CoA'yı oluşturmak için oksidatif dekarboksilasyona uğrar ve bu daha sonra karbondioksit ve suya oksitlenir.
Piruvatın asetil-CoA'ya oksidasyonu piruvat dehidrojenaz sistemi tarafından katalize edilir ve birkaç aşamada meydana gelir. Toplam reaksiyon:
Piruvat + NADH + NS-CoA asetil-CoA + NADH2 + CO2 reaksiyonu pratik olarak geri döndürülemez
Asetil-CoA'nın tam oksidasyonu trikarboksilik asit döngüsünde veya Krebs döngüsünde meydana gelir. Bu süreç mitokondride gerçekleşir.
Döngü birbirini takip eden 8 reaksiyondan oluşur:
Bu döngüde, 2 karbon atomu içeren bir molekül (asetil-CoA formundaki asetik asit), bir oksaloasetik asit molekülü ile reaksiyona girer ve bunun sonucunda 6 karbon atomlu bir bileşik - sitrik asit oluşur. Hidrojen giderme, dekarboksilasyon ve hazırlık reaksiyonu sırasında sitrik asit, başka bir asetil-CoA molekülüyle kolayca birleşen oksaloasetik asite dönüştürülür.
1) asetil-CoA + oksaloasetat (OA) sitrik asit
sitrat sentaz
2) sitrik asit izositrik asit
akonitate hidrataz
3) izositrik asit + NAD-ketoglutarik asit + NADH2 + C02
izositrat dehidrojenaz
4)-ketoglutarik asit + NS-CoA + NADsüksinilSCoA + NADH2 + CO2
5) süksinil-CoA + GDP + Süksinik asit + GTP + HS-CoA
süksinil CoA sentetaz
6) süksinik asit + FAD fumarik asit + FADN 2
süksinat dehidrojenaz
7) fumarik asit + H20 L malik asit
fumarat hidrataz
8) malat + NADoksaloasetat + NADH 2
malat dehidrojenaz
Toplamda dokularda bir glikoz molekülü parçalandığında 36 ATP molekülü sentezlenir. Kuşkusuz bu, enerji açısından glikolizden daha verimli bir süreçtir.
Krebs döngüsü, karbonhidratların, yağ asitlerinin ve amino asitlerin metabolizmasının tamamlandığı ortak son yoldur. Tüm bu maddeler Krebs döngüsüne bir aşamada dahil edilir. Daha sonra biyolojik oksidasyon veya doku solunumu meydana gelir; bunun temel özelliği, çok sayıda enzimatik aşamadan geçerek kademeli olarak ilerlemesidir. Bu süreç, çok sayıda enzimin yoğunlaştığı hücresel organeller olan mitokondride meydana gelir. İşlem, piridin bağımlı dehidrojenazları, flavin bağımlı dehidrojenazları, sitokromları, koenzim Q - ubikinonu, hem olmayan demir içeren proteinleri içerir.
Solunum hızı ATP/ADP oranı tarafından kontrol edilir. Bu oran ne kadar düşük olursa, o kadar yoğun solunum gerçekleşir ve ATP üretimi sağlanır.
Ayrıca sitrik asit döngüsü, yağ asitlerinin ve glukoneojenezin sentezini başlatan karboksilasyon reaksiyonları için hücredeki ana karbondioksit kaynağıdır. Aynı karbondioksit, üre ve pürin ve pirimidin halkalarının bazı birimleri için karbon sağlar.
Karbonhidrat ve nitrojen metabolizması süreçleri arasındaki ilişki aynı zamanda sitrik asit döngüsünün ara ürünleri aracılığıyla da sağlanır.
Sitrik asit döngüsü ara maddelerinin lipogenez sürecine dahil edildiği çeşitli yollar vardır. Sitratın parçalanması, yağ asitlerinin biyosentezinde öncü rol oynayan asetil-CoA'nın oluşumuna yol açar.
İzositrat ve malat, yağ sentezinin sonraki indirgeyici aşamalarında tüketilen NADP oluşumunu sağlar.
NADH'nin dönüşümünü belirleyen anahtar faktörün rolü, adenin nükleotidlerinin durumu tarafından oynanır. Yüksek ADP ve düşük ATP, düşük enerji rezervlerini gösterir. Bu durumda NADH, enerji depolamayla ilişkili oksidatif fosforilasyon süreçlerini geliştirerek solunum zincirinin reaksiyonlarına dahil olur. Düşük ADP içeriğinde ve yüksek ATP içeriğinde ise tam tersi bir durum gözlenir. Elektron taşıma sistemini sınırlayarak, NADH'nin glutamat sentezi ve glukoneogenez gibi diğer indirgeyici reaksiyonlarda kullanımını teşvik ederler.
Atmosferdeki moleküler oksijenin biyojenik kökenli olduğu açıktır. Görünüşü, fotosentez sürecinde suyu hidrojen donörü olarak ilk kullanan en eski siyanobakterilerin veya atalarının fotosentez süreciyle ilişkilidir. Prokaryotların anaerobik oksidasyona geçişinin ancak belirli bir aşamada mümkün olduğu ortaya çıktı
evrim, hücrede az çok eksiksiz bir solunum zinciri oluştuğunda.
Çoğu aerobik prokaryotik organizma, enerji kaynağı olarak çeşitli organik bileşikleri tüketir ve bunları nihai ürünler CO2 ve H2O'ya oksitler. Prokaryotik bir hücrede organik maddelerin aerobik oksidasyonu, ökaryotların aerobik solunumuna benzer şekilde gerçekleşir. Pirüvik asidin trikarboksilik asit döngüsü (TCA - Krebs döngüsü) yoluyla oksidasyonuna dayanır.
Piruvik asidin Krebs döngüsüne dahil edilmesinden önce, piruvat dehidrojenaz kompleksi tarafından katalize edilen, asetil-K 0 A'ya oksidasyonunun karmaşık bir reaksiyonu gelir:
CH 3 – CO – COOH + K 0 A – SH + NAD + →CH 3 – CO ~ K 0 A + NAD H 2 + CO 2
Krebs döngüsünün kendisi, asetil-K 0 A'nın moleküler oksaloasetik asit ile sitras sentaz enziminin katılımıyla yoğunlaşma reaksiyonuyla başlar.
Krebs döngüsü hücre için iki önemli işlevi yerine getirir. Bu döngünün reaksiyonlarında, hidrojenin elimine edilmesi ve enzime aktarılmasıyla organik substrat tamamen oksitlenir; ayrıca hücreye öncü maddeler sağlanır.
Özetle Krebs döngüsü aşağıdaki denklemle ifade edilebilir:
CH3COCOOH + 2H20 → 3CO2 + 8H
Trikarboksilik asit döngüsünün başlangıç substratı sadece karbonhidratlar değil aynı zamanda yağ asitleri ve birçok amino asittir.
Krebs döngüsü solunum zinciriyle ilişkilidir. Solunum zincirinin temel işlevi, elektron transferi sırasında açığa çıkan enerjiyi ATP moleküllerindeki fosfat bağlarının kimyasal enerjisine dönüştürerek hücrede depolamaktır.
Aerobik prokaryotik organizmaların solunum zinciri şunları içerir: NAD dehidrojenazlar, FAD veya FMN dehidrojenazlar, ubikinon ve sitokrom sistemi. NAD dehidrojenazlar, oksitlenmiş substrattan hidrojenin çıkarılmasını ve bunun solunum zincirinin başlangıç taşıyıcıları olan NAD H2 dehidrojenaza transferini katalize eder. Bunlardan hidrojen, solunum zincirine FAD veya FMN dehidrojenazlara, ardından ubikinona ve ardından sitokrom sistemine aktarılır. Hidrojen solunum zinciri yoluyla aktarıldığında atomları protonlara ve elektronlara bölünür. Protonlar ortama salınır ve elektronlar solunum zinciri boyunca terminal taşıyıcıya - sitokrom oksidaza aktarılır. İkincisi, onları aktive edilen ve hidrojen ile birleşen son alıcıya - moleküler oksijene aktarır.
Elektronların solunum zinciri boyunca daha düşük ve daha düşük enerji seviyelerine transferi, hücre tarafından fosfat bağlarında biriken önemli miktarda serbest enerjinin salınmasına yol açar.
ATP moleküllerinin formu. Fosforilasyon reaksiyonları oksidasyon reaksiyonlarıyla eşleştiğinden bu işleme denir. oksidatif fosforilasyon. Elektron verici ve alıcının redoks potansiyeli arasındaki farka dayanır. ATP üretimi genellikle solunum zincirinin büyük potansiyel farklılıkları olan bölgelerinde meydana gelir. Örnek olarak ökaryotik mitokondri kullanılarak solunum zincirinde üç bölge tanımlanmıştır. İlk bölge, hidrojenin NAD H2 dehidrojenaz tarafından FAD veya FMN dehidrojenaza aktarılmasıyla ilişkilidir. İkinci bölüm, elektronları FAD veya FMN dehidrojenazdan sitokrom sistemine aktaran ubikinonun aktivitesi ile ilişkilidir. Üçüncü, son bölüm ise elektronların sitokrom oksidaz yoluyla moleküler oksijene aktarımı ile ilişkilidir.
Oksidatif fosforilasyon reaksiyonlarının etkinliği p/o oranı (emilen oksijenin bir atomu başına tüketilen inorganik fosfor moleküllerinin sayısı) ile değerlendirilir. Eokaryotlarda p/o oranı 3'tür. Birçok prokaryotta p/o oran üçten az olabilir, bu da solunum zincirinin bazı bölümlerinin kaybıyla açıklanabilir.
Aerobik prokaryotlar arasında, belirli organik maddelerin eksik aerobik oksidasyonu yoluyla enerji elde edebilen mikroorganizmalar vardır.
Asetik asit bakterileri küçük çubuklarla temsil edilir; genç bir kültürde hareketlidirler. Tüm türler zorunlu aeroblardır ve özellikle vitaminler ve öncelikle pantotenik asit olmak üzere substratlara oldukça ihtiyaç duyarlar. Bu gruptaki bakterilerin en karakteristik yeteneği, NAD'a bağımlı dehidrojenazların katılımıyla etil alkolü asetik asit oluşturmak üzere oksitlemektir.
Eksik anaerobik oksidasyon işlemlerinin fermantasyon işlemleriyle hiçbir ortak yanının olmadığını vurguluyoruz. Oksitlenmiş fosforasyon reaksiyonlarında hücre yaşamı için enerji üretilir, ancak bu enerjinin bir kısmı az oksitlenmiş son ürünlerde depolandığından, tam aerobik oksidasyondan daha küçük miktarlarda olur.
Çeşitli asetik asit bakteri türleri, hem monohidrik alkolleri hem de polihidrik alkolleri (şeker türevleri) oksitlenebilir bir substrat olarak kullanma yeteneğine sahiptir. Monohidrik alkollerin oksidasyonu çeşitli asitler üretir. Polihidrik alkollerin oksidasyonu altoz ve ketoz oluşumuna yol açar. Endüstride asetik asit bakterileri sofra sirkesi ve askorbik asit üretmek için kullanılır.
Solunumu yalnızca organik maddelerin oksidasyonu yoluyla gerçekleştiren ökaryotların aksine, prokaryotlar arasında, substrattaki inorganik maddeleri katabolizma süreci yoluyla oksitleyebilen kemolitotrofik mikroorganizma grupları vardır.
Kemolitotrofik mikroorganizmaların solunum zinciri, temel olarak kemoorganotrofların solunum zinciriyle aynı elektron transfer enzimlerini içerir. Bu mikroorganizma grubunun solunum zincirinin işleyişinin özgüllüğü, farklı redoks potansiyellerine sahip inorganik bileşiklerin oksidasyonu sırasında, oksitlenmiş substrattan gelen elektronların farklı enerji seviyelerinde solunum zincirine dahil edilmesidir. Bu nedenle, hücreye enerji sağlamak için mikroorganizmalar büyük miktarda substratı oksitlemeye zorlanır.
İnorganik maddelerin oksidasyonu yoluyla enerji elde eden kemolitotrofik mikroorganizmalar arasında nitrifikasyon bakterileri, demir bakterileri, tiyonik bakteriler vb. bulunur.
Metabolik kararsızlık açısından karboksibakteriler özellikle ilgi çekicidir. Bu mikroorganizmalar, tek karbon ve enerji kaynağı olarak karbon monoksit (CO) tüketen ototroflar olarak ve karbon ve enerji kaynağı olarak organik maddeleri (alkoller ve organik asitler) kullanan heterotroflar gibi davranabilirler.
Ototrofik karboksidobakterilerin toplam katabolizma ve anabolizma süreçleri aşağıdaki denklemle temsil edilebilir:
24CO + 11O2 + H2O → 23CO2 + (CH2O),
burada (CH 2 O) biyokütlenin sembolüdür.
Denklemden, CO oksidasyonunun enerji elde etmenin etkisiz bir yolu olduğu, dolayısıyla mikroorganizmaların büyük miktarda substratı oksitlemeye zorlandığı sonucu çıkar.
Modern ulaşım ve endüstriyel işletmelerin CO2 salımı, bu bileşikle atmosferi kirletmektedir. CO'yu ortamdan uzaklaştırmanın tek yolu onu mikrobiyal hücrelerin metabolizmasında kullanmaktır.
Anaerobik oksidasyon: nitrat ve sülfat solunumu
Anaerobik oksidasyon yalnızca prokaryotik krallığın temsilcileri arasında meydana gelir. Son elektron alıcısı olarak hem moleküler oksijeni hem de nitratlardan nitrojeni ve sülfatlardan kükürdü kullanarak aerobik bir yaşam tarzından anaerobik bir yaşam tarzına geçiş yapabilen mikroorganizmaların doğasında vardır.
Bu tür mikroorganizmaların tipik bir örneği denitrifikasyon bakterileridir.
Denitrifikasyon bakterilerinin solunum zinciri, aerobik solunum zincirinin karakteristik tüm önemli elektron transfer enzimlerini içerir. Sitokrom sisteminin yalnızca son halkası olan sitokrom oksidazın yerini, elektronların nitrat nitrojene transferini katalize eden nitrat redüktaz alır. Nitrat redüktazlar, ortamdaki nitratların varlığında yalnızca anaerobik koşullar altında hücreler tarafından sentezlenen indüklenebilir enzimlerdir.
Denitrifikasyon prosesi, her biri karşılık gelen nitrat redüktaz tarafından katalize edilen 4 indirgeme aşamasından oluşur. İlk aşamada nitratlar nitritlere indirgenir:
nitrojen +5, 2 proton ve 2 elektron alarak nitrit nitrojen NO 2 - +3'e indirgenir:
NO 3 - + 2e - + 2H + →NO 2 - + H 2 O.
HAYIR 2 - + e - + H + → HAYIR + OH -
2NO + 2e - + 2H + → N2O + H2O
N 2 O + 2e - + 2H + →N 2 + H 2 O
Azotun elektron alıcısı olarak kullanılması, denitrifikasyon bakterilerinin substrattaki organik maddeleri nihai ürünler CO2 ve H2O'ya tamamen oksitlemesine olanak tanır. Bu nedenle, nitrat solunumunun enerji verimi geleneksel aerobik oksidasyona neredeyse yakındır.
Denitrifikasyon bakterileri yalnızca anaerobik koşullara maruz kaldıklarında nitrat solunumuna geçtiklerinden, anaerobik yaşam tarzına adaptasyonları evrimsel olarak ikincil olarak kabul edilmeli ve tipik aerobik oksidasyondan anaerobiyoza dönüş olarak değerlendirilmelidir.
Desulfotomaculum, Desulfonema, Desulfovibrio vb. cinslerine ait sülfat indirgeyen bakteriler de anaerobik oksidasyon yeteneğine sahiptir. Sülfat indirgeyen bakterilerin enerji elde etme yolları farklı olabilir. Bu, organik maddelerin anaerobik koşullar altında elektronların sülfat kükürte aktarılmasıyla oksidasyonunu içeren substrat fosforilasyonu, sülfat solunumu sonucu ATP oluşumunun eşlik ettiği organik maddelerin fermantasyon sürecidir. Bu heterojen grubun bakterileri ayrıca sülfatların indirgenmesiyle birlikte moleküler hidrojenin oksidasyonu yoluyla enerji elde etme yeteneğine de sahiptir.
Sülfat indirgeyen bakterilerin enerji üretmek için moleküler hidrojeni kullanma yeteneği, bunların anaerobik kemolitotrofik mikroorganizmalar olarak sınıflandırılmasına olanak tanır.
Moleküler hidrojenin oksidasyonu sürecinde metan üreten bakteriler de karbondioksiti elektron alıcısı olarak kullanarak enerji elde ederler. Bu gruptaki bakteriler için CO2 hem karbon kaynağı hem de elektron alıcısı olarak görev yapar:
4H2 + C02 →CH4 + 2H20
Çeşitli prokaryotik katabolizma türlerinin incelenmesi, bu krallığın temsilcilerinin evriminin altında yatan şeyin, hücrenin enerji elde etme yollarının iyileştirilmesi olduğunu varsaymayı mümkün kılar.
Prokaryotların en eski grubu, substrat fosforilasyonu nedeniyle fermantasyon süreçlerinde enerji üreten anaerobik bakterilerdir.
Prokaryotların evrimindeki önemli bir aşama, ana enerji kaynağı olarak güneş ışığını ve ana karbon kaynağı olarak CO2'yi kullanan fototrofik bakterilerin ortaya çıkışı olarak düşünülmelidir.
Başta siyanobakteriler olmak üzere fotosentetik aerobların gelişimi, çevrenin moleküler oksijenle zenginleşmesine yol açtı. Aerobik bakteri hücresinde başka bir elektron taşıma sistemi gelişmiştir ve bununla ilişkili bir fosforilasyon mekanizması oksidatif fosforilasyondur.
Şu anda prokaryotların krallığında inanılmaz çeşitlilikte katabolizma türleriyle karşılaşıyoruz. Bununla birlikte, baskın ve evrimsel olarak baskın katabolizma türü, tüm donör ve alıcı çeşitliliğiyle birlikte şüphesiz aerobik oksidasyondur.