Bitta oqsilning tuzilishi genlar guruhi bilan belgilanadi. Oqsillarning strukturaviy tashkil etilishi. Eukariotlarda transkripsiya sodir bo'ladi
Biologik kimyo Lelevich Vladimir Valeryanovich
Oqsillarning strukturaviy tashkiliy darajalari
Birlamchi tuzilma- polipeptid zanjiridagi aminokislotalarning qat'iy belgilangan chiziqli ketma-ketligi.
Oqsillarning birlamchi tuzilishini o‘rganishning strategik tamoyillari qo‘llanilgan usullarning rivojlanishi va takomillashishi natijasida sezilarli o‘zgarishlarga uchradi. Ularning rivojlanishidagi uchta asosiy bosqichni ta'kidlash kerak. Birinchi bosqich F. Sangerning (1953) insulinning aminokislotalar ketma-ketligini o'rnatish bo'yicha klassik ishi bilan boshlanadi, ikkinchisi - oqsillarni strukturaviy tahliliga avtomatik sekvenserni keng joriy etish bilan (20-asrning 70-yillari boshlari), uchinchisi - DNKning nukleotidlar ketma-ketligini tahlil qilishning yuqori tezlikdagi usullarini ishlab chiqish bilan (20-asrning 80-yillari boshlari).
Proteinning asosiy tuzilishi quyidagilar bilan belgilanadi:
1. Molekula tarkibiga kirgan aminokislotalarning tabiati.
2. Har bir aminokislotaning nisbiy miqdori.
3. Polipeptid zanjiridagi aminokislotalarning qat'iy belgilangan ketma-ketligi.
Proteinning birlamchi tuzilishini aniqlashdan oldin dastlabki tadqiqotlar
1. Proteinni tozalash
2. Molekulyar massani aniqlash.
3. Protez guruhlarning turi va sonini aniqlash (agar oqsil konjugatsiyalangan bo'lsa).
4. Molekulyar yoki molekulalararo disulfid bog'lanishlar mavjudligini aniqlash. Odatda, mahalliy oqsilda sulfhidril guruhlarning mavjudligi bir vaqtning o'zida aniqlanadi.
5. Subbirliklarni dissotsiatsiyalash, ularni ajratish va keyinchalik o'rganish maqsadida 4-chi tuzilishga ega bo'lgan oqsillarni oldindan davolash.
Oqsillar va polipeptidlarning birlamchi tuzilishini aniqlash bosqichlari
1. Aminokislota tarkibini aniqlash (gidroliz, aminokislota analizatori).
2. N- va C-terminal aminokislotalarni aniqlash.
3. Polipeptid zanjirining bo'laklarga bo'linishi (tripsin, ximotripsin, sianogen bromid, gidroksilamin va boshqalar).
4. Peptid fragmentlarining aminokislotalar ketma-ketligini aniqlash (sekvenser).
5. Dastlabki polipeptid zanjirini boshqa usullar bilan parchalash va ularning aminokislotalar ketma-ketligini aniqlash.
6. Peptid fragmentlarini bir-birining ustiga chiqadigan joylarda joylashtirish tartibini belgilash (peptid xaritalarini olish).
N-terminal aminokislotalarni aniqlash usullari
1. Sanger usuli.
2. Edman usuli (sequencerda amalga oshiriladi).
3. Dansilxlorid bilan reaksiyasi.
4. Aminopeptidazadan foydalanish usuli.
C-terminal aminokislotalarni aniqlash usullari
1. Akabori usuli.
2. Karboksipeptidazadan foydalanish usuli.
3. Natriy borgidriddan foydalanish usuli.
Oqsillarning aminokislotalar ketma-ketligiga oid umumiy naqshlar
1. Barcha oqsillarga xos bo'lgan yagona ketma-ketlik yoki qisman ketma-ketliklar guruhi mavjud emas.
2. Turli funktsiyalarni bajaradigan oqsillar turli xil ketma-ketlikka ega.
3. O'xshash funktsiyalarga ega bo'lgan oqsillar o'xshash ketma-ketliklarga ega, lekin odatda faqat kichik darajadagi ketma-ketliklarning bir-biriga o'xshashligi mavjud.
4. Bir xil funktsiyalarni bajaradigan, lekin turli organizmlardan ajratilgan bir xil oqsillar, odatda, muhim ketma-ketlik o'xshashligiga ega.
5. Xuddi shu funktsiyalarni bajaradigan va bir xil turdagi organizmlardan ajratilgan bir xil oqsillar deyarli doimo bir xil ketma-ketlikka ega.
Protein tuzilishining eng yuqori darajalari va ularning biologik faolligi bir-biri bilan chambarchas bog'liq va aslida aminokislotalar ketma-ketligi bilan belgilanadi. Ya'ni, birlamchi struktura genetik jihatdan aniqlanadi va oqsillarning individual xususiyatlarini, ularning tur o'ziga xosligini belgilaydi, uning asosida barcha keyingi tuzilmalar shakllanadi.
Proteinning ikkilamchi tuzilishi - bu uning funktsional guruhlari orasidagi o'zaro ta'sir natijasida yuzaga keladigan polipeptid zanjirining konfiguratsiyasi.
Ikkilamchi tuzilish turlari:
1. ?-spiral.
2. Buklangan varaq (?-struktura).
3. Statistik chalkashlik.
Birinchi ikkita nav tartibli tartibni, uchinchisi - tartibsiz.
Oqsillarning ikkinchi darajali tuzilishi.
Turli tuzilish va funktsiyalarga ega bo'lgan oqsillarning konformatsiyasini taqqoslash ularda ikkilamchi tuzilish elementlarining o'xshash birikmalarining mavjudligini aniqladi. Ikkilamchi tuzilmalarni shakllantirishning bu o'ziga xos tartibi supero'rta tuzilma deb ataladi. Supero'rta tuzilma radikallararo o'zaro ta'sirlar tufayli shakllanadi.
Oqsillarning ikkinchi darajali tuzilishi turlari:
1.?-barrel tipidagi o'ta ikkilamchi tuzilishi. U haqiqatan ham barrelga o'xshaydi, bu erda har bir?-struktura ichida joylashgan va sirtda joylashgan zanjirning?-spiral qismi bilan bog'langan. Ba'zi fermentlarning xarakteristikasi - triosefosfat izomeraza, piruvat kinaz.
2. Strukturaviy motiv “?-spiral – burilish – ?-spiral”. Ko'pgina DNKni bog'laydigan oqsillarda topilgan.
3. "Sink barmog'i" shaklida o'ta ikkinchi darajali tuzilish. Shuningdek, DNKni bog'lovchi oqsillarga xosdir. "Rux barmog'i" 20 ga yaqin aminokislotalarni o'z ichiga olgan oqsil bo'lagi bo'lib, unda sink atomi to'rtta aminokislota radikallari bilan bog'langan: odatda ikkita sistein qoldig'i va ikkita gistidin qoldig'i.
4. Leysin fermuar supersekondar tuzilishi. Protomerlar yoki alohida oqsillarni komplekslarga birlashtirish ba'zan "leytsin fermuarlar" deb ataladigan strukturaviy motivlar yordamida amalga oshiriladi. Bunday oqsil birikmasiga gistonlar misol bo'la oladi. Bu juda ko'p miqdordagi musbat zaryadlangan aminokislotalar - arginin va lizinni o'z ichiga olgan yadro oqsillari. Giston molekulalari barcha monomerlar kuchli musbat zaryadga ega bo'lishiga qaramay, "leysin fermuarlar" yordamida murakkablashadi.
a-spirallar va b-tuzilmalarning mavjudligiga ko'ra, globulyar oqsillarni 4 toifaga bo'lish mumkin:
Proteinning uchinchi darajali tuzilishi polipeptid zanjirining fazoviy yo'nalishi yoki ma'lum hajmda buklanish usulidir.
Uchinchi darajali strukturaning shakliga qarab, globulyar va fibrillar oqsillar farqlanadi. Globulyar oqsillarda ko'pincha a-spiral ustunlik qiladi, fibrillyar oqsillar a-tuzilma asosida hosil bo'ladi.
Globulyar oqsilning uchinchi darajali tuzilishini barqarorlashtirishda quyidagilar ishtirok etishi mumkin:
1. spiralsimon strukturaning vodorod aloqalari;
2. vodorod boglari?-tuzilmalar;
3. yon zanjirli radikallar orasidagi vodorod aloqalari;
4. qutbsiz guruhlar orasidagi hidrofobik o'zaro ta'sirlar;
5. qarama-qarshi zaryadlangan guruhlar orasidagi elektrostatik o'zaro ta'sirlar;
6. disulfid aloqalari;
7. metall ionlarining koordinatsion aloqalari.
Proteinning to'rtlamchi tuzilishi - bu bir xil (yoki boshqacha) birlamchi, ikkilamchi yoki uchinchi darajali tuzilishga ega bo'lgan alohida polipeptid zanjirlarini kosmosda joylashtirish va strukturaviy va funktsional jihatdan birlashtirilgan makromolekulyar shakllanishni shakllantirish usuli.
To'rtlamchi tuzilish bir nechta bo'linmalardan tashkil topgan oqsillarga xosdir. To'rtlamchi tuzilishdagi bo'linmalarning to'ldiruvchi mintaqalari o'rtasidagi o'zaro ta'sir vodorod va ion aloqalari, van der Vaals kuchlari va hidrofobik o'zaro ta'sirlar yordamida amalga oshiriladi. Kovalent bog'lanishlar kamroq uchraydi.
Bir uzun polipeptid zanjiri bilan solishtirganda, subunit protein konstruktsiyasining afzalliklari.
Birinchidan, subunit tuzilmasining mavjudligi genetik materialni "saqlash" imkonini beradi. Bir xil bo'linmalardan tashkil topgan oligomerik oqsillar uchun strukturaviy genning o'lchami va shunga mos ravishda xabarchi RNKning uzunligi keskin kamayadi.
Ikkinchidan, nisbatan kichik zanjir hajmi bilan oqsil molekulalarining biosintezi jarayonida yuzaga kelishi mumkin bo'lgan tasodifiy xatolarning ta'siri kamayadi. Bundan tashqari, subbirliklarni yagona kompleksga birlashtirishda "noto'g'ri", noto'g'ri polipeptidlarni qo'shimcha ravishda rad etish mumkin.
Uchinchidan, ko'pgina oqsillarda subbirlik strukturasi mavjudligi hujayraning assotsiatsiya-dissosiatsiya muvozanatini u yoki bu tomonga siljishi orqali o'z faoliyatini osonlik bilan tartibga solish imkonini beradi.
Nihoyat, subunit strukturasi molekulyar evolyutsiya jarayonini osonlashtiradi va tezlashtiradi. To'rtlamchi tuzilishga o'tish jarayonida bu o'zgarishlarning ko'p marta kuchayishi tufayli uchinchi darajali tuzilish darajasida faqat kichik konformatsion o'zgarishlarga olib keladigan mutatsiyalar oqsilda yangi xususiyatlarning paydo bo'lishiga yordam beradi.
Biologiya kitobidan [Yagona davlat imtihoniga tayyorgarlik ko'rish uchun to'liq ma'lumotnoma] muallif Lerner Georgiy Isaakovich "Yo'qolgan bo'g'in" kitobidan Edie Maitland tomonidanOila daraxti (oqsillarning dalillari) Oila daraxti (oqsillarning dalillari) Ikki turning oqsillaridagi farqlar bu turlarning umumiy ajdoddan ajralganidan keyin evolyutsion o'zgarishlarini aks ettiradi. Tahlil shuni ko'rsatadiki, shimpanze qon zardobida albuminlar o'rtasida
Hayot haqida suhbatlar kitobidan muallif Galaktionov Stanislav Gennadievich2-bob. Oqsillarning molekulyar arxitekturasi Yashirmaylik: birinchi bobni tugatgandan so'ng mualliflar (ehtimol, o'quvchi) biroz yengillik his qilishdi. Axir, uning maqsadi faqat o'quvchiga keyingi boblarni tushunish uchun zarur bo'lgan ma'lumotlarni berish edi,
Evolyutsiya kitobidan [Yangi kashfiyotlar nurida klassik g'oyalar] muallifQadimgi oqsillar olami kengayishda davom etmoqda 2010 yilda Nature jurnali fitnes landshaftlari bo'ylab oqsillarning evolyutsion harakati haqida qiziqarli maqola chop etdi (Povolotskaya, Kondrashov, 2010). Ish mualliflari 572 ta qadimgi oqsillarning aminokislotalar ketma-ketligini solishtirishga qaror qilishdi.
"Genlar va tananing rivojlanishi" kitobidan muallif Neyfax Aleksandr Aleksandrovich4. Strukturaviy gipoteza variantlari Shunday qilib, bir nechta eksperimental ma'lumotlar mitoz va replikatsiya jarayonida saqlanib qoladigan, bir qator hujayra avlodlari orqali uzatilishi va epigenetik ta'sir ko'rsatishi mumkin bo'lgan bunday tarkibiy o'zgarishlarning imkoniyatini ko'rsatadi.
Inson evolyutsiyasi kitobidan. Kitob 1. Maymunlar, suyaklar va genlar muallif Markov Aleksandr VladimirovichProtein o'zgarishi Genomning oqsillarni kodlaydigan qismlari hayratlanarli darajada kam o'zgargan. Odamlar va shimpanzelar o'rtasidagi oqsillarning aminokislotalar ketma-ketligidagi farqlar sezilarli darajada 1% dan kamroq va hatto bu bir nechta farqlarning ko'pchiligida hech qanday farq yo'q.
Biologiya kitobidan. Umumiy biologiya. 10-sinf. Asosiy daraja muallif Sivoglazov Vladislav Ivanovich3. Tirik materiyaning tashkiliy darajalari. Biologiya metodlari Esda tuting!Tirik materiyaning qanday tashkiliy darajalarini bilasiz?Qanday ilmiy tadqiqot usullarini bilasiz?Tirik materiyaning tashkiliy darajalarini. Atrofimizdagi tirik mavjudotlar dunyosi biologik tizimlar to'plamidir
"Antropologiya va biologiya tushunchalari" kitobidan muallif Kurchanov Nikolay AnatolievichHayotni tashkil etishning strukturaviy-funksional darajalari Biologiyada tirik materiyani tashkil qilishning bir qancha strukturaviy va funksional darajalari mavjud.Molekulyar daraja. Tirik organizmni tashkil etuvchi biokimyoviy moddalar bilan tavsiflanadi Hujayra darajasi.
Biologik kimyo kitobidan muallif Lelevich Vladimir Valeryanovich2-bob.Oqsillarning tuzilishi va funktsiyalari Proteinlar yuqori molekulyar azotli organik birikmalar bo'lib, peptid bog'lari yordamida polipeptid zanjirlariga bog'langan aminokislotalardan iborat va murakkab tuzilishga ega.Oqsillarni o'rganish tarixi 1728 y.
Muallifning kitobidanProteinlarning ishlashi Noyob birlamchi tuzilishi va konformatsiyasiga ega bo'lgan har bir alohida oqsil, shuningdek, uni boshqa barcha oqsillardan ajratib turadigan o'ziga xos funktsiyaga ega. Alohida oqsillar to'plami hujayrada juda ko'p turli xil va murakkab vazifalarni bajaradi.
Muallifning kitobidanOqsillardagi translatsiyadan keyingi o'zgarishlar Ko'pgina oqsillar faol bo'lmagan shaklda (prekursorlar) sintezlanadi va ribosomalar bilan konvergentsiyadan so'ng, postsintetik strukturaviy o'zgarishlarga uchraydi. Qabul qilingan polipeptid zanjirlarida bu konformatsion va strukturaviy o'zgarishlar
Muallifning kitobidanModdalar almashinuvini o'rganish darajalari Moddalar almashinuvini o'rganish darajalari:1. Butun organizm.2. Izolyatsiya qilingan organlar (perfuziyalangan).3. To‘qima bo‘limlari.4. Hujayra kulturalari.5. To‘qimalarning gomogenatlari.6. Izolyatsiya qilingan hujayra organellalari.7. Molekulyar daraja (tozalangan fermentlar, retseptorlar va
Muallifning kitobidanOqsillarning oshqozon-ichak traktida hazm bo'lishi Oqsillarning hazm bo'lishi me'da shirasidagi fermentlar ta'sirida oshqozonda boshlanadi. Kuniga 2,5 litrgacha ajraladi va u yuqori kislotaliligi bilan boshqa ovqat hazm qilish sharbatlaridan farq qiladi
Muallifning kitobidanTo'qimalarda oqsillarning parchalanishi proteolitik lizosomal fermentlar katepsinlari yordamida amalga oshiriladi. Faol markazning tuzilishiga ko'ra sistein, serin, karboksil va metalloprotein katepsinlari ajralib turadi. Katepsinlarning roli: 1. biologik faol moddalarni yaratish
Muallifning kitobidanJigarning aminokislotalar va oqsillar almashinuvidagi roli Jigar oqsillar va boshqa azotli birikmalar almashinuvida markaziy rol o'ynaydi. U quyidagi funktsiyalarni bajaradi: 1. o'ziga xos plazma oqsillari sintezi: - jigarda sintezlanadi: 100% albuminlar, 75-90% b-globulinlar, 50%
Muallifning kitobidanQon zardobi oqsillarining xarakteristikalari Komplement tizimining oqsillari - bu tizim qonda faol bo'lmagan prekursorlar shaklida aylanib yuradigan 20 ta oqsilni o'z ichiga oladi. Ularning faollashishi proteolitik faollikka ega bo'lgan o'ziga xos moddalar ta'sirida sodir bo'ladi.
Proteinlarning kimyoviy tuzilishi zanjirda peptid bog'i orqali bog'langan alfa aminokislotalar bilan ifodalanadi. Tirik organizmlarda tarkibi genetik kod bilan belgilanadi. Sintez jarayonida ko'p hollarda standart turdagi 20 ta aminokislotalar ishlatiladi. Ularning ko'p kombinatsiyasi turli xil xususiyatlarga ega bo'lgan oqsil molekulalarini hosil qiladi. Aminokislota qoldiqlari ko'pincha translatsiyadan keyingi o'zgarishlarga duchor bo'ladi. Ular oqsil o'z funktsiyalarini bajarishni boshlashdan oldin va hujayradagi faoliyati davomida paydo bo'lishi mumkin. Tirik organizmlarda bir nechta molekulalar ko'pincha murakkab komplekslarni hosil qiladi. Masalan, fotosintetik assotsiatsiya.
Ulanishlarning maqsadi
Proteinlar inson va hayvonlarning oziqlanishining muhim tarkibiy qismi hisoblanadi, chunki ularning tanasi barcha kerakli aminokislotalarni sintez qila olmaydi. Ulardan ba'zilari proteinli ovqatlar bilan birga kelishi kerak. Aralashmalarning asosiy manbalari go'sht, yong'oq, sut, baliq va don hisoblanadi. Kamroq darajada oqsillar sabzavot, qo'ziqorin va reza mevalarida mavjud. Fermentlar orqali hazm qilish jarayonida iste'mol qilingan oqsillar aminokislotalarga bo'linadi. Ular allaqachon tanadagi o'z oqsillarining biosintezida ishlatiladi yoki energiya olish uchun keyingi parchalanishga uchraydi.

Tarixiy ma'lumotnoma
Insulin oqsili tuzilishi ketma-ketligi birinchi marta Frederik Senger tomonidan aniqlangan. Bu ishi uchun u 1958 yilda Nobel mukofotiga sazovor bo'lgan. Sanger sekvensiya usulidan foydalangan. Keyinchalik rentgen nurlari diffraktsiyasidan foydalanib, miyoglobin va gemoglobinning uch o'lchovli tuzilmalari olingan (1950-yillarning oxirida). Ish Jon Kendrew va Maks Perutz tomonidan amalga oshirildi.
Protein molekulasi tuzilishi
U chiziqli polimerlarni o'z ichiga oladi. Ular, o'z navbatida, monomerlar bo'lgan alfa aminokislotalar qoldiqlaridan iborat. Bundan tashqari, oqsil tuzilishi aminokislota bo'lmagan tabiatning tarkibiy qismlarini va o'zgartirilgan aminokislota qoldiqlarini o'z ichiga olishi mumkin. Komponentlarni belgilashda 1 yoki 3 harfli qisqartmalar qo'llaniladi. Ikki dan bir necha o'nlab qoldiqlarni o'z ichiga olgan birikma ko'pincha "polipeptid" deb ataladi. Bir aminokislotaning alfa-karboksil guruhining boshqasining alfa-amino guruhi bilan o'zaro ta'siri natijasida bog'lar paydo bo'ladi (oqsil tuzilishini shakllantirish jarayonida). Aminokislota qoldig'ining qaysi guruhi erkin bo'lishiga qarab birikmaning C- va N-terminal uchlari farqlanadi: -COOH yoki -NH 2 . Ribosomada oqsil sintezi jarayonida birinchi terminal qoldiq odatda metionin qoldig'i bo'ladi; keyingilari oldingilarning C-terminusiga biriktirilgan.

Tashkilot darajalari
Ular Lindrem-Lang tomonidan taklif qilingan. Ushbu bo'linma biroz eskirgan deb hisoblanishiga qaramay, u hali ham qo'llaniladi. Ulanishni tashkil etishning to'rtta darajasini ajratish taklif qilindi. Protein molekulasining asosiy tuzilishi genning genetik kodi va xususiyatlari bilan belgilanadi. Yuqori darajalar oqsillarni katlama paytida hosil bo'lishi bilan tavsiflanadi. Proteinning fazoviy tuzilishi aminokislotalar zanjiri bilan bir butun sifatida aniqlanadi. Shunga qaramay, bu juda o'zgaruvchan. Bunga tashqi omillar ta'sir qilishi mumkin. Shu munosabat bilan, eng maqbul va energiya jihatidan afzalroq bo'lgan birikmaning konformatsiyasi haqida gapirish to'g'riroq.
1-daraja
U polipeptid zanjirining aminokislotalar qoldiqlari ketma-ketligi bilan ifodalanadi. Qoida tariqasida, u bir yoki uch harfli belgilar yordamida tasvirlanadi. Oqsillarning birlamchi tuzilishi aminokislotalar qoldiqlarining barqaror birikmalari bilan tavsiflanadi. Ular aniq vazifalarni bajaradilar. Bunday "konservativ motivlar" turlar evolyutsiyasi davrida saqlanib qoladi. Ular ko'pincha noma'lum protein muammosini taxmin qilish uchun ishlatilishi mumkin. Turli organizmlarning aminokislotalar zanjiridagi o'xshashlik (homologiya) darajasini baholab, bu organizmlarni tashkil etuvchi taksonlar o'rtasida hosil bo'lgan evolyutsion masofani aniqlash mumkin. Oqsillarning birlamchi tuzilishi genetik kodlar jadvali yordamida sekvensiyalash yoki uning mRNKning asl kompleksi bilan aniqlanadi.

Zanjir qismini mahalliy buyurtma qilish
Bu tashkilotning navbatdagi darajasi - oqsillarning ikkilamchi tuzilishi. Uning bir necha turlari mavjud. Polipeptid zanjirining bir qismining mahalliy tartibi vodorod aloqalari bilan barqarorlashadi. Eng mashhur turlari quyidagilardir:

Fazoviy tuzilma
Oqsillarning uchinchi darajali tuzilishi oldingi darajadagi elementlarni o'z ichiga oladi. Ular har xil turdagi o'zaro ta'sirlar bilan barqarorlashadi. Gidrofobik aloqalar katta ahamiyatga ega. Stabilizatsiya quyidagilarni o'z ichiga oladi:
- Kovalent o'zaro ta'sirlar.
- Qarama-qarshi zaryadga ega bo'lgan aminokislotalar yon guruhlari o'rtasida hosil bo'lgan ion aloqalari.
- Vodorodning o'zaro ta'siri.
- Gidrofobik aloqalar. Atrofdagi H 2 O elementlari bilan o'zaro ta'sir qilish jarayonida oqsil yon polar bo'lmagan aminokislota guruhlari suvli eritmadan ajratilishi uchun burmalanadi. Molekula yuzasida gidrofil guruhlar (polyar) paydo bo'ladi.
Oqsillarning uchinchi darajali tuzilishi magnit (yadro) rezonans usullari, mikroskopning ayrim turlari va boshqa usullar bilan aniqlanadi.
O'rnatish printsipi
Tadqiqotlar shuni ko'rsatdiki, 2 va 3 darajalar orasida yana bir darajani aniqlash qulay. U "arxitektura", "qo'yish motivi" deb ataladi. U ikkilamchi strukturaning tarkibiy qismlarining (beta iplari va alfa spirallari) ixcham globula - oqsil sohasi chegaralaridagi nisbiy joylashuvi bilan belgilanadi. U mustaqil ravishda mavjud bo'lishi yoki boshqa shunga o'xshash oqsillar bilan birga kattaroq protein tarkibiga kirishi mumkin. Styling motivlari ancha konservativ ekanligi aniqlandi. Ular na evolyutsion, na funktsional munosabatlarga ega bo'lmagan oqsillarda mavjud. Arxitekturaning ta'rifi ratsional (jismoniy) tasniflashning asosidir.

Domen tashkiloti
Bitta protein kompleksi ichida bir nechta polipeptid zanjirlarining o'zaro joylashishi bilan oqsillarning to'rtlamchi tuzilishi hosil bo'ladi. Uni tashkil etuvchi elementlar ribosomalarda alohida hosil bo'ladi. Faqat sintez tugagandan so'ng, bu protein tuzilishi shakllana boshlaydi. U turli xil va bir xil polipeptid zanjirlarini o'z ichiga olishi mumkin. Oqsillarning to'rtlamchi tuzilishi avvalgi darajadagi o'zaro ta'sirlar tufayli barqarorlashadi. Ba'zi komplekslar bir necha o'nlab oqsillarni o'z ichiga olishi mumkin.
Protein tuzilishi: himoya vazifalari
Tsitoskeletning polipeptidlari qaysidir ma'noda mustahkamlovchi rolini o'ynaydi, ko'plab organellalarga o'z shakllarini beradi va uning o'zgarishida ishtirok etadi. Strukturaviy oqsillar tanani himoya qiladi. Masalan, kollagen shunday oqsildir. U biriktiruvchi to'qimalarning hujayralararo moddasida asosni tashkil qiladi. Keratin ham himoya funktsiyasiga ega. U epidermisning shoxlari, patlari, sochlari va boshqa hosilalarining asosini tashkil qiladi. Proteinlar toksinlarni bog'laganda, ko'p hollarda detoksifikatsiya sodir bo'ladi. Tanani kimyoviy himoya qilish vazifasi shu tarzda amalga oshiriladi. Jigar fermentlari inson organizmidagi toksinlarni zararsizlantirish jarayonida ayniqsa muhim rol o'ynaydi. Ular zaharlarni parchalash yoki ularni eruvchan shaklga aylantirishga qodir. Bu ularni tanadan tezroq tashishni osonlashtiradi. Qon va boshqa tana suyuqliklarida mavjud bo'lgan oqsillar ham patogen hujumga, ham shikastlanishga javob berish orqali immunitetni himoya qiladi. Immunoglobulinlar (antikorlar va komplement tizimining tarkibiy qismlari) bakteriyalarni, begona oqsillarni va viruslarni zararsizlantirishga qodir.
Tartibga solish mexanizmi
Na energiya manbai, na qurilish materiali vazifasini o'tamaydigan oqsil molekulalari ko'plab hujayra ichidagi jarayonlarni boshqaradi. Shunday qilib, ular tufayli translatsiya, transkripsiya, kesish va boshqa polipeptidlarning faolligi tartibga solinadi. Tartibga solish mexanizmi fermentativ faollikka asoslangan yoki boshqa molekulalar bilan o'ziga xos bog'lanish tufayli o'zini namoyon qiladi. Masalan, transkripsiya omillari, faollashtiruvchi polipeptidlar va repressor oqsillar gen transkripsiyasining intensivligini boshqarishga qodir. Bunda ular genlarni tartibga soluvchi ketma-ketliklar bilan o'zaro ta'sir qiladi. Hujayra ichidagi jarayonlarning borishini nazorat qilishda eng muhim rol oqsil fosfatazalari va oqsil kinazalariga beriladi. Bu fermentlar boshqa oqsillarga fosfat guruhlarini qo'shish yoki olib tashlash orqali ularning faolligini qo'zg'atadi yoki inhibe qiladi.

Signal vazifasi
Ko'pincha tartibga solish funktsiyasi bilan birlashtiriladi. Buning sababi shundaki, ko'plab hujayra ichidagi, shuningdek hujayradan tashqari polipeptidlar signallarni uzatishi mumkin. O'sish omillari, sitokinlar, gormonlar va boshqa birikmalar bu qobiliyatga ega. Steroidlar qon orqali tashiladi. Gormonning retseptor bilan o'zaro ta'siri hujayra reaktsiyasini qo'zg'atuvchi signal sifatida ishlaydi. Steroidlar qon va hujayralardagi birikmalar tarkibini, ko'payish, o'sish va boshqa jarayonlarni nazorat qiladi. Masalan, insulin. U glyukoza darajasini tartibga soladi. Hujayralarning o'zaro ta'siri hujayralararo modda orqali uzatiladigan signalli oqsil birikmalari orqali amalga oshiriladi.
Elementlarni tashish
Kichik molekulalarning harakatida ishtirok etadigan eruvchan oqsillar yuqori konsentratsiyada mavjud bo'lgan substratga yuqori yaqinlikka ega. Ular, shuningdek, uning tarkibi past bo'lgan joylarda osongina ozod qilish qobiliyatiga ega. Bunga misol sifatida transport oqsili gemoglobinni keltirish mumkin. U kislorodni o'pkadan boshqa to'qimalarga o'tkazadi va ulardan karbonat angidridni o'tkazadi. Ba'zi membrana oqsillari hujayra devorlari orqali kichik molekulalarni tashishda, ularni o'zgartirishda ham ishtirok etadi. Sitoplazmaning lipid qatlami suv o'tkazmaydigan. Bu zaryadlangan yoki qutbli molekulalarning tarqalishini oldini oladi. Membranli transport aloqalari odatda tashuvchilar va kanallarga bo'linadi.
Zaxira ulanishlar
Bu oqsillar zahira deb ataladigan narsalarni hosil qiladi. Ular, masalan, o'simlik urug'lari va hayvonlarning tuxumlarida to'planadi. Bunday oqsillar modda va energiyaning zahiraviy manbai sifatida ishlaydi. Ba'zi birikmalar organizm tomonidan aminokislotalar rezervuari sifatida ishlatiladi. Ular, o'z navbatida, metabolizmni tartibga solishda ishtirok etadigan faol moddalarning prekursorlari hisoblanadi.
Hujayra retseptorlari
Bunday oqsillar to'g'ridan-to'g'ri sitoplazmada yoki devorga ko'milgan bo'lishi mumkin. Ulanishning bir qismi signalni qabul qiladi. Qoida tariqasida, bu kimyoviy moddadir va ba'zi hollarda mexanik ta'sir (masalan, cho'zish), yorug'lik va boshqa ogohlantirishlar. Signalning molekulaning ma'lum bir qismiga - polipeptid retseptoriga ta'sir qilish jarayonida uning konformatsion o'zgarishlari boshlanadi. Ular qo'zg'atuvchini hujayraning boshqa tarkibiy qismlariga o'tkazadigan qolgan qismning konformatsiyasining o'zgarishiga olib keladi. Signal yuborish turli yo'llar bilan amalga oshirilishi mumkin. Ba'zi retseptorlar kimyoviy reaktsiyani katalizlash qobiliyatiga ega, boshqalari esa stimul ta'sirida yopiladigan yoki ochiladigan ion kanallari vazifasini bajaradi. Ba'zi birikmalar hujayra ichidagi messenjer molekulalarni maxsus bog'laydi.
Motor polipeptidlari
Tananing harakatini ta'minlaydigan oqsillarning butun sinfi mavjud. Dvigatel oqsillari mushaklarning qisqarishida, hujayralar harakatida, flagella va siliya faoliyatida ishtirok etadi. Shuningdek, ular yo'nalishli va faol transportni ta'minlaydi. Kinesinlar va dininlar energiya manbai sifatida ATP gidrolizidan foydalangan holda mikrotubulalar bo'ylab molekulalarni tashiydilar. Ikkinchisi organellalar va boshqa elementlarni periferik hujayra sohalaridan sentrozoma tomon siljitadi. Kinesinlar teskari yo'nalishda harakat qiladi. Dyneinlar flagella va siliya faoliyati uchun ham javobgardir.
 l l Oqsillarning strukturaviy tashkil etilishini yoritish zamonaviy biokimyoning asosiy muammolaridan biridir.Oqsillarning funksiyalarining juda xilma-xilligini tushunish uchun muhim ilmiy va amaliy ahamiyatga ega.Oqsil molekulalari 20 xil aminokislotalarning polikondensatsiyasi mahsuloti bo‘lib, ular bilan bog‘lanmagan. xaotik, lekin oqsil sintezi kodiga qat'iy muvofiq.Oqsil molekulasida necha o'nlab va yuzlab aminokislotalar bir-biri bilan qanday bog'langan?
l l Oqsillarning strukturaviy tashkil etilishini yoritish zamonaviy biokimyoning asosiy muammolaridan biridir.Oqsillarning funksiyalarining juda xilma-xilligini tushunish uchun muhim ilmiy va amaliy ahamiyatga ega.Oqsil molekulalari 20 xil aminokislotalarning polikondensatsiyasi mahsuloti bo‘lib, ular bilan bog‘lanmagan. xaotik, lekin oqsil sintezi kodiga qat'iy muvofiq.Oqsil molekulasida necha o'nlab va yuzlab aminokislotalar bir-biri bilan qanday bog'langan?
 Protein tuzilishining polipeptid nazariyasi l l 1902 yil, Emil Fisher Proteinlar murakkab polipeptidlar bo'lib, ularda alohida aminokislotalar bir-biri bilan aminokislotalarning karboksil va aminokislotalarning o'zaro ta'siridan kelib chiqadigan peptid (R-CO-NH-R) bog'lari bilan bog'langan.
Protein tuzilishining polipeptid nazariyasi l l 1902 yil, Emil Fisher Proteinlar murakkab polipeptidlar bo'lib, ularda alohida aminokislotalar bir-biri bilan aminokislotalarning karboksil va aminokislotalarning o'zaro ta'siridan kelib chiqadigan peptid (R-CO-NH-R) bog'lari bilan bog'langan.
 Nazariyaning eksperimental isboti l l l Tabiiy oqsillarda nisbatan kam erkin guruhlar mavjud –COOH va –NH 2 Oqsil gidroliz jarayonida stexiometrik miqdorda titrlanadigan guruhlar – COOH va –NH 2 hosil bo‘ladi.Proteolitik fermentlar ta’sirida, oqsillar qat'iy belgilangan bo'laklarga (polipeptidlar) bo'linadi terminal aminokislotalar ferment ta'sirining mos keluvchi selektivligi Oqsillar biuret reaktsiyasini beradi Rentgen nurlari difraksiya naqshlarini tahlil qilish oqsilning polipeptid tuzilishini tasdiqlaydi Oqsil tuzilishining polipeptid nazariyasining muhim tasdig'i imkoniyatdir. allaqachon ma'lum tuzilishga ega bo'lgan (insulin, lizozim, ribonukleaza va boshqalar) polipeptidlar va oqsillarni kimyoviy usullar bilan sintez qilish.
Nazariyaning eksperimental isboti l l l Tabiiy oqsillarda nisbatan kam erkin guruhlar mavjud –COOH va –NH 2 Oqsil gidroliz jarayonida stexiometrik miqdorda titrlanadigan guruhlar – COOH va –NH 2 hosil bo‘ladi.Proteolitik fermentlar ta’sirida, oqsillar qat'iy belgilangan bo'laklarga (polipeptidlar) bo'linadi terminal aminokislotalar ferment ta'sirining mos keluvchi selektivligi Oqsillar biuret reaktsiyasini beradi Rentgen nurlari difraksiya naqshlarini tahlil qilish oqsilning polipeptid tuzilishini tasdiqlaydi Oqsil tuzilishining polipeptid nazariyasining muhim tasdig'i imkoniyatdir. allaqachon ma'lum tuzilishga ega bo'lgan (insulin, lizozim, ribonukleaza va boshqalar) polipeptidlar va oqsillarni kimyoviy usullar bilan sintez qilish.
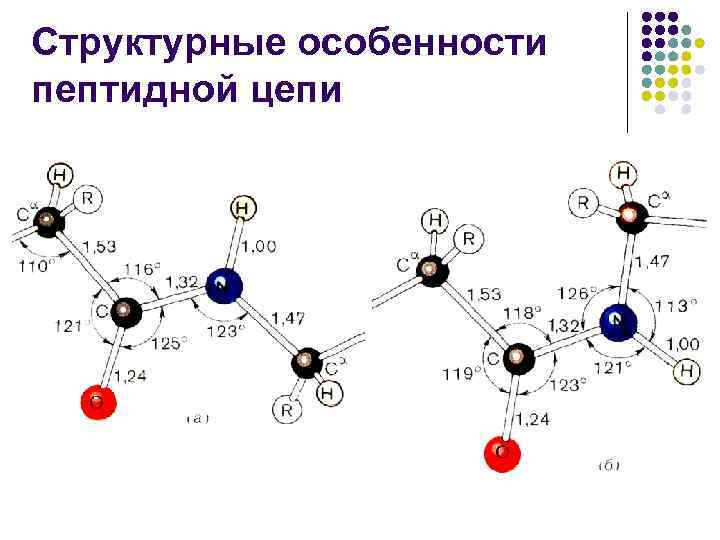
 Peptid zanjirining strukturaviy xususiyatlari l l Peptid bog'i -C-N- bog'idan taxminan 10% qisqaroq va "qisman qo'sh" bog'lanish xarakteriga ega -C=N- L. Pauling va R. Kori 1948 yilda taklif qilgan. 1955 yil. C-N aloqasining o'ziga xos xususiyatini ikkita shakl orasidagi "rezonans" bilan tushuntiring
Peptid zanjirining strukturaviy xususiyatlari l l Peptid bog'i -C-N- bog'idan taxminan 10% qisqaroq va "qisman qo'sh" bog'lanish xarakteriga ega -C=N- L. Pauling va R. Kori 1948 yilda taklif qilgan. 1955 yil. C-N aloqasining o'ziga xos xususiyatini ikkita shakl orasidagi "rezonans" bilan tushuntiring
 Peptid zanjirining strukturaviy xususiyatlari l l l Boshqacha qilib aytganda, oqsillar va peptidlarda azot atomining yolg'iz elektron juftligining karbonil guruhi elektron tizimi bilan o'zaro ta'siri tufayli C-N bog'lanish qisman ko'p bo'lib, bu qiyinlikka olib keladi. Odatda peptid bog'i konfiguratsiyaga ega bo'lib, sis konfiguratsiyasiga qaraganda 2,6 kkal/mol (10,878 kJ/mol) qulayroqdir, chunki sis konfiguratsiyasida -uglerod atomlarining birikishi tufayli. fazoviy qiyinchiliklarni keltirib chiqaradi.Agar peptid bog'ining azoti taxminan ga tegishli bo'lsa, bu farq katta darajada yo'qoladi.
Peptid zanjirining strukturaviy xususiyatlari l l l Boshqacha qilib aytganda, oqsillar va peptidlarda azot atomining yolg'iz elektron juftligining karbonil guruhi elektron tizimi bilan o'zaro ta'siri tufayli C-N bog'lanish qisman ko'p bo'lib, bu qiyinlikka olib keladi. Odatda peptid bog'i konfiguratsiyaga ega bo'lib, sis konfiguratsiyasiga qaraganda 2,6 kkal/mol (10,878 kJ/mol) qulayroqdir, chunki sis konfiguratsiyasida -uglerod atomlarining birikishi tufayli. fazoviy qiyinchiliklarni keltirib chiqaradi.Agar peptid bog'ining azoti taxminan ga tegishli bo'lsa, bu farq katta darajada yo'qoladi.
 Peptid zanjirining strukturaviy xususiyatlari l l l Asimmetrik atomga tutashgan ikkita oddiy bog'lanish (N–C va C–C 1) atrofida aylanishlar mumkin. Bunday aylanishlar dihedral burchaklar bilan o'lchanadi va peptid zanjirining har qanday konformatsiyasi bir qator bilan tavsiflanishi mumkin. burchak qiymatlari va C atomlarining har birida (odatda = 180 - trans shakli)
Peptid zanjirining strukturaviy xususiyatlari l l l Asimmetrik atomga tutashgan ikkita oddiy bog'lanish (N–C va C–C 1) atrofida aylanishlar mumkin. Bunday aylanishlar dihedral burchaklar bilan o'lchanadi va peptid zanjirining har qanday konformatsiyasi bir qator bilan tavsiflanishi mumkin. burchak qiymatlari va C atomlarining har birida (odatda = 180 - trans shakli)
 Peptid zanjirining strukturaviy xususiyatlari l l Peptid zanjiridagi o'rinbosarlarning o'zaro ta'siri tufayli burchaklar va hech qanday qiymatlarni qabul qila olmaydi - ular uchun faqat peptid zanjirining energiya jihatdan qulay konformatsiyalariga mos keladigan ba'zi diskret hududlarga ruxsat beriladi. burchaklar va bir-biriga bog'langan, ulardan birining o'zgarishi ikkinchi Gopalachamudram Narayana Ramachandranning o'zgarishiga olib keladi.
Peptid zanjirining strukturaviy xususiyatlari l l Peptid zanjiridagi o'rinbosarlarning o'zaro ta'siri tufayli burchaklar va hech qanday qiymatlarni qabul qila olmaydi - ular uchun faqat peptid zanjirining energiya jihatdan qulay konformatsiyalariga mos keladigan ba'zi diskret hududlarga ruxsat beriladi. burchaklar va bir-biriga bog'langan, ulardan birining o'zgarishi ikkinchi Gopalachamudram Narayana Ramachandranning o'zgarishiga olib keladi.

 l K.V.Linderström-Lang oqsil molekulalarining tashkil etilishining 4 darajasini - birlamchi, ikkilamchi, uchinchi darajali va to'rtlamchi tuzilmalarni ajratishni taklif qildi. Ushbu toifalar biroz eskirgan bo'lsa-da, ulardan foydalanish davom etmoqda
l K.V.Linderström-Lang oqsil molekulalarining tashkil etilishining 4 darajasini - birlamchi, ikkilamchi, uchinchi darajali va to'rtlamchi tuzilmalarni ajratishni taklif qildi. Ushbu toifalar biroz eskirgan bo'lsa-da, ulardan foydalanish davom etmoqda

 Proteinning birlamchi tuzilishi l l l polipeptid zanjiridagi aminokislotalar qoldiqlarining ketma-ketligi ma'lum bir oqsilning strukturaviy geni tomonidan kodlangan va uning fazoviy tuzilishini o'z-o'zini tashkil qilish uchun zarur bo'lgan hamma narsani o'z ichiga oladi Barcha oqsillar birlamchi tuzilishida farqlanadi. Bunday tuzilmalarning soni cheksizdir, ammo tirik tabiatda bu imkoniyatlarning barchasi amalga oshirilmaydi: umuman olganda, barcha turdagi tirik organizmlarda har xil turdagi oqsillar soni 1010-1012 ga teng.
Proteinning birlamchi tuzilishi l l l polipeptid zanjiridagi aminokislotalar qoldiqlarining ketma-ketligi ma'lum bir oqsilning strukturaviy geni tomonidan kodlangan va uning fazoviy tuzilishini o'z-o'zini tashkil qilish uchun zarur bo'lgan hamma narsani o'z ichiga oladi Barcha oqsillar birlamchi tuzilishida farqlanadi. Bunday tuzilmalarning soni cheksizdir, ammo tirik tabiatda bu imkoniyatlarning barchasi amalga oshirilmaydi: umuman olganda, barcha turdagi tirik organizmlarda har xil turdagi oqsillar soni 1010-1012 ga teng.
 l l l Oqsilning biologik funksiyasini, xususan, fiziologik ta’sirining molekulyar mexanizmini uning tuzilishini batafsil bilmay turib bilish mumkin emas.Oqsilning birlamchi strukturasini aniqlash ikkilamchi va uchinchi darajali tuzilmalarini aniqlashning asosidir. uning faol markazida funktsional guruhlarning joylashishi, uning ishlash mexanizmini tushunish yo'li. “Mutagen” oqsillarning birlamchi tuzilishini o‘rganish irsiy kasalliklarning mohiyatini molekulyar darajada aniqlash imkonini beradi.Birlamchi tuzilish haqidagi ma’lumotlar turli tipdagi tirik organizmlar o‘rtasidagi taksonomik aloqalarni o‘rnatish va tekshirishda ko‘rsatkichlardan biri sifatida foydalaniladi. va biologik evolyutsiya sxemasini qurish
l l l Oqsilning biologik funksiyasini, xususan, fiziologik ta’sirining molekulyar mexanizmini uning tuzilishini batafsil bilmay turib bilish mumkin emas.Oqsilning birlamchi strukturasini aniqlash ikkilamchi va uchinchi darajali tuzilmalarini aniqlashning asosidir. uning faol markazida funktsional guruhlarning joylashishi, uning ishlash mexanizmini tushunish yo'li. “Mutagen” oqsillarning birlamchi tuzilishini o‘rganish irsiy kasalliklarning mohiyatini molekulyar darajada aniqlash imkonini beradi.Birlamchi tuzilish haqidagi ma’lumotlar turli tipdagi tirik organizmlar o‘rtasidagi taksonomik aloqalarni o‘rnatish va tekshirishda ko‘rsatkichlardan biri sifatida foydalaniladi. va biologik evolyutsiya sxemasini qurish
 Birlamchi tuzilmani aniqlash usullari l l Birlamchi strukturani tahlil qilish uchun oqsilni tayyorlash uning tashkil etilishining boshqa, yuqori darajadagi ta'sirini minimallashtirish uchun mo'ljallangan.Boshqacha aytganda, tahlil ob'ekti o'zaro bog'langan kovalent bog'larsiz tartibsiz oqsil zanjiri bo'lishi kerak ( masalan, disulfid bog'lari) shuning uchun uning barcha birliklari, barcha peptid bog'lari kimyoviy reagentlar va fermentlarning ta'siriga teng darajada kirish mumkin edi.
Birlamchi tuzilmani aniqlash usullari l l Birlamchi strukturani tahlil qilish uchun oqsilni tayyorlash uning tashkil etilishining boshqa, yuqori darajadagi ta'sirini minimallashtirish uchun mo'ljallangan.Boshqacha aytganda, tahlil ob'ekti o'zaro bog'langan kovalent bog'larsiz tartibsiz oqsil zanjiri bo'lishi kerak ( masalan, disulfid bog'lari) shuning uchun uning barcha birliklari, barcha peptid bog'lari kimyoviy reagentlar va fermentlarning ta'siriga teng darajada kirish mumkin edi.
 l l Oqsil birinchi navbatda chuqur denaturatsiyaga duchor bo'lishi va to'rtlamchi, uchlamchi va iloji bo'lsa, ikkilamchi tuzilmalarini yo'qotishi kerak.Agar tarkibida disulfid bog'lari bo'lsa, ular odatda merkapto birikmasining ko'p miqdorda qaytarilishi yordamida parchalanadi.
l l Oqsil birinchi navbatda chuqur denaturatsiyaga duchor bo'lishi va to'rtlamchi, uchlamchi va iloji bo'lsa, ikkilamchi tuzilmalarini yo'qotishi kerak.Agar tarkibida disulfid bog'lari bo'lsa, ular odatda merkapto birikmasining ko'p miqdorda qaytarilishi yordamida parchalanadi.
 l Disulfid aloqalarining yopilishiga yo'l qo'ymaslik uchun sulfidril guruhlari yana ortiqcha yodoasetik kislota bilan bloklanadi: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI.
l Disulfid aloqalarining yopilishiga yo'l qo'ymaslik uchun sulfidril guruhlari yana ortiqcha yodoasetik kislota bilan bloklanadi: R–SH + I–CH 2–COOH R–S–CH 2–COOH + HI.
 Denatüratsiyalangan oqsil keyinchalik bir-birining ustiga chiqadigan peptidlar tizimini hosil qilish uchun tanlab gidrolizlanadi. Gidroliz fermentlar tomonidan amalga oshiriladi: l l l tripsin (arg yoki lysning karboksil guruhlari tomonidan hosil bo'lgan bog'lar), ximotripsin (aromatik aminokislotalarning –COOH guruhlari tomonidan hosil qilingan bog'lar) va boshqalar; kimyoviy reagentlar: siyanogen bromid Br -S N (-COOH met guruhlari tomonidan hosil qilingan aloqalar) va boshqalar.
Denatüratsiyalangan oqsil keyinchalik bir-birining ustiga chiqadigan peptidlar tizimini hosil qilish uchun tanlab gidrolizlanadi. Gidroliz fermentlar tomonidan amalga oshiriladi: l l l tripsin (arg yoki lysning karboksil guruhlari tomonidan hosil bo'lgan bog'lar), ximotripsin (aromatik aminokislotalarning –COOH guruhlari tomonidan hosil qilingan bog'lar) va boshqalar; kimyoviy reagentlar: siyanogen bromid Br -S N (-COOH met guruhlari tomonidan hosil qilingan aloqalar) va boshqalar.
 l l l Keyingi bosqich - hosil bo'lgan peptidlarni fraksiyalash - xromatografiya usullari bilan amalga oshiriladi.Nihoyat, oxirgi bosqich - birlamchi strukturani aniqlash l uning molekulyar massasini, aminokislotalar tarkibini, N- va terminal aminokislotalar qoldiqlarini aniqlashdan boshlanadi.Keyin. barcha fragmentlarning tuzilishi aniqlangan, oqsil boshqa agent yordamida parchalanadi va birinchisidan farqli ikkinchi peptid bo'laklari to'plami olinadi, ular oqsil tarkibidagi aminokislotalarning tartibini aniqlash uchun ajratiladi va tahlil qilinadi.
l l l Keyingi bosqich - hosil bo'lgan peptidlarni fraksiyalash - xromatografiya usullari bilan amalga oshiriladi.Nihoyat, oxirgi bosqich - birlamchi strukturani aniqlash l uning molekulyar massasini, aminokislotalar tarkibini, N- va terminal aminokislotalar qoldiqlarini aniqlashdan boshlanadi.Keyin. barcha fragmentlarning tuzilishi aniqlangan, oqsil boshqa agent yordamida parchalanadi va birinchisidan farqli ikkinchi peptid bo'laklari to'plami olinadi, ular oqsil tarkibidagi aminokislotalarning tartibini aniqlash uchun ajratiladi va tahlil qilinadi.

 F.Senger usuli l 1945-yilda ingliz biokimyogari F.Senger N-terminal aminokislotalarni aniqlashning birinchi usullaridan birini taklif qildi.
F.Senger usuli l 1945-yilda ingliz biokimyogari F.Senger N-terminal aminokislotalarni aniqlashning birinchi usullaridan birini taklif qildi.
 F. Sanger usuli l Protein (peptid) eritmasi 2, 4 dinitroflorobenzol (DNF) bilan ishlanadi, u erkin aminokislota bilan dinitrofenil (DNP) hosilasi beradi, sariq rangga ega:
F. Sanger usuli l Protein (peptid) eritmasi 2, 4 dinitroflorobenzol (DNF) bilan ishlanadi, u erkin aminokislota bilan dinitrofenil (DNP) hosilasi beradi, sariq rangga ega:
 F.Senger usuli l l Keyingi kislota gidrolizi (5,7 n HCl) peptid bog’larining parchalanishiga va N-terminal aminokislotalarning DNP hosilasi hosil bo’lishiga olib keladi.Oxirgi efir bilan ekstraksiya qilinadi va standartlar mavjud bo’lganda TLC tomonidan aniqlanadi.
F.Senger usuli l l Keyingi kislota gidrolizi (5,7 n HCl) peptid bog’larining parchalanishiga va N-terminal aminokislotalarning DNP hosilasi hosil bo’lishiga olib keladi.Oxirgi efir bilan ekstraksiya qilinadi va standartlar mavjud bo’lganda TLC tomonidan aniqlanadi.
 Dansil usuli l 1963 yilda ingliz biokimyogarlari V. Grey va B. Xartli tomonidan ishlab chiqilgan l Shuningdek, oqsilning aminokislotalariga "teg" ni kiritish asosida, keyingi gidrolizda Brayan Xartli olib tashlanmaydi.
Dansil usuli l 1963 yilda ingliz biokimyogarlari V. Grey va B. Xartli tomonidan ishlab chiqilgan l Shuningdek, oqsilning aminokislotalariga "teg" ni kiritish asosida, keyingi gidrolizda Brayan Xartli olib tashlanmaydi.
 Dansil usuli l Birinchi bosqich - dansilxloridning (1-dimetilaminoftalin-5-sulfoxlorid) peptidning (yoki oqsilning) protonlanmagan amino guruhi bilan dansil peptidini (DNS peptid) hosil qilish reaksiyasi.
Dansil usuli l Birinchi bosqich - dansilxloridning (1-dimetilaminoftalin-5-sulfoxlorid) peptidning (yoki oqsilning) protonlanmagan amino guruhi bilan dansil peptidini (DNS peptid) hosil qilish reaksiyasi.

 Dansil usuli l Olingan DNS oqsili 5,7 n ishtirokida gidrolizlanadi. HCl, 105 C da 12-16 soat davomida parchalangan DNS aminokislotasi UV nurlarida floresans bilan aniqlanadi.
Dansil usuli l Olingan DNS oqsili 5,7 n ishtirokida gidrolizlanadi. HCl, 105 C da 12-16 soat davomida parchalangan DNS aminokislotasi UV nurlarida floresans bilan aniqlanadi.


 S. Akabori usuli l l Peptid (yoki oqsil) suvsiz gidrazin bilan 100-120 S da qizdirilganda peptid bog lari gidrolizlanib aminokislota gidrazidlarini hosil qiladi, C-uchun aminokislota erkin aminokislota sifatida qoladi va uni ajratib olish mumkin. aralashmasidan va aniqlangan:
S. Akabori usuli l l Peptid (yoki oqsil) suvsiz gidrazin bilan 100-120 S da qizdirilganda peptid bog lari gidrolizlanib aminokislota gidrazidlarini hosil qiladi, C-uchun aminokislota erkin aminokislota sifatida qoladi va uni ajratib olish mumkin. aralashmasidan va aniqlangan:

 P.Edman usuli l fenilizotiosiyanat (FITC) yordamida polipeptid zanjirini parchalash usuli 1950–56 yillarda shved kimyogari P. V. Edman tomonidan ishlab chiqilgan. l Edman usuli N-terminal aminokislotalar qoldiqlarini feniltiohidantoinlar (PTH) shaklida ketma-ket yo'q qilishga imkon beradi l Har bir parchalanish sikli 3 bosqichni o'z ichiga oladi.
P.Edman usuli l fenilizotiosiyanat (FITC) yordamida polipeptid zanjirini parchalash usuli 1950–56 yillarda shved kimyogari P. V. Edman tomonidan ishlab chiqilgan. l Edman usuli N-terminal aminokislotalar qoldiqlarini feniltiohidantoinlar (PTH) shaklida ketma-ket yo'q qilishga imkon beradi l Har bir parchalanish sikli 3 bosqichni o'z ichiga oladi.




 P.Edman usuli l l l ajralgan PTG ni aniqlash Edman usulida hal qiluvchi ahamiyatga ega.Uzoq vaqt davomida qog’oz xromatografiyasi, so’ngra silikagel va poliamidda mikro yupqa qatlamli xromatografiya, suyuq va gaz-suyuqlik xromatografiyasi qo’llanilgan.Yurik yutuq. oqsillarni strukturaviy tadqiq qilish sohasi 1967 yilda P. Edman va J. Begg sekvenser (inglizcha ketma-ketlikdan - ketma-ketlik) - yuqori samaradorlik bilan N-terminal aminokislota qoldiqlarini ketma-ket avtomatik ravishda parchalashni amalga oshiradigan qurilmani yaratish edi. Edman usuli
P.Edman usuli l l l ajralgan PTG ni aniqlash Edman usulida hal qiluvchi ahamiyatga ega.Uzoq vaqt davomida qog’oz xromatografiyasi, so’ngra silikagel va poliamidda mikro yupqa qatlamli xromatografiya, suyuq va gaz-suyuqlik xromatografiyasi qo’llanilgan.Yurik yutuq. oqsillarni strukturaviy tadqiq qilish sohasi 1967 yilda P. Edman va J. Begg sekvenser (inglizcha ketma-ketlikdan - ketma-ketlik) - yuqori samaradorlik bilan N-terminal aminokislota qoldiqlarini ketma-ket avtomatik ravishda parchalashni amalga oshiradigan qurilmani yaratish edi. Edman usuli
 l Peptidlar va oqsillarning tuzilishini aniqlash uchun quyidagilardan foydalanishingiz mumkin: l l l N- va C-terminal aminokislotalar qoldiqlarining parchalanishini katalizlovchi fermentlar - aminokislotalar va karboksipeptidazalar; fizik-kimyoviy usullar, xususan, massa spektrometriyasi.
l Peptidlar va oqsillarning tuzilishini aniqlash uchun quyidagilardan foydalanishingiz mumkin: l l l N- va C-terminal aminokislotalar qoldiqlarining parchalanishini katalizlovchi fermentlar - aminokislotalar va karboksipeptidazalar; fizik-kimyoviy usullar, xususan, massa spektrometriyasi.
 Birlamchi struktura haqidagi ma'lumotlarni tahlil qilish quyidagi umumiy xulosalar chiqarish imkonini beradi l l Birlamchi strukturaning barqarorligi asosan peptid bog'lari bilan ta'minlanadi; oz sonli disulfid boglarning ishtirok etishi ham mumkin.Polipeptid zanjirida aminokislotalarning har xil birikmalarini topish mumkin.Barcha tasavvur qilinadigan dipeptidlar polipeptidlarda topilgan.Polipeptidlarda takrorlanuvchi ketma-ketliklar nisbatan kam uchraydi.
Birlamchi struktura haqidagi ma'lumotlarni tahlil qilish quyidagi umumiy xulosalar chiqarish imkonini beradi l l Birlamchi strukturaning barqarorligi asosan peptid bog'lari bilan ta'minlanadi; oz sonli disulfid boglarning ishtirok etishi ham mumkin.Polipeptid zanjirida aminokislotalarning har xil birikmalarini topish mumkin.Barcha tasavvur qilinadigan dipeptidlar polipeptidlarda topilgan.Polipeptidlarda takrorlanuvchi ketma-ketliklar nisbatan kam uchraydi.
 Birlamchi struktura haqidagi ma'lumotlarni tahlil qilish quyidagi umumiy xulosalar chiqarishga imkon beradi l l Har bir alohida bir hil oqsil o'ziga xos birlamchi tuzilish bilan tavsiflanadi; ko'pincha aminokislotalarning o'rnini bosish nafaqat tarkibiy o'zgarishlarga, balki fizik-kimyoviy xususiyatlar va biologik funktsiyalarning o'zgarishiga olib keladi.O'xshash xususiyatlarga ega bo'lgan ba'zi oqsillarda (ayniqsa fermentlarda) bir xil peptid tuzilmalari (aminokislotalar ketma-ketligi), ayniqsa, ularning joylashgan hududlarida topiladi. faol markazlar. Ushbu strukturaviy o'xshashlik printsipi bir qator proteolitik fermentlar - tripsin, ximotripsin va boshqalar uchun eng xosdir.
Birlamchi struktura haqidagi ma'lumotlarni tahlil qilish quyidagi umumiy xulosalar chiqarishga imkon beradi l l Har bir alohida bir hil oqsil o'ziga xos birlamchi tuzilish bilan tavsiflanadi; ko'pincha aminokislotalarning o'rnini bosish nafaqat tarkibiy o'zgarishlarga, balki fizik-kimyoviy xususiyatlar va biologik funktsiyalarning o'zgarishiga olib keladi.O'xshash xususiyatlarga ega bo'lgan ba'zi oqsillarda (ayniqsa fermentlarda) bir xil peptid tuzilmalari (aminokislotalar ketma-ketligi), ayniqsa, ularning joylashgan hududlarida topiladi. faol markazlar. Ushbu strukturaviy o'xshashlik printsipi bir qator proteolitik fermentlar - tripsin, ximotripsin va boshqalar uchun eng xosdir.

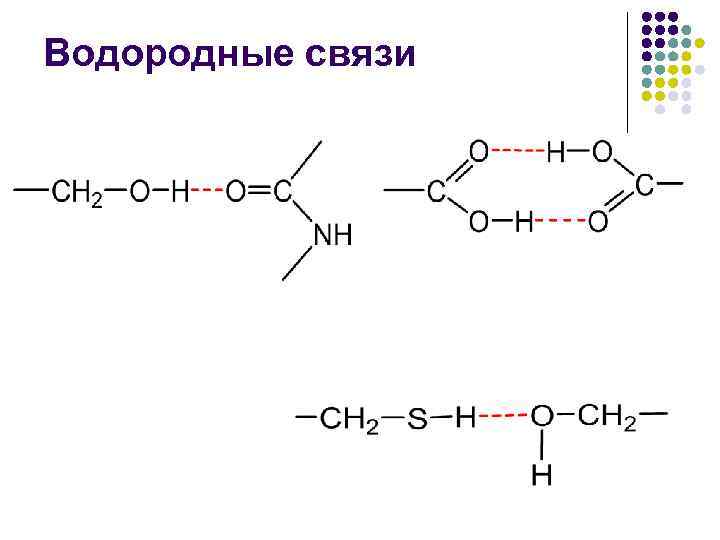
 Vodorod aloqalari Qutbsiz muhitda vodorod aloqasining energiyasi -CO NN- taxminan 16,7 kJ/mol ni tashkil qiladi va atrof-muhitning qutbliligini oshirish bu energiyani kamaytiradi.
Vodorod aloqalari Qutbsiz muhitda vodorod aloqasining energiyasi -CO NN- taxminan 16,7 kJ/mol ni tashkil qiladi va atrof-muhitning qutbliligini oshirish bu energiyani kamaytiradi.
 Gidrofobik o'zaro ta'sirlar l l Entropik xususiyatga ega.Ular qutbsiz o'rinbosarlarning suvdan siqib chiqarilishi va u bilan aloqasini cheklashga moyilligi bilan bog'liq.Aksincha, suv o'zining tuzilish holatini tiklashga intiladi va go'yo o'rnini bosuvchi moddalarni majburiy ravishda minimal energiyaga ega bo'lgan klasterlarga guruhlaydi.Asosan aminokislotalarning qutb bo'lmagan yon guruhlari qoldiqlarga kiradi.
Gidrofobik o'zaro ta'sirlar l l Entropik xususiyatga ega.Ular qutbsiz o'rinbosarlarning suvdan siqib chiqarilishi va u bilan aloqasini cheklashga moyilligi bilan bog'liq.Aksincha, suv o'zining tuzilish holatini tiklashga intiladi va go'yo o'rnini bosuvchi moddalarni majburiy ravishda minimal energiyaga ega bo'lgan klasterlarga guruhlaydi.Asosan aminokislotalarning qutb bo'lmagan yon guruhlari qoldiqlarga kiradi.

 Van der Vaals o'zaro ta'sirlari l l Atomlarning tortishish dispersiya kuchlari va ularning elektron qobiqlarining o'zaro itarilish kuchlaridan iborat Har bir kontaktning energiya hissasi kichik (
Van der Vaals o'zaro ta'sirlari l l Atomlarning tortishish dispersiya kuchlari va ularning elektron qobiqlarining o'zaro itarilish kuchlaridan iborat Har bir kontaktning energiya hissasi kichik (
 Ion yoki elektrostatik o'zaro ta'sirlar l Zaryadlangan guruhlarning o'zaro ta'siri l l Tuz bog'larini hosil qiluvchi ionogen guruhlarning o'zaro ta'siri Hidrofob muhitda tuz bog'larining energiyasi 41,9 kJ/mol ga yetishi mumkin, lekin ularning soni nisbatan kichik Muhitning dielektrik o'tkazuvchanligining oshishi. tuz aloqalarining energiyasini kamaytiradi ion-dipol va dipol-dipol o'zaro ta'siri ko'p jihatdan o'xshashdir
Ion yoki elektrostatik o'zaro ta'sirlar l Zaryadlangan guruhlarning o'zaro ta'siri l l Tuz bog'larini hosil qiluvchi ionogen guruhlarning o'zaro ta'siri Hidrofob muhitda tuz bog'larining energiyasi 41,9 kJ/mol ga yetishi mumkin, lekin ularning soni nisbatan kichik Muhitning dielektrik o'tkazuvchanligining oshishi. tuz aloqalarining energiyasini kamaytiradi ion-dipol va dipol-dipol o'zaro ta'siri ko'p jihatdan o'xshashdir
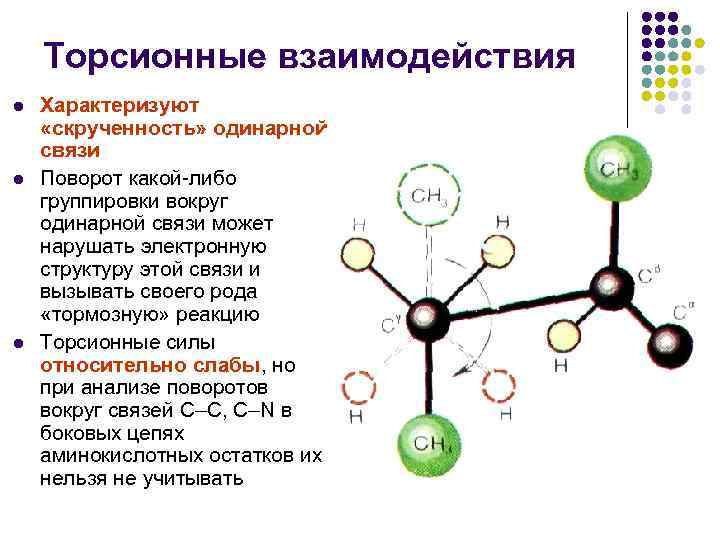 Buralish o'zaro ta'sirlari l l l Yagona bog'lanishning "burilishi" ni tavsiflang Har qanday guruhning bitta bog' atrofida aylanishi bu bog'lanishning elektron tuzilishini buzishi va o'ziga xos "tormoz" reaktsiyasini keltirib chiqarishi mumkin Buralish kuchlari nisbatan zaif, lekin C atrofida aylanishlarni tahlil qilganda - Aminokislotalar qoldiqlarining yon zanjirlaridagi C, C-N aloqalari ularni e'tiborsiz qoldirib bo'lmaydi
Buralish o'zaro ta'sirlari l l l Yagona bog'lanishning "burilishi" ni tavsiflang Har qanday guruhning bitta bog' atrofida aylanishi bu bog'lanishning elektron tuzilishini buzishi va o'ziga xos "tormoz" reaktsiyasini keltirib chiqarishi mumkin Buralish kuchlari nisbatan zaif, lekin C atrofida aylanishlarni tahlil qilganda - Aminokislotalar qoldiqlarining yon zanjirlaridagi C, C-N aloqalari ularni e'tiborsiz qoldirib bo'lmaydi

 Oqsilning ikkilamchi tuzilishi... l l aminokislota yon radikallarining turi va konformatsiyasini hisobga olmasdan, polipeptid zanjirining alohida bo‘limlarining fazoda joylashishi ham bir zanjirning, ham turli zanjirlarning peptid guruhlari o‘rtasidagi vodorod bog‘lari natijasida hosil bo‘ladi.
Oqsilning ikkilamchi tuzilishi... l l aminokislota yon radikallarining turi va konformatsiyasini hisobga olmasdan, polipeptid zanjirining alohida bo‘limlarining fazoda joylashishi ham bir zanjirning, ham turli zanjirlarning peptid guruhlari o‘rtasidagi vodorod bog‘lari natijasida hosil bo‘ladi.
 Oqsilning ikkilamchi tuzilishi l l l Oqsil molekulasining har qanday hududi ikkilamchi tuzilishga ega.Ba'zan faqat uning davriy elementlari ikkilamchi tuzilish sifatida qaraladi: -spiral va -struktura.Ammo oqsillarda juda o'ziga xos bir doirada joylashgan hududlar mavjud. yo'l, garchi ularning fazoviy tuzilishida davriy takrorlanadigan tuzilma bo'lmasa ham, muntazam motiv. Shunga qaramay, ikkilamchi tuzilish tushunchasi ular uchun anchagina qo'llaniladi.Ikkilamchi tuzilmalarning 2 turi mavjud: muntazam va tartibsiz.Ikkilamchi tuzilish tushunchasi butun oqsil molekulasiga emas, balki alohida ko'proq yoki kamroq kengaytirilgan bo'limlarga tegishli. uning polipeptid zanjiri
Oqsilning ikkilamchi tuzilishi l l l Oqsil molekulasining har qanday hududi ikkilamchi tuzilishga ega.Ba'zan faqat uning davriy elementlari ikkilamchi tuzilish sifatida qaraladi: -spiral va -struktura.Ammo oqsillarda juda o'ziga xos bir doirada joylashgan hududlar mavjud. yo'l, garchi ularning fazoviy tuzilishida davriy takrorlanadigan tuzilma bo'lmasa ham, muntazam motiv. Shunga qaramay, ikkilamchi tuzilish tushunchasi ular uchun anchagina qo'llaniladi.Ikkilamchi tuzilmalarning 2 turi mavjud: muntazam va tartibsiz.Ikkilamchi tuzilish tushunchasi butun oqsil molekulasiga emas, balki alohida ko'proq yoki kamroq kengaytirilgan bo'limlarga tegishli. uning polipeptid zanjiri
 Oqsilning ikkilamchi tuzilishi l Ikkilamchi strukturaning shakllanishida eng muhim rol o'ynaydigan o'zaro ta'sirlar quyidagilar bilan belgilanadi: l l l peptid bog'lanish xususiyatlari sterik cheklovlar (- va - burchaklar) Asosiy rolni peptidlar orasidagi vodorod bog'lari bajaradi. zanjirda vaqti-vaqti bilan takrorlanadigan guruhlar
Oqsilning ikkilamchi tuzilishi l Ikkilamchi strukturaning shakllanishida eng muhim rol o'ynaydigan o'zaro ta'sirlar quyidagilar bilan belgilanadi: l l l peptid bog'lanish xususiyatlari sterik cheklovlar (- va - burchaklar) Asosiy rolni peptidlar orasidagi vodorod bog'lari bajaradi. zanjirda vaqti-vaqti bilan takrorlanadigan guruhlar
 Vodorod aloqasi tufayli ikkilamchi strukturaning barqarorlashishi l l oqsilni o'rab turgan suvning ta'siri Suv vodorod bog'larini hosil qilish uchun samarali raqobatlasha oladi: Suvning ta'siri oqsilning ixcham fazoviy tuzilishi shakllanishi bilan kamayadi, tarkibining ko'payishi. peptid bog'lari va ularning o'zaro ta'sir qilish ehtimolini oshiradi.Demak, ikkilamchi strukturaning barqarorligi uning ixcham uchinchi tuzilishga kiritilishiga bog'liq.
Vodorod aloqasi tufayli ikkilamchi strukturaning barqarorlashishi l l oqsilni o'rab turgan suvning ta'siri Suv vodorod bog'larini hosil qilish uchun samarali raqobatlasha oladi: Suvning ta'siri oqsilning ixcham fazoviy tuzilishi shakllanishi bilan kamayadi, tarkibining ko'payishi. peptid bog'lari va ularning o'zaro ta'sir qilish ehtimolini oshiradi.Demak, ikkilamchi strukturaning barqarorligi uning ixcham uchinchi tuzilishga kiritilishiga bog'liq.

 -Spiral l XX asrning 50-yillarida. L. Pauling va R. Kori aminokislotalar va oddiy peptidlar kristallarining tuzilishi haqidagi ma'lumotlarga asoslanib, polipeptid zanjirining mumkin bo'lgan davriy konformatsiyasini o'rganib chiqdilar va eng ehtimolli struktura - bu - spiral deb atashgan degan xulosaga kelishdi.
-Spiral l XX asrning 50-yillarida. L. Pauling va R. Kori aminokislotalar va oddiy peptidlar kristallarining tuzilishi haqidagi ma'lumotlarga asoslanib, polipeptid zanjirining mumkin bo'lgan davriy konformatsiyasini o'rganib chiqdilar va eng ehtimolli struktura - bu - spiral deb atashgan degan xulosaga kelishdi.
 -Spiral Uni tanlash quyidagi mezonlarga asoslandi: 1. Bo'shliqlarsiz va bir-birining ustiga chiqadigan atomlarsiz yaqin o'ralgan ixcham strukturaning shakllanishi 2. Ularning geometriyasi chiziqli bo'lishi sharti bilan strukturaning vodorod bog'lari bilan maksimal to'yinganligi 3. Muvofiqlik. aminokislotalar va oddiy peptidlarga xos bo'lgan atomlararo masofalar va burchaklar l
-Spiral Uni tanlash quyidagi mezonlarga asoslandi: 1. Bo'shliqlarsiz va bir-birining ustiga chiqadigan atomlarsiz yaqin o'ralgan ixcham strukturaning shakllanishi 2. Ularning geometriyasi chiziqli bo'lishi sharti bilan strukturaning vodorod bog'lari bilan maksimal to'yinganligi 3. Muvofiqlik. aminokislotalar va oddiy peptidlarga xos bo'lgan atomlararo masofalar va burchaklar l
 -spiral l Ushbu shartlarga ko'ra, o'ng qo'l va chap qo'l spirallarini qurish mumkin, ammo peptid zanjiri L bilan hosil qilingan bo'lsa, o'ng qo'l spiral energiya jihatidan chapga qaraganda bir oz qulayroq bo'lib chiqadi. -aminokislotalar
-spiral l Ushbu shartlarga ko'ra, o'ng qo'l va chap qo'l spirallarini qurish mumkin, ammo peptid zanjiri L bilan hosil qilingan bo'lsa, o'ng qo'l spiral energiya jihatidan chapga qaraganda bir oz qulayroq bo'lib chiqadi. -aminokislotalar
 -Spiral l l Polipeptid zanjirining barcha bo'g'inlarida oddiy bog'lar (i) atrofida aylanish burchaklari bir xil kattalik va ishoraga ega bo'lganda (60 ga yaqin) spiral struktura paydo bo'ladi, bu esa zanjirning asta-sekin burilishiga olib keladi. aminokislotalar qoldiqlarining radikallari spiraldan hosil bo'lgan silindrning periferiyasida paydo bo'ladi va ularning tabiatiga qarab silindrsimon sirtning hidrofobik yoki hidrofil xususiyatini ta'minlashi mumkin.
-Spiral l l Polipeptid zanjirining barcha bo'g'inlarida oddiy bog'lar (i) atrofida aylanish burchaklari bir xil kattalik va ishoraga ega bo'lganda (60 ga yaqin) spiral struktura paydo bo'ladi, bu esa zanjirning asta-sekin burilishiga olib keladi. aminokislotalar qoldiqlarining radikallari spiraldan hosil bo'lgan silindrning periferiyasida paydo bo'ladi va ularning tabiatiga qarab silindrsimon sirtning hidrofobik yoki hidrofil xususiyatini ta'minlashi mumkin.

 -Spiral geometrik parametrlar: l l l l radius r = 2,3 Å (0,23 nm) spiral balandligi (o’zgartirish) 1 qoldiq uchun d = 1,5 Å spiral qadam (identifikatsiya davri) P = 5,4 Å 1 spiral burilish shakli 3 Å 6, barcha kislotalar – 3 , C=O bog'lari oldinga, C-uchiga, -N-H guruhlari esa orqaga yo'naltirilgan.Spiralda har bir -NH guruhi to'rtinchi aminokislotalar qoldig'ining -CO guruhiga vodorod bog'i orqali bog'langan. undan (5 1 obligatsiya)
-Spiral geometrik parametrlar: l l l l radius r = 2,3 Å (0,23 nm) spiral balandligi (o’zgartirish) 1 qoldiq uchun d = 1,5 Å spiral qadam (identifikatsiya davri) P = 5,4 Å 1 spiral burilish shakli 3 Å 6, barcha kislotalar – 3 , C=O bog'lari oldinga, C-uchiga, -N-H guruhlari esa orqaga yo'naltirilgan.Spiralda har bir -NH guruhi to'rtinchi aminokislotalar qoldig'ining -CO guruhiga vodorod bog'i orqali bog'langan. undan (5 1 obligatsiya)
 -Spiral l l Globulyar oqsillarda - spiral kesmalarining uzunligi nisbatan qisqa (5–15 aminokislota qoldiqlari, spiralning 3–4 burilishi), fibrilyar oqsillarda u ancha uzunroq. pro qoldiqlari kiritilgan joylarda vodorod bog'lanish tizimini to'xtatadi. Bunday holda, spiral o'qi 20-30 ga og'adi
-Spiral l l Globulyar oqsillarda - spiral kesmalarining uzunligi nisbatan qisqa (5–15 aminokislota qoldiqlari, spiralning 3–4 burilishi), fibrilyar oqsillarda u ancha uzunroq. pro qoldiqlari kiritilgan joylarda vodorod bog'lanish tizimini to'xtatadi. Bunday holda, spiral o'qi 20-30 ga og'adi

 -Tuzilish l l 1941 yilda V.T.Astberi tomonidan -keratinning rentgen diffraksion tadqiqotlari asosida taklif qilingan. O'n yil o'tgach, L.Pauling va R.Kori struktura yoki "buklangan varaq" cho'zilgan, zigzagning assotsiatsiyasi ekanligini aniqladilar. zanjirlararo vodorod aloqalari peptid zanjirlari bilan barqarorlashtirilgan tuzilmalar
-Tuzilish l l 1941 yilda V.T.Astberi tomonidan -keratinning rentgen diffraksion tadqiqotlari asosida taklif qilingan. O'n yil o'tgach, L.Pauling va R.Kori struktura yoki "buklangan varaq" cho'zilgan, zigzagning assotsiatsiyasi ekanligini aniqladilar. zanjirlararo vodorod aloqalari peptid zanjirlari bilan barqarorlashtirilgan tuzilmalar




 -Tuzilish l l -tuzilmani tashkil etuvchi peptid zanjiri segmentidagi aminokislotalar qoldiqlari soni, odatda 3 dan 8 gacha. Kengaytirilgan struktura, -qatlam yoki buklangan varaq deb ataladigan, ko'pincha 2-dan iborat. 6 ta zanjir, ba'zan 10 tagacha
-Tuzilish l l -tuzilmani tashkil etuvchi peptid zanjiri segmentidagi aminokislotalar qoldiqlari soni, odatda 3 dan 8 gacha. Kengaytirilgan struktura, -qatlam yoki buklangan varaq deb ataladigan, ko'pincha 2-dan iborat. 6 ta zanjir, ba'zan 10 tagacha
 -Tuzilish l l l Struktura hosil bo`lishi jarayonida uning sirtining turli tomonlarida aminokislotalar qoldiqlarining yon guruhlari paydo bo`ladi.Siraning o`zi burma shaklga ega bo`lib, burmalar -uglerod atomlari bilan belgilanadi.Ulardan cho`zilgan yon guruhlar hosil bo`ladi. tizmalar.Bu bir xil turdagi (masalan, hidrofobik) yon radikallar bilan toʻyingan ancha choʻzilgan yuzalarni hosil qilish imkonini beradi. oqsilning fazoviy tuzilishini barqarorlashtiradigan intramolekulyar hidrofobik yadrolar
-Tuzilish l l l Struktura hosil bo`lishi jarayonida uning sirtining turli tomonlarida aminokislotalar qoldiqlarining yon guruhlari paydo bo`ladi.Siraning o`zi burma shaklga ega bo`lib, burmalar -uglerod atomlari bilan belgilanadi.Ulardan cho`zilgan yon guruhlar hosil bo`ladi. tizmalar.Bu bir xil turdagi (masalan, hidrofobik) yon radikallar bilan toʻyingan ancha choʻzilgan yuzalarni hosil qilish imkonini beradi. oqsilning fazoviy tuzilishini barqarorlashtiradigan intramolekulyar hidrofobik yadrolar

 -Bending l l - spiral va - strukturasi odatda globulyar oqsillarda nisbatan qisqa bo'laklarda ifodalanadi, shuning uchun oqsilning ikkilamchi tuzilishining muhim qismi peptid yo'nalishini o'zgartirishga imkon beradigan turli xil ko'chadan iborat. zanjir.Eng tejamkor strukturaviy element, bu polipeptid 180ni atigi 3 ta peptid guruhidan foydalangan holda aylantirish imkonini beradi, – -bukish Bitta vodorod aloqasi bilan barqarorlashadi. boshqa molekulalar bilan o'zaro ta'siri
-Bending l l - spiral va - strukturasi odatda globulyar oqsillarda nisbatan qisqa bo'laklarda ifodalanadi, shuning uchun oqsilning ikkilamchi tuzilishining muhim qismi peptid yo'nalishini o'zgartirishga imkon beradigan turli xil ko'chadan iborat. zanjir.Eng tejamkor strukturaviy element, bu polipeptid 180ni atigi 3 ta peptid guruhidan foydalangan holda aylantirish imkonini beradi, – -bukish Bitta vodorod aloqasi bilan barqarorlashadi. boshqa molekulalar bilan o'zaro ta'siri


 l Rentgen nurlari diffraktsiyasi tahlili natijalariga ko'ra, oqsil molekulasining katlanma yo'li uning aminokislotalar ketma-ketligi bilan aniqlanishi aniqlandi l l spiral hosil bo'lishiga ala, ley, glu tuzilmalarining shakllanishi yordam beradi - met, val, ile, zanjirning egilishi - gly, pro, asn Agar 6 ta guruhlangan aminokislota qoldig'idan 4 tasi spiral hosil bo'lishiga yordam bersa, u holda bu guruh spirallanish markazi hisoblanadi.Agar 5 tadan 3 ta qoldiq guruhlangan bo'lsa. -tuzilmasining shakllanishiga hissa qo'shadi, keyin bu guruh -qatlam uchun urug'dir
l Rentgen nurlari diffraktsiyasi tahlili natijalariga ko'ra, oqsil molekulasining katlanma yo'li uning aminokislotalar ketma-ketligi bilan aniqlanishi aniqlandi l l spiral hosil bo'lishiga ala, ley, glu tuzilmalarining shakllanishi yordam beradi - met, val, ile, zanjirning egilishi - gly, pro, asn Agar 6 ta guruhlangan aminokislota qoldig'idan 4 tasi spiral hosil bo'lishiga yordam bersa, u holda bu guruh spirallanish markazi hisoblanadi.Agar 5 tadan 3 ta qoldiq guruhlangan bo'lsa. -tuzilmasining shakllanishiga hissa qo'shadi, keyin bu guruh -qatlam uchun urug'dir

 Oqsillarning supersekondar (ikkinchi darajali) tuzilishi l l Oqsil molekulasining tashkil etilishining bu darajasi bir-biri bilan o'zaro ta'sir qiluvchi ikkilamchi tuzilmalarning ansambllarining mavjudligi bilan bog'liq.Bunday ansambllarning fazoviy tuzilishi oqsil molekulasining o'ta ikkinchi darajali strukturasi deb ataladi, u energiya jihatidan. eng afzal
Oqsillarning supersekondar (ikkinchi darajali) tuzilishi l l Oqsil molekulasining tashkil etilishining bu darajasi bir-biri bilan o'zaro ta'sir qiluvchi ikkilamchi tuzilmalarning ansambllarining mavjudligi bilan bog'liq.Bunday ansambllarning fazoviy tuzilishi oqsil molekulasining o'ta ikkinchi darajali strukturasi deb ataladi, u energiya jihatidan. eng afzal

 O'ta o'ralgan - spiral l l Bir nechta spirallarni protofibrillalarga (-keratin) burish natijasida hosil bo'lgan, ular mikrofibrillalarga birlashadi - spirallarni van der Vaals kuchlari ushlab turadi.
O'ta o'ralgan - spiral l l Bir nechta spirallarni protofibrillalarga (-keratin) burish natijasida hosil bo'lgan, ular mikrofibrillalarga birlashadi - spirallarni van der Vaals kuchlari ushlab turadi.

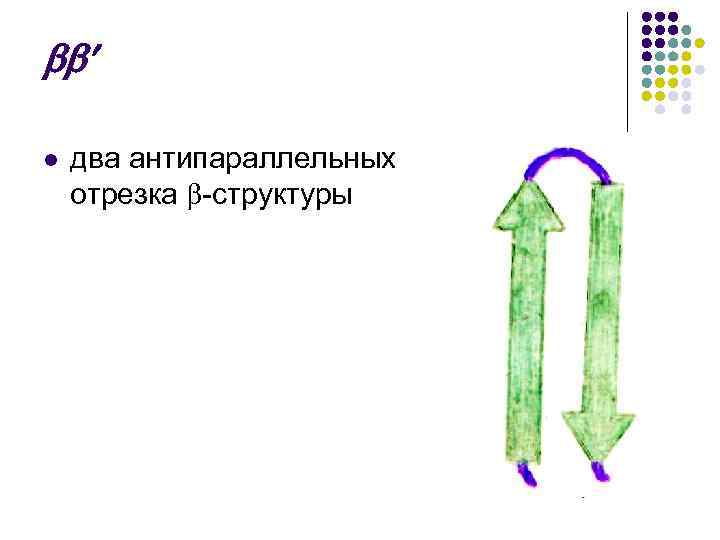
 x l ikkita parallel qatlamdan iborat bo'lgan bog'lanish, ular orasidagi tutashuv quyidagi shaklda: l tartibsiz g'altak – c
x l ikkita parallel qatlamdan iborat bo'lgan bog'lanish, ular orasidagi tutashuv quyidagi shaklda: l tartibsiz g'altak – c
 x l ikkita parallel qatlamdan iborat bo‘g‘inli bo‘g‘inli bo‘g‘in: l l -spiral – – Rossmanga ko‘ra zanjir yotqizish.Ikki qism ketma-ket bog‘langan “yunon ornamenti” strukturasini hosil qiladi –
x l ikkita parallel qatlamdan iborat bo‘g‘inli bo‘g‘inli bo‘g‘in: l l -spiral – – Rossmanga ko‘ra zanjir yotqizish.Ikki qism ketma-ket bog‘langan “yunon ornamenti” strukturasini hosil qiladi –
 x l bo'g'in ikki parallel -qatlamdan iborat bo'lib, ular orasidagi bo'g'in shaklida: l -tuzilma – – - zigzag, “yunoncha kalit”
x l bo'g'in ikki parallel -qatlamdan iborat bo'lib, ular orasidagi bo'g'in shaklida: l -tuzilma – – - zigzag, “yunoncha kalit”

 Domenlar l l Ko'pgina oqsillar nisbatan zaif o'zaro ta'sir qiluvchi mintaqalarni o'z ichiga oladi - domenlar Domenlar (fransuzcha domene - egalik) - molekulaning strukturaviy va funktsional jihatdan alohida hududlari (subregionlari), polipeptid zanjirining qisqa bo'limlari bilan bir-biriga bog'langan, ular menteşe mintaqalari deb ataladi. Funktsional domenlar bir yoki bir nechta strukturaviy domenlardan iborat bo'lishi mumkin.Bir qator fermentlarda faol markaz domenlar orasidagi chuqurchalarda joylashgan.
Domenlar l l Ko'pgina oqsillar nisbatan zaif o'zaro ta'sir qiluvchi mintaqalarni o'z ichiga oladi - domenlar Domenlar (fransuzcha domene - egalik) - molekulaning strukturaviy va funktsional jihatdan alohida hududlari (subregionlari), polipeptid zanjirining qisqa bo'limlari bilan bir-biriga bog'langan, ular menteşe mintaqalari deb ataladi. Funktsional domenlar bir yoki bir nechta strukturaviy domenlardan iborat bo'lishi mumkin.Bir qator fermentlarda faol markaz domenlar orasidagi chuqurchalarda joylashgan.



 3. + - Proteinlar l to'liq spirallardan tashkil topgan hududlarga va to'liq insulinga qarshi parallel qatlamlardan tashkil topgan hududlarga ega.
3. + - Proteinlar l to'liq spirallardan tashkil topgan hududlarga va to'liq insulinga qarshi parallel qatlamlardan tashkil topgan hududlarga ega.

 4. / -Oqsillar l l -Zanjir bo'ylab spiral va -tuzilmalar almashinadi.Ko'pchilik -tuzilmalar (parallel) molekulaning markaziy qismida lokalizatsiya qilingan bo'lib, bu tuzilmalar parvona kabi egilib (“burilish” strukturasi) hosil qiladi. molekulaning qolgan qismlari ulangan qattiq "tayanch"
4. / -Oqsillar l l -Zanjir bo'ylab spiral va -tuzilmalar almashinadi.Ko'pchilik -tuzilmalar (parallel) molekulaning markaziy qismida lokalizatsiya qilingan bo'lib, bu tuzilmalar parvona kabi egilib (“burilish” strukturasi) hosil qiladi. molekulaning qolgan qismlari ulangan qattiq "tayanch"


 Domenlar l l l Domenning o'rtacha hajmi odatda 100-150 qoldiq bo'lib, diametri taxminan 2,5 nm bo'lgan globulaga to'g'ri keladi.Shu bilan birga, ancha katta domenlar ham topiladi.Ehtimol, oqsilning fazoviy tuzilishining shakllanishi. birinchi navbatda kelajakdagi domenlar ichida sodir bo'ladi va domenlarning o'zaro stackingi, ya'ni uchinchi darajali strukturaning shakllanishi globula shakllanishining oxirgi bosqichida sodir bo'ladi.
Domenlar l l l Domenning o'rtacha hajmi odatda 100-150 qoldiq bo'lib, diametri taxminan 2,5 nm bo'lgan globulaga to'g'ri keladi.Shu bilan birga, ancha katta domenlar ham topiladi.Ehtimol, oqsilning fazoviy tuzilishining shakllanishi. birinchi navbatda kelajakdagi domenlar ichida sodir bo'ladi va domenlarning o'zaro stackingi, ya'ni uchinchi darajali strukturaning shakllanishi globula shakllanishining oxirgi bosqichida sodir bo'ladi.

 Oqsilning uchinchi darajali tuzilishi... l l l butun polipeptid zanjiridagi tartibli va amorf hududlarning fazoda joylashishini xarakterlaydi, bu yon radikallarning o'zaro ta'siri orqali erishiladi va ularning turi va konformatsiyasiga bog'liq.Shunday qilib, uchinchi darajali struktura tavsiflaydi. butun oqsil molekulasining fazoviy joylashuvi, agar u bitta polipeptid zanjiri tomonidan tashkil etilgan bo'lsa, u to'g'ridan-to'g'ri oqsil molekulalarining shakliga bog'liq bo'lib, ular har xil bo'lishi mumkin: sharsimondan ipga o'xshash.
Oqsilning uchinchi darajali tuzilishi... l l l butun polipeptid zanjiridagi tartibli va amorf hududlarning fazoda joylashishini xarakterlaydi, bu yon radikallarning o'zaro ta'siri orqali erishiladi va ularning turi va konformatsiyasiga bog'liq.Shunday qilib, uchinchi darajali struktura tavsiflaydi. butun oqsil molekulasining fazoviy joylashuvi, agar u bitta polipeptid zanjiri tomonidan tashkil etilgan bo'lsa, u to'g'ridan-to'g'ri oqsil molekulalarining shakliga bog'liq bo'lib, ular har xil bo'lishi mumkin: sharsimondan ipga o'xshash.
 Ipga o'xshash yoki fibrilyar oqsillar l l ipak fibroin keratin sochlar, shoxlar, tuyoqlar kollagen va boshqalar.
Ipga o'xshash yoki fibrilyar oqsillar l l ipak fibroin keratin sochlar, shoxlar, tuyoqlar kollagen va boshqalar.

 Protein l l ning uchinchi darajali tuzilishi oqsil funksionalligining asosi bo‘lib, u ko‘plab aminokislotalar qoldiqlaridan tuzilgan yirik ansambllarning fazoda aniq tashkil etilishini talab qiladi.Bunday ansambllar (domenlar) hosil bo‘ladi: l l fermentlarning faol markazlari, boshqa biologik molekulalarning bog‘lanish zonalari, oqsillarning effektor markazlari va boshqalar, shuning uchun uchinchi darajali protein tuzilishining buzilishi (denaturatsiya) doimo uning ishlash qobiliyatini yo'qotishiga olib keladi.
Protein l l ning uchinchi darajali tuzilishi oqsil funksionalligining asosi bo‘lib, u ko‘plab aminokislotalar qoldiqlaridan tuzilgan yirik ansambllarning fazoda aniq tashkil etilishini talab qiladi.Bunday ansambllar (domenlar) hosil bo‘ladi: l l fermentlarning faol markazlari, boshqa biologik molekulalarning bog‘lanish zonalari, oqsillarning effektor markazlari va boshqalar, shuning uchun uchinchi darajali protein tuzilishining buzilishi (denaturatsiya) doimo uning ishlash qobiliyatini yo'qotishiga olib keladi.
 Oqsilning uchinchi darajali tuzilishi l l l Uchinchi darajali strukturaning barqarorligi oqsil globulasi ichidagi kovalent bo'lmagan o'zaro ta'sirlar tizimiga bog'liq (?) Ba'zi oqsillar kovalent - disulfid - bog'lanishlar bilan qo'shimcha ravishda barqarorlashadi G zanjiri = RTln. K = Nchain - T Schain Nchain 0 (S
Oqsilning uchinchi darajali tuzilishi l l l Uchinchi darajali strukturaning barqarorligi oqsil globulasi ichidagi kovalent bo'lmagan o'zaro ta'sirlar tizimiga bog'liq (?) Ba'zi oqsillar kovalent - disulfid - bog'lanishlar bilan qo'shimcha ravishda barqarorlashadi G zanjiri = RTln. K = Nchain - T Schain Nchain 0 (S
 Oqsilning uchinchi darajali tuzilishi l H zanjiri va –T S zanjirining nisbati quyidagilarga bog'liq: l l l oqsil molekulasining globulaga buklanishi jarayonida o'rnatilgan kovalent bo'lmagan bog'lanishlar soniga, demak, polipeptid zanjirining uzunligiga va haroratga. T |T S| ortishi bilan ertami-kechmi | dan oshadi N| va fazoviy struktura barqarorlikni yo'qotadi - oqsilning termal denatürasyonu
Oqsilning uchinchi darajali tuzilishi l H zanjiri va –T S zanjirining nisbati quyidagilarga bog'liq: l l l oqsil molekulasining globulaga buklanishi jarayonida o'rnatilgan kovalent bo'lmagan bog'lanishlar soniga, demak, polipeptid zanjirining uzunligiga va haroratga. T |T S| ortishi bilan ertami-kechmi | dan oshadi N| va fazoviy struktura barqarorlikni yo'qotadi - oqsilning termal denatürasyonu
 Oqsilning uchlamchi tuzilishi l l Globul burmalanganda peptid zanjirining entropiyasi kamayadi, lekin shu bilan birga erituvchi suvning entropiyasi ortadi.Bu uchinchi darajali strukturani barqarorlashtirishda hal qiluvchi rol o'ynaydi.
Oqsilning uchlamchi tuzilishi l l Globul burmalanganda peptid zanjirining entropiyasi kamayadi, lekin shu bilan birga erituvchi suvning entropiyasi ortadi.Bu uchinchi darajali strukturani barqarorlashtirishda hal qiluvchi rol o'ynaydi.
 Proteinning uchinchi darajali tuzilishi l l Suv noodatiy, yuqori tuzilishga ega erituvchi (?) Proteinning qutbli guruhlari suvning muzsimon tuzilmalariga osonlik bilan kirib, u bilan vodorod aloqalarini o'rnatadi.Polar bo'lmagan guruhlar suv molekulalari bog'langan holda hosil bo'lgan o'ziga xos bo'shliqlarni egallaydi. Bularning barchasi ko'p miqdordagi suv molekulalarining xaotik harakatini istisno qiladi, ya'ni tartiblash sodir bo'ladi, suvning entropiyasi kamayadi.
Proteinning uchinchi darajali tuzilishi l l Suv noodatiy, yuqori tuzilishga ega erituvchi (?) Proteinning qutbli guruhlari suvning muzsimon tuzilmalariga osonlik bilan kirib, u bilan vodorod aloqalarini o'rnatadi.Polar bo'lmagan guruhlar suv molekulalari bog'langan holda hosil bo'lgan o'ziga xos bo'shliqlarni egallaydi. Bularning barchasi ko'p miqdordagi suv molekulalarining xaotik harakatini istisno qiladi, ya'ni tartiblash sodir bo'ladi, suvning entropiyasi kamayadi.
 Proteinning uchinchi darajali tuzilishi l l l Ammo oqsilning qutb bo'lmagan guruhlari molekuladan "olib tashlanishi" mumkin emasligi sababli, globula buklanadi, bunda gidrofobik guruhlarning muhim qismi (kamida ½) suv bilan aloqa qilishdan yashiriladi. Bu hidrofobik kontaktlarning o'rnatilishiga olib keladi, van der Waals kuchlari Protein entropiyasining pasayishi. Globulaning paydo bo'lishining natijasi vodorod aloqalarining muhim qismini va yon guruhlarning suv molekulalari bilan gidrofobik o'zaro ta'sirini yo'q qilishdir. Ikkinchisining harakatining tasodifiyligi oshadi va suvning entropiyasi ortadi
Proteinning uchinchi darajali tuzilishi l l l Ammo oqsilning qutb bo'lmagan guruhlari molekuladan "olib tashlanishi" mumkin emasligi sababli, globula buklanadi, bunda gidrofobik guruhlarning muhim qismi (kamida ½) suv bilan aloqa qilishdan yashiriladi. Bu hidrofobik kontaktlarning o'rnatilishiga olib keladi, van der Waals kuchlari Protein entropiyasining pasayishi. Globulaning paydo bo'lishining natijasi vodorod aloqalarining muhim qismini va yon guruhlarning suv molekulalari bilan gidrofobik o'zaro ta'sirini yo'q qilishdir. Ikkinchisining harakatining tasodifiyligi oshadi va suvning entropiyasi ortadi

 Oqsilning to'rtlamchi tuzilishi... l l l Oligomer oqsillar to'rtlamchi tuzilishga ega bo'lib, ularning molekulalari kovalent bo'lmagan bog'langan ikki yoki undan ortiq polipeptid zanjirlaridan iborat.To'rtlamchi tuzilish - oqsilning alohida polipeptid zanjirlari orqali hosil bo'lgan o'zaro ta'sir qiluvchi bo'linmalarning fazoda joylashishi. Subbirliklar orasidagi o'zaro ta'sir etarlicha kuchli, shuning uchun ularning ansambli ( globulalar ansambli) bitta molekula vazifasini bajaradi, shu bilan birga birlashgan globullarning har biri muhim avtonomiyani saqlab qoladi.
Oqsilning to'rtlamchi tuzilishi... l l l Oligomer oqsillar to'rtlamchi tuzilishga ega bo'lib, ularning molekulalari kovalent bo'lmagan bog'langan ikki yoki undan ortiq polipeptid zanjirlaridan iborat.To'rtlamchi tuzilish - oqsilning alohida polipeptid zanjirlari orqali hosil bo'lgan o'zaro ta'sir qiluvchi bo'linmalarning fazoda joylashishi. Subbirliklar orasidagi o'zaro ta'sir etarlicha kuchli, shuning uchun ularning ansambli ( globulalar ansambli) bitta molekula vazifasini bajaradi, shu bilan birga birlashgan globullarning har biri muhim avtonomiyani saqlab qoladi.

 Oqsilning to'rtlamchi tuzilishi l l Oqsilning to'rtlamchi tuzilishini tashkil etuvchi bo'linmalar tuzilishi va funksiyasi jihatidan butunlay boshqacha bo'lishi mumkin - geteromer oqsillar.Bu bir strukturada o'zaro bog'liq bo'lgan bir nechta funktsiyalarni birlashtirib, ko'p funktsiyali molekula hosil qilish imkonini beradi l l Misol. Protein kinazasi: C-kinaza fermentativ faollik uchun javob beradi, R-subbirlik esa tartibga soluvchidir.Gomerik oqsillarda subbirliklar bir xil bo'ladi.
Oqsilning to'rtlamchi tuzilishi l l Oqsilning to'rtlamchi tuzilishini tashkil etuvchi bo'linmalar tuzilishi va funksiyasi jihatidan butunlay boshqacha bo'lishi mumkin - geteromer oqsillar.Bu bir strukturada o'zaro bog'liq bo'lgan bir nechta funktsiyalarni birlashtirib, ko'p funktsiyali molekula hosil qilish imkonini beradi l l Misol. Protein kinazasi: C-kinaza fermentativ faollik uchun javob beradi, R-subbirlik esa tartibga soluvchidir.Gomerik oqsillarda subbirliklar bir xil bo'ladi.
 To'rtlamchi oqsil tuzilishi l Bo'limlararo kontaktlar - kovalent bo'lmagan o'zaro ta'sirlarning juda rivojlangan tizimi l l l l l Hidrofobik guruhlar va subbirliklarning (aloqa joylari yoki zonalari) yuzasidagi butun hidrofobik hududlar o'rtasidagi aloqalar, bu tizim entropiyasining oshishiga olib keladi va a. muvozanatning oligomer vodorod bog'lari tomon siljishi yon guruhlar orasidagi elektrostatik o'zaro ta'sirlar To'rtlamchi struktura uchinchi tuzilishga qaraganda kamroq kuchli, chunki hidrofobik kontaktlarning hissasi kamroq.
To'rtlamchi oqsil tuzilishi l Bo'limlararo kontaktlar - kovalent bo'lmagan o'zaro ta'sirlarning juda rivojlangan tizimi l l l l l Hidrofobik guruhlar va subbirliklarning (aloqa joylari yoki zonalari) yuzasidagi butun hidrofobik hududlar o'rtasidagi aloqalar, bu tizim entropiyasining oshishiga olib keladi va a. muvozanatning oligomer vodorod bog'lari tomon siljishi yon guruhlar orasidagi elektrostatik o'zaro ta'sirlar To'rtlamchi struktura uchinchi tuzilishga qaraganda kamroq kuchli, chunki hidrofobik kontaktlarning hissasi kamroq.

 To'rtlamchi tuzilmaning funktsiyalari 1. Bir tuzilishda bir nechta o'zaro bog'liq funktsiyalarning kombinatsiyasi 2. Arxitektura funktsiyasi 1. Oqsilning (ferritin) o'ziga xos funktsional imkoniyatlarini ta'minlaydigan juda murakkab konfiguratsiyaning fazoviy shakllanishlarini shakllantirish 2. Ketma-ket reaktsiyalarning umumiy ko'rsatkichlari. fermentlar tomonidan 3. Funktsional markazlarning shakllanishi (fermentlarning faol markazlari)
To'rtlamchi tuzilmaning funktsiyalari 1. Bir tuzilishda bir nechta o'zaro bog'liq funktsiyalarning kombinatsiyasi 2. Arxitektura funktsiyasi 1. Oqsilning (ferritin) o'ziga xos funktsional imkoniyatlarini ta'minlaydigan juda murakkab konfiguratsiyaning fazoviy shakllanishlarini shakllantirish 2. Ketma-ket reaktsiyalarning umumiy ko'rsatkichlari. fermentlar tomonidan 3. Funktsional markazlarning shakllanishi (fermentlarning faol markazlari)
 To'rtlamchi strukturaning funktsiyalari 3. Kengaytirilgan tuzilmalar bilan bir nechta oqsil o'zaro ta'sirini ta'minlash l DNKni bog'lovchi oqsillar - dimerlar (?) 4. Regulyatsiya funktsiyasi. Effektni o'tkazish (substrat bilan o'zaro ta'sir qilish paytida uchinchi darajali tuzilmaning buzilishi) bir bo'linmadan ikkinchisiga o'tadi, bu esa butun to'rtlamchi strukturaning qayta tuzilishiga olib keladi.
To'rtlamchi strukturaning funktsiyalari 3. Kengaytirilgan tuzilmalar bilan bir nechta oqsil o'zaro ta'sirini ta'minlash l DNKni bog'lovchi oqsillar - dimerlar (?) 4. Regulyatsiya funktsiyasi. Effektni o'tkazish (substrat bilan o'zaro ta'sir qilish paytida uchinchi darajali tuzilmaning buzilishi) bir bo'linmadan ikkinchisiga o'tadi, bu esa butun to'rtlamchi strukturaning qayta tuzilishiga olib keladi.


 1. Har bir alohida oqsil o'ziga xos tuzilish bilan tavsiflanadi, bu uning funktsiyalarining o'ziga xosligini ta'minlaydi 2. Shuning uchun turli oqsillarning tuzilishini yoritish tirik tizimlarning tabiatini va shunga mos ravishda hayotning mohiyatini tushunish uchun kalit bo'lib xizmat qilishi mumkin.
1. Har bir alohida oqsil o'ziga xos tuzilish bilan tavsiflanadi, bu uning funktsiyalarining o'ziga xosligini ta'minlaydi 2. Shuning uchun turli oqsillarning tuzilishini yoritish tirik tizimlarning tabiatini va shunga mos ravishda hayotning mohiyatini tushunish uchun kalit bo'lib xizmat qilishi mumkin.
 Adabiyot l l l l Berezov T. T., Korovkin B. F. Biologik kimyo. – M .: Tibbiyot, 1983 Metzler D. Biokimyo. Tirik hujayradagi kimyoviy reaksiyalar. 3 jildda - T. 1, 2. - M.: Mir, 1980 Ovchinnikov Yu. A. Bioorganik kimyo. – M.: Ta'lim, 1987 Biokimyo asoslari / Ed. A. A. Anisimova. – M .: Oliy maktab, 1986 Rees E., Strenberg M. Hujayralardan atomlarga. Molekulyar biologiyaga tasvirlangan kirish. – M.: Mir, 1988 Stepanov V. M. Molekulyar biologiya. Oqsillarning tuzilishi va funktsiyalari. – M.: Oliy maktab, 1996 Filippovich Yu. B. Biokimyo asoslari. – M.: Oliy maktab, 1993 yil
Adabiyot l l l l Berezov T. T., Korovkin B. F. Biologik kimyo. – M .: Tibbiyot, 1983 Metzler D. Biokimyo. Tirik hujayradagi kimyoviy reaksiyalar. 3 jildda - T. 1, 2. - M.: Mir, 1980 Ovchinnikov Yu. A. Bioorganik kimyo. – M.: Ta'lim, 1987 Biokimyo asoslari / Ed. A. A. Anisimova. – M .: Oliy maktab, 1986 Rees E., Strenberg M. Hujayralardan atomlarga. Molekulyar biologiyaga tasvirlangan kirish. – M.: Mir, 1988 Stepanov V. M. Molekulyar biologiya. Oqsillarning tuzilishi va funktsiyalari. – M.: Oliy maktab, 1996 Filippovich Yu. B. Biokimyo asoslari. – M.: Oliy maktab, 1993 yil
Protein - bu bir-biri bilan peptid bog'lari bilan bog'langan aminokislotalarning ketma-ketligi.
Tasavvur qilish osonki, aminokislotalar soni har xil bo'lishi mumkin: kamida ikkitadan har qanday oqilona qiymatlargacha. Biokimyogarlar, agar aminokislotalar soni 10 dan oshmasa, unda bunday birikma peptid deb ataladi; agar 10 yoki undan ortiq aminokislotalar bo'lsa - polipeptid. Konformatsiya deb ataladigan ma'lum fazoviy tuzilmani o'z-o'zidan hosil qilish va saqlab turish qobiliyatiga ega bo'lgan polipeptidlar oqsillar deb tasniflanadi. Bunday tuzilmani barqarorlashtirish faqat polipeptidlar ma'lum bir uzunlikka (40 dan ortiq aminokislotalar) erishganda mumkin; shuning uchun molekulyar og'irligi 5000 Da dan ortiq bo'lgan polipeptidlar odatda oqsillar hisoblanadi. (1Da uglerod izotopining 1/12 qismiga teng). Oqsil faqat ma'lum fazoviy tuzilishga (nativ tuzilishga) ega bo'lgan holda o'z vazifalarini bajarishi mumkin.
Proteinning o'lchamini daltonlarda (molekulyar og'irlik) o'lchash mumkin, bu ko'pincha molekulaning hosil bo'lgan birliklarida, kilodaltonlarda (kDa) nisbatan kattaligi bilan bog'liq. Xamirturush oqsillari o'rtacha 466 aminokislotadan iborat va molekulyar og'irligi 53 kDa ni tashkil qiladi. Hozirgi vaqtda ma'lum bo'lgan eng katta protein, titin, mushak sarkomerlarining tarkibiy qismidir; Uning turli xil izoformalarining molekulyar og'irligi 3000 dan 3700 kDa gacha o'zgarib turadi va u 38138 aminokislotadan iborat (odamning solius mushaklarida).
Protein tuzilishi
Proteinning uch o'lchovli tuzilishi katlama jarayonida hosil bo'ladi. katlama -"katlama") Pastki darajadagi tuzilmalarning o'zaro ta'siri natijasida uch o'lchovli struktura hosil bo'ladi.
Protein tuzilishining to'rtta darajasi mavjud:
Birlamchi tuzilma- polipeptid zanjiridagi aminokislotalarning ketma-ketligi.
Ikkilamchi tuzilma- bu polipeptid zanjirining alohida bo'limlarini bo'shliqda joylashtirish.
Protein ikkilamchi tuzilishining eng keng tarqalgan turlari quyidagilardir:
a-spirallar- molekulaning uzun o'qi atrofida zich burilishlar, bir burilish 3,6 aminokislota qoldig'idan iborat va spiralning qadami 0,54 nm (har bir aminokislota qoldig'iga 0,15 nm), spiral vodorod bog'lari bilan barqarorlashadi. H va O peptid guruhlari bir-biridan 4 ta aminokislota qoldig'i bilan ajralib turadi. Spiral faqat bitta turdagi aminokislotalar stereoizomeridan (L) qurilgan. U chap qo'l yoki o'ng qo'l bo'lishi mumkin bo'lsa-da, oqsillarda o'ng qo'l ustunlik qiladi. Spiral glutamik kislota, lizin va argininning elektrostatik o'zaro ta'siri bilan buziladi. Bir-biriga yaqin joylashgan asparagin, serin, treonin va leysin qoldiqlari spiral shakllanishiga sterik ravishda xalaqit berishi mumkin, prolin qoldiqlari zanjirning egilishiga olib keladi, shuningdek, a-spiral tuzilishini buzadi.
b-plifli qatlamlar- aminokislotalar yoki birlamchi tuzilishda bir-biridan nisbatan uzoqda bo'lgan (har bir aminokislota qoldig'iga 0,347 nm) turli xil oqsil zanjirlari o'rtasida vodorod aloqalari hosil bo'lgan bir nechta zigzag polipeptid zanjirlari, a dagi kabi bir-biriga yaqin joylashmagan. - spiral. Ushbu zanjirlar odatda N-terminal uchlari qarama-qarshi yo'nalishda (parallelga qarshi yo'nalish) bo'ladi. Kichik o'lchamdagi aminokislotalar yon guruhlari b-varaqlar hosil bo'lishi uchun muhimdir; odatda glitsin va alanin ustunlik qiladi.

Proteinning b-pleated varag'iga katlanması
Tartibsiz tuzilmalar - kosmosda oqsil zanjirining tartibsiz joylashishi.
Har bir oqsilning fazoviy tuzilishi individualdir va uning asosiy tuzilishi bilan belgilanadi. Biroq, turli tuzilish va funktsiyalarga ega bo'lgan oqsillarning konformatsiyasini taqqoslash ularda ikkilamchi tuzilish elementlarining o'xshash birikmalarining mavjudligini aniqladi. Ikkilamchi tuzilmalar hosil bo'lishining bu o'ziga xos tartibi oqsillarning ikkinchi darajali tuzilishi deb ataladi. Supero'rta tuzilma radikallararo o'zaro ta'sirlar tufayli shakllanadi.
a-spiral va b-tuzilmalarning ma'lum xarakterli birikmalari ko'pincha "strukturaviy motivlar" deb ataladi. Ularning o'ziga xos nomlari bor: "a-spiral-burilish-a-spiral", "a/b-barrel tuzilishi", "leytsinli fermuar", "sink barmoq" va boshqalar.
Uchinchi darajali tuzilish- Bu butun polipeptid zanjirini kosmosga joylashtirish usuli. Uchinchi darajali struktura a-spirallar, b-plitsimon varaqlar va supersekondar tuzilmalar bilan bir qatorda molekulaning muhim qismini egallashi mumkin bo'lgan tartibsiz konformatsiyani aniqlaydi.

Proteinning uchinchi darajali tuzilishga aylanishining sxematik tasviri.
To'rtlamchi tuzilish bir nechta polipeptid zanjirlaridan (subbirliklar, protomerlar yoki monomerlar) tashkil topgan oqsillarda, bu subbirliklarning uchinchi darajali tuzilmalari birlashganda paydo bo'ladi. Masalan, gemoglobin molekulasi 4 ta subbirlikdan iborat. Supramolekulyar shakllanishlar to'rtlamchi tuzilishga ega - ko'p fermentli komplekslar, ular bir nechta fermentlar va kofermentlar (piruvat dehidrogenaza) va izofermentlar (laktat dehidrogenaza - LDH, kreatin fosfokinaz - CPK) dan iborat.
Shunday qilib. Fazoviy struktura polipeptid zanjirining uzunligiga bog'liq emas, balki har bir oqsilga xos bo'lgan aminokislotalar qoldiqlari ketma-ketligiga, shuningdek, tegishli aminokislotalarga xos bo'lgan yon radikallarga bog'liq. Oqsil makromolekulalarining fazoviy uch o‘lchamli tuzilishi yoki konformatsiyasi birinchi navbatda vodorod bog‘lari, aminokislotalarning qutb bo‘lmagan yon radikallari orasidagi hidrofobik o‘zaro ta’sirlar va aminokislotalar qoldiqlarining qarama-qarshi zaryadlangan yon guruhlari orasidagi ionli o‘zaro ta’sirlar natijasida hosil bo‘ladi. Vodorod aloqalari oqsil makromolekulasining fazoviy tuzilishini shakllantirish va saqlashda katta rol o'ynaydi.
Hidrofobik o'zaro ta'sirlarga kelsak, ular suv molekulalari orasidagi vodorod aloqalarini uzishga qodir bo'lmagan qutbsiz radikallar o'rtasidagi aloqa natijasida yuzaga keladi, ular oqsil globulasi yuzasiga joylashadi. Protein sintezi davom etar ekan, globulaning ichida qutbsiz kimyoviy guruhlar to'planadi va qutblilar uning yuzasiga chiqib ketadi. Shunday qilib, oqsil molekulasi erituvchining pH darajasiga va oqsil tarkibidagi ion guruhlariga qarab neytral, musbat zaryadlangan yoki manfiy zaryadlangan bo'lishi mumkin. Bundan tashqari, oqsillarning konformatsiyasi ikkita sistein qoldig'i o'rtasida hosil bo'lgan kovalent S-S aloqalari bilan ta'minlanadi. Oqsilning tabiiy tuzilishining shakllanishi natijasida polipeptid zanjirining uzoq qismlarida joylashgan ko'plab atomlar yaqinlashadi va bir-biriga ta'sir qilib, alohida aminokislotalar yoki kichik polipeptidlarda mavjud bo'lmagan yangi xususiyatlarga ega bo'ladi.
Qatlamlanish - oqsillarning (va boshqa biomakromolekulyarlarning) ochilmagan konformatsiyadan "mahalliy" shaklga o'ralishi - fizik va kimyoviy jarayon ekanligini tushunish muhimdir, buning natijasida oqsillar o'zlarining tabiiy "yashash joylarida" (eritma, sitoplazma yoki membrana) faqat fazoviy joylashuvi va funktsiyalariga xos bo'lgan xususiyatlarga ega bo'ladi.
Hujayralarda katalitik jihatdan faol bo'lmagan bir qator oqsillar mavjud bo'lib, ular fazoviy oqsil tuzilmalarining shakllanishiga katta hissa qo'shadilar. Bular chaperonlar deb ataladi. Chaperonlar qisman katlanmış polipeptid zanjiri bilan qaytariladigan kovalent bo'lmagan komplekslarni hosil qilish orqali uch o'lchovli oqsil konformatsiyasini to'g'ri yig'ishga yordam beradi, shu bilan birga funktsional faol bo'lmagan oqsil tuzilmalarining shakllanishiga olib keladigan noto'g'ri shakllangan aloqalarni inhibe qiladi. Chaperonlarga xos bo'lgan funktsiyalar ro'yxati eritilgan (qisman buklangan) globullarni agregatsiyadan himoya qilishni, shuningdek, yangi sintezlangan oqsillarni turli hujayra lokuslariga o'tkazishni o'z ichiga oladi.
Chaperonlar asosan issiqlik zarbasi oqsillari bo'lib, ularning sintezi stressli harorat ta'sirida keskin kuchayadi, shuning uchun ularni hsp (issiqlik zarbasi oqsillari) deb ham atashadi. Ushbu oqsillarning oilalari mikroblar, o'simliklar va hayvonlar hujayralarida mavjud. Chaperonlarning tasnifi ularning molekulyar og'irligiga asoslanadi, bu 10 dan 90 kDa gacha. Ular oqsillarning uch o'lchovli tuzilishini shakllantirishga yordam beradigan oqsillardir. Chaperonlar yangi sintez qilingan polipeptid zanjirini ochilmagan holatda ushlab turadi, uning tabiiydan farqli shaklga aylanishiga yo'l qo'ymaydi va yagona to'g'ri, mahalliy protein tuzilishi uchun sharoit yaratadi.
Protein katlamasi jarayonida molekulaning ba'zi konformatsiyalari erigan globul bosqichida rad etiladi. Bunday molekulalarning parchalanishi oqsil ubiquitin tomonidan boshlanadi.
Ubiquitin yo'li orqali oqsilning parchalanishi ikkita asosiy bosqichni o'z ichiga oladi:
1) ubiquitinning qoldiq orqali parchalanadigan oqsilga kovalent biriktirilishi lizin, oqsilda bunday tegning mavjudligi hosil bo'lgan konjugatlarni proteazomalarga yo'naltiruvchi birlamchi saralash signalidir, aksariyat hollarda oqsilga ipda boncuklar shaklida tashkil etilgan bir nechta ubiquitin molekulalari biriktiriladi;
2) proteazoma tomonidan oqsil gidrolizi (proteazomaning asosiy vazifasi keraksiz va shikastlangan oqsillarni qisqa peptidlarga proteolitik degradatsiyadan iborat). Ubiquitin haqli ravishda oqsillar uchun "o'lim belgisi" deb ataladi.
Dom?n sincap? - oqsilning uchinchi darajali strukturasining elementi, oqsilning etarlicha barqaror va mustaqil quyi tuzilishi bo'lib, uning katlanishi boshqa qismlardan mustaqil ravishda sodir bo'ladi. Domen odatda bir nechta ikkilamchi tuzilish elementlarini o'z ichiga oladi. Strukturaviy jihatdan oʻxshash domenlar nafaqat oʻzaro bogʻlangan oqsillarda (masalan, turli hayvonlarning gemoglobinlarida), balki butunlay boshqa oqsillarda ham uchraydi. Protein bir nechta domenlarga ega bo'lishi mumkin va bu hududlar bir xil proteinda turli funktsiyalarni bajarishi mumkin. Ba'zi fermentlar va barcha immunoglobulinlar domen tuzilishiga ega. Uzoq polipeptid zanjirli oqsillar (200 dan ortiq aminokislota qoldiqlari) ko'pincha domen tuzilmalarini yaratadi.
Proteinlar organik moddalardir. Ushbu yuqori molekulyar birikmalar ma'lum bir tarkibga ega bo'lib, gidrolizlanganda aminokislotalarga parchalanadi. Protein molekulalari turli xil shakllarda bo'lishi mumkin, ularning aksariyati bir nechta polipeptid zanjirlaridan iborat. Oqsilning tuzilishi haqidagi ma'lumotlar DNKda kodlangan bo'lib, oqsil molekulalarini sintez qilish jarayoni translatsiya deb ataladi.
Proteinlarning kimyoviy tarkibi
O'rtacha protein tarkibiga quyidagilar kiradi:
- 52% uglerod;
- 7% vodorod;
- 12% azot;
- 21% kislorod;
- 3% oltingugurt.
Protein molekulalari polimerlardir. Ularning tuzilishini tushunish uchun ularning monomerlari - aminokislotalar nima ekanligini bilish kerak.
Aminokislotalar
Ular odatda ikki toifaga bo'linadi: doimiy va vaqti-vaqti bilan sodir bo'ladigan. Birinchisi 18 va yana 2 ta amidni o'z ichiga oladi: aspartik va glutamik kislota. Ba'zida faqat uchta kislota topiladi.
Bu kislotalarni turli yo'llar bilan tasniflash mumkin: yon zanjirlarning tabiati yoki ularning radikallarining zaryadiga ko'ra, ularni CN va COOH guruhlari soniga ham bo'lish mumkin.
Birlamchi protein tuzilishi
Protein zanjiridagi aminokislotalarning almashinish tartibi uning keyingi tashkiliy darajalari, xossalari va funktsiyalarini belgilaydi. Monomerlar orasidagi asosiysi peptiddir. U bir aminokislotadan vodorod va boshqasidan OH guruhini olish natijasida hosil bo'ladi.
Protein molekulasining tashkil etilishining birinchi darajasi undagi aminokislotalarning ketma-ketligi, oddiygina oqsil molekulalarining tuzilishini belgilovchi zanjirdir. U muntazam tuzilishga ega bo'lgan "skelet" dan iborat. Bu takrorlanuvchi ketma-ketlik -NH-CH-CO-. Alohida yon zanjirlar aminokislota radikallari (R) bilan ifodalanadi, ularning xossalari oqsil strukturasining tarkibini aniqlaydi.
Oqsil molekulalarining tuzilishi bir xil bo'lsa ham, ular monomerlari zanjirda boshqa ketma-ketlikka ega bo'lgani uchungina xossalari bilan farq qilishi mumkin. Proteindagi aminokislotalarning tartibi genlar tomonidan belgilanadi va oqsilga ma'lum biologik funktsiyalarni buyuradi. Xuddi shu funktsiya uchun javob beradigan molekulalardagi monomerlarning ketma-ketligi ko'pincha turli turlarda o'xshashdir. Bunday molekulalar tashkiliy jihatdan bir xil yoki o'xshash bo'lib, har xil turdagi organizmlarda - gomologik oqsillarda bir xil funktsiyalarni bajaradi. Kelajakdagi molekulalarning tuzilishi, xususiyatlari va funktsiyalari aminokislotalar zanjirining sintezi bosqichida allaqachon o'rnatilgan.
Ba'zi umumiy xususiyatlar
Oqsillarning tuzilishi uzoq vaqt davomida o'rganilib, ularning birlamchi tuzilishini tahlil qilish ba'zi umumlashtirishlarni amalga oshirish imkonini berdi. Ko'proq miqdordagi oqsillar yigirmata aminokislotalarning mavjudligi bilan tavsiflanadi, ularda ayniqsa ko'p glitsin, alanin, glutamin va ozgina triptofan, arginin, metionin va histidin mavjud. Istisno faqat oqsillarning ba'zi guruhlari, masalan, gistonlardir. Ular DNKni qadoqlash uchun zarur va ko'p histidinni o'z ichiga oladi.
Organizmlar harakatining har qanday turi (mushaklar ishi, hujayradagi protoplazmaning harakatlanishi, oddiy hujayralardagi kirpiklarning miltillashi va boshqalar) oqsillar tomonidan amalga oshiriladi. Oqsillarning tuzilishi ularning harakatlanishi va tolalar va halqalarni hosil qilishiga imkon beradi.

Tashish funktsiyasi shundaki, ko'plab moddalar hujayra membranasi bo'ylab maxsus tashuvchi oqsillar orqali tashiladi.
Ushbu polimerlarning gormonal roli darhol aniq: bir qator gormonlar tuzilishdagi oqsillar, masalan, insulin, oksitotsin.
Zaxira funktsiyasi oqsillarning konlarni hosil qila olishi bilan belgilanadi. Masalan, tuxum valgumini, sut kazeini, o'simlik urug'i oqsillari - ular ko'p miqdorda ozuqa moddalarini saqlaydi.
Barcha tendonlar, artikulyar bo'g'inlar, skelet suyaklari va tuyoqlar oqsillardan hosil bo'ladi, bu bizni ularning keyingi funktsiyasiga - tayanchga olib keladi.
Protein molekulalari retseptorlar bo'lib, ma'lum moddalarni tanlab tan oladi. Bu rol uchun glikoproteinlar va lektinlar ayniqsa mashhur.
Immunitetning eng muhim omillari antikorlar va oqsillardir. Masalan, qon ivish jarayoni fibrinogen oqsilidagi o'zgarishlarga asoslanadi. Qizilo'ngach va oshqozonning ichki devorlari shilliq oqsillar - litsinlarning himoya qatlami bilan qoplangan. Toksinlar ham kelib chiqishi oqsillaridir. Hayvonlar tanasini himoya qiluvchi terining asosini kollagen tashkil qiladi. Bu protein funktsiyalarining barchasi himoyadir.

Xo'sh, oxirgi funktsiya tartibga solishdir. Genomning ishlashini nazorat qiluvchi oqsillar mavjud. Ya'ni, ular transkripsiya va tarjimani tartibga soladi.
Proteinlar qanchalik muhim rol o'ynamasin, oqsillarning tuzilishi olimlar tomonidan ancha oldin ochilgan. Va endi ular bu bilimlardan foydalanishning yangi usullarini kashf qilmoqdalar.


