Iš ko susideda DNR? Kas yra nukleotidas? Nukleotidų sudėtis, struktūra, skaičius ir seka DNR grandinėje Dvigubos DNR spiralės stabilizuoja jonus
Nukleorūgštys yra didelės molekulinės masės medžiagos, susidedančios iš mononukleotidų, kurie yra sujungti vienas su kitu polimero grandinėje naudojant 3 ", 5" - fosfodiesterio ryšius ir yra tam tikru būdu supakuoti ląstelėse.
Nukleino rūgštys yra dviejų tipų biopolimerai: ribonukleorūgštis (RNR) ir dezoksiribonukleorūgštis (DNR). Kiekvienas biopolimeras susideda iš nukleotidų, kurie skiriasi angliavandenių liekana (riboze, dezoksiriboze) ir viena iš azoto bazių (uracilo, timino). Pagal šiuos skirtumus nukleorūgštys gavo savo pavadinimą.
Dezoksiribonukleino rūgšties struktūra
Nukleino rūgštys turi pirminę, antrinę ir tretinę struktūrą.
Pirminė DNR struktūra
Pirminė DNR struktūra vadinama linijine polinukleotidų grandine, kurioje mononukleotidai yra sujungti 3 ", 5" -fosfodiesterio ryšiais. Pradinė medžiaga nukleorūgščių grandinės surinkimui ląstelėje yra nukleozidas 5"-trifosfatas, kuris, pašalinus β ir γ fosforo rūgšties likučius, gali prijungti kito nukleozido 3" anglies atomą. . Taigi, vienos dezoksiribozės 3 "anglies atomas kovalentiškai jungiasi su 5" kitos dezoksiribozės anglies atomu per vieną fosforo rūgšties liekaną ir sudaro linijinę polinukleotidinę nukleorūgšties grandinę. Iš čia ir kilo pavadinimas: 3 ", 5" -fosfodiesterio jungtys. Azoto bazės nedalyvauja vienos grandinės nukleotidų kombinacijoje (1 pav.).
Dėl tokio ryšio tarp likusios vieno nukleotido fosforo rūgšties molekulės ir kito angliavandenių susidaro polinukleotido molekulės pentozės-fosfato karkasas, prie kurio šone viena po kitos jungiasi azoto bazės. Jų išsidėstymo seka nukleorūgščių molekulių grandinėse yra griežtai specifinė skirtingų organizmų ląstelėms, t.y. yra specifinio pobūdžio (Chargaffo taisyklė).
Linijinė DNR grandinė, kurios ilgis priklauso nuo grandinėje esančių nukleotidų skaičiaus, turi du galus: vienas vadinamas 3 "galu ir turi laisvą hidroksilį, o kitame, 5" gale, yra fosforo rūgštis. likutis. Grandinė yra poliarizuota ir gali turėti 5 "-> 3" ir 3 "-> 5" kryptį. Išimtis yra žiedinė DNR.
Genetinį DNR „tekstą“ sudaro kodiniai „žodžiai“ – nukleotidų tripletai, vadinami kodonais. DNR sritys, kuriose yra informacijos apie pirminę visų tipų RNR struktūrą, vadinamos struktūriniais genais.
Polinukleoditinės DNR grandinės pasiekia milžiniškus dydžius, todėl ląstelėje yra tam tikru būdu supakuotos.
Tyrinėdamas DNR sudėtį, Chargaffas (1949) nustatė svarbius įstatymus dėl atskirų DNR bazių turinio. Jie padėjo atskleisti antrinę DNR struktūrą. Šie modeliai vadinami Chargaff taisyklėmis. Chargaff taisyklės
Šios taisyklės rodo, kad kuriant DNR reikia laikytis gana griežtos atitikties (poravimosi) tarp purino ir pirimidino bazių apskritai, o konkrečiai timino su adeninu ir citozino su guaninu. Remdamiesi šiomis taisyklėmis, įskaitant 1953 m., Watsonas ir Crickas pasiūlė antrinės DNR struktūros modelį, vadinamą dviguba spirale (pav.). |

Antrinė DNR struktūra
Antrinė DNR struktūra yra dviguba spiralė, kurios modelį 1953 metais pasiūlė D. Watsonas ir F. Crickas.
Būtinos sąlygos DNR modeliui sukurti
Pirminės analizės metu susidarė įspūdis, kad bet kokios kilmės DNR turi visus keturis nukleotidus vienodais moliniais kiekiais. Tačiau praėjusio amžiaus ketvirtajame dešimtmetyje E. Chargaffas ir jo bendradarbiai, atlikę iš įvairių organizmų išskirtos DNR analizės rezultatus, aiškiai parodė, kad azoto bazių juose yra įvairiais kiekybiniais santykiais. Chargaffas nustatė, kad nors šie santykiai yra vienodi DNR iš visų to paties tipo organizmo ląstelių, skirtingų rūšių DNR gali labai skirtis tam tikrų nukleotidų kiekiu. Tai leido manyti, kad azoto bazių santykio skirtumai gali būti susiję su tam tikru biologiniu kodu. Nors atskirų purino ir pirimidino bazių santykis skirtinguose DNR mėginiuose pasirodė skirtingas, lyginant analizės rezultatus išryškėjo tam tikras modelis: visuose mėginiuose bendras purinų kiekis buvo lygus bendram pirimidinų kiekiui (A + G = T + C), adenino kiekis buvo lygus timino kiekiui (A = T), o guanino kiekis - citozino kiekiui (G = C). DNR, išskirta iš žinduolių ląstelių, paprastai buvo turtingesnė adenino ir timino ir santykinai skurdesne guanino ir citozino, o bakterijų DNR buvo turtingesnė guanino ir citozino ir santykinai skurdesne adenino ir timino. Šie duomenys sudarė svarbią faktinės medžiagos dalį, kurios pagrindu vėliau buvo sukurtas Watson-Crick DNR struktūros modelis.
Kitas svarbus netiesioginis galimos DNR struktūros požymis buvo L. Paulingo duomenys apie baltymų molekulių sandarą. Paulingas parodė, kad galimos kelios skirtingos stabilios aminorūgščių grandinės konfigūracijos baltymo molekulėje. Viena iš įprastų peptidinės grandinės konfigūracijų, α-spiralė, yra taisyklinga spiralinė struktūra. Esant tokiai struktūrai, gali susidaryti vandeniliniai ryšiai tarp aminorūgščių, esančių gretimuose grandinės posūkiuose. Paulingas aprašė polipeptidinės grandinės α-spiralinę konfigūraciją 1950 m. ir pasiūlė, kad DNR molekulės taip pat tikriausiai turi spiralinę struktūrą, fiksuotą vandeniliniais ryšiais.
Tačiau vertingiausią informaciją apie DNR molekulės sandarą suteikė rentgeno struktūrinės analizės rezultatai. Rentgeno spinduliai, praeinantys per DNR kristalą, patiria difrakciją, tai yra, jie nukreipiami tam tikromis kryptimis. Spindulių nukreipimo laipsnis ir pobūdis priklauso nuo pačių molekulių struktūros. Rentgeno spindulių difrakcijos modelis (3 pav.) patyrusiai akiai suteikia nemažai netiesioginių požymių, susijusių su tiriamos medžiagos molekulių sandara. DNR rentgeno spindulių difrakcijos modelių analizė leido padaryti išvadą, kad azoto bazės (turinčios plokščią formą) yra sukrautos kaip plokščių krūva. Rentgeno spindulių difrakcijos modeliai atskleidė tris pagrindinius kristalinės DNR struktūros periodus: 0,34, 2 ir 3,4 nm.

Watson-Crick DNR modelis
Remiantis Chargaff analitiniais duomenimis, Wilkinso gautais rentgeno spindulių difrakcijos modeliais ir chemikų, pateikusių informaciją apie tikslius atstumus tarp molekulėje esančių atomų, apie kampus tarp tam tikro atomo ryšių ir atomų dydžio, tyrimais. Watsonas ir Crickas tam tikru mastu pradėjo kurti atskirų DNR molekulės sudedamųjų dalių fizinius modelius. ir „suderinti“ juos tarpusavyje taip, kad gauta sistema atitiktų skirtingus eksperimentinius duomenis. [Rodyti] .
Dar anksčiau buvo žinoma, kad gretimi nukleotidai DNR grandinėje yra sujungti fosfodiesterio tilteliais, jungiančiais vieno nukleotido dezoksiribozės 5'-anglies atomą su kito nukleotido dezoksiribozės 3'-anglies atomu. Watsonas ir Crickas neabejojo, kad 0,34 nm periodas atitinka atstumą tarp nuoseklių nukleotidų DNR grandinėje. Be to, galima daryti prielaidą, kad 2 nm periodas atitinka grandinės storį. Ir norėdami paaiškinti, kuri tikroji struktūra atitinka 3,4 nm periodą, Watsonas ir Crickas, kaip ir anksčiau Paulingas, pasiūlė, kad grandinė būtų susukta spiralės pavidalu (arba, tiksliau, sudaro sraigtinę liniją, nes spiralė griežtąja šio žodžio prasme šis žodis gaunamas, kai posūkiai erdvėje sudaro kūginį, o ne cilindrinį paviršių). Tada 3,4 nm periodas atitiks atstumą tarp nuoseklių šios spiralės posūkių. Tokia spiralė gali būti labai tanki arba šiek tiek ištempta, tai yra, jos posūkiai gali būti švelnūs arba statūs. Kadangi 3,4 nm periodas yra lygiai 10 kartų didesnis už atstumą tarp nuoseklių nukleotidų (0,34 nm), aišku, kad kiekviename pilname spiralės posūkyje yra 10 nukleotidų. Iš šių duomenų Watsonas ir Crickas sugebėjo apskaičiuoti polinukleotidų grandinės, susuktos į 2 nm skersmens spiralę, kurios atstumas tarp posūkių yra 3,4 nm, tankį. Paaiškėjo, kad tokios grandinės tankis būtų pusė faktinio DNR tankio, kuris jau buvo žinomas. Turėjau daryti prielaidą, kad DNR molekulė susideda iš dviejų gijų – kad tai dviguba nukleotidų spiralė.
Kita užduotis, žinoma, buvo išsiaiškinti erdvinį ryšį tarp dviejų grandinių, sudarančių dvigubą spiralę. Išbandę daugybę grandinių išdėstymo savo fiziniame modelyje, Watsonas ir Crickas nustatė, kad visi turimi duomenys geriausiai tinka variantui, kuriame dvi polinukleotidų spiralės eina priešingomis kryptimis; šiuo atveju grandinės, susidedančios iš cukraus ir fosfatų likučių, sudaro dvigubos spiralės paviršių, o viduje yra purinai ir pirimidinai. Bazės, esančios viena priešais kitą, priklausančios dviem grandinėms, yra sujungtos poromis vandeniliniais ryšiais; būtent šie vandeniliniai ryšiai laiko grandines kartu ir taip fiksuoja bendrą molekulės konfigūraciją.
Dvigubą DNR spiralę galima įsivaizduoti kaip spiralės formos lynų kopėčias, kad jos laipteliai liktų horizontalioje padėtyje. Tada dvi išilginės virvės atitiks cukraus ir fosfato likučių grandines, o skersiniai – azotinių bazių poras, sujungtas vandeniliniais ryšiais.
Tolesnio galimų modelių tyrimo rezultatas, Watsonas ir Crickas padarė išvadą, kad kiekvieną "barą" turėtų sudaryti vienas purinas ir vienas pirimidinas; esant 2 nm periodui (tai atitinka dvigubos spiralės skersmenį), dviem purinams neužtektų vietos, o du pirimidinai nebūtų pakankamai arti vienas kito, kad susidarytų tinkami vandeniliniai ryšiai. Išsamus išsamaus modelio tyrimas parodė, kad adeninas ir citozinas, sudarantys tinkamo dydžio derinį, vis tiek negali būti išdėstyti taip, kad tarp jų susidarytų vandeniliniai ryšiai. Panašūs pranešimai privertė neįtraukti guanino-timino derinio, o adenino-timino ir guanino-citozino deriniai buvo gana priimtini. Vandenilio jungčių prigimtis yra tokia, kad adeninas sudaro porą su timinu, o guaninas - su citozinu. Ši specifinių bazių poravimosi samprata leido paaiškinti „Chargaff“ taisyklę, pagal kurią bet kurioje DNR molekulėje adenino kiekis visada lygus timino kiekiui, o guanino – citozino kiekiui. Tarp adenino ir timino susidaro dvi vandenilinės jungtys, o tarp guanino ir citozino – trys vandenilio ryšiai. Dėl šio vandenilio ryšių formavimosi prieš kiekvieną adeniną vienoje grandinėje timinas randamas kitoje; taip pat prieš kiekvieną guaniną galima rasti tik citoziną. Taigi grandinės yra viena kitą papildančios, tai yra, nukleotidų seka vienoje grandinėje vienareikšmiškai lemia jų seką kitoje. Dvi grandinės eina priešingomis kryptimis, o jų galinės fosfatų grupės yra priešinguose dvigubos spiralės galuose.
Atlikdami savo tyrimus, 1953 metais Watsonas ir Crickas pasiūlė DNR molekulės struktūros modelį (3 pav.), kuris išlieka aktualus iki šių dienų. Pagal modelį DNR molekulė susideda iš dviejų vienas kitą papildančių polinukleotidų grandinių. Kiekviena DNR grandinė yra kelių dešimčių tūkstančių nukleotidų polinukleotidas. Jame gretimi nukleotidai sudaro taisyklingą pentozės-fosfato stuburą dėl fosforo rūgšties liekanos ir dezoksiribozės sujungimo stipriu kovalentiniu ryšiu. Šiuo atveju vienos polinukleotidinės grandinės azotinės bazės yra išdėstytos griežtai apibrėžta tvarka prieš kitos azotines bazes. Azoto bazių kaita polinukleotidų grandinėje yra nereguliari.
Azotinių bazių išsidėstymas DNR grandinėje yra komplementarus (iš graikiško „komplemento“ – papildymas), t.y. prieš adeniną (A) visada yra timinas (T), o prieš guaniną (G) tik citozinas (C). Taip yra dėl to, kad A ir T, taip pat G ir C griežtai atitinka vienas kitą, t.y. papildo vienas kitą. Šį atitikimą suteikia bazių cheminė struktūra, leidžianti susidaryti vandeniliniams ryšiams purino ir pirimidino poroje. Yra dvi jungtys tarp A ir T, trys tarp G ir C. Šie ryšiai suteikia dalinį DNR molekulės stabilizavimą erdvėje. Šiuo atveju dvigubos spiralės stabilumas yra tiesiogiai proporcingas G≡C jungčių skaičiui, kurios yra stabilesnės, palyginti su A = T jungtimis.
Žinoma nukleotidų išsidėstymo vienoje DNR grandinėje seka leidžia pagal komplementarumo principą nustatyti kitos grandinės nukleotidus.
Be to, buvo nustatyta, kad azoto bazės, turinčios aromatinę struktūrą vandeniniame tirpale, yra viena virš kitos, sudarydamos tarsi monetų šūsnį. Šis kaminų formavimo procesas organinės molekulės vadinamas stakingu. Nagrinėjamo Watson-Crick modelio DNR molekulės polinukleotidinės grandinės turi panašią fizikinę ir cheminę būseną, jų azotinės bazės yra monetų krūvos pavidalu, tarp kurių plokštumų atsiranda van der Waals sąveikos (stacking sąveikos).
Vandeniliniai ryšiai tarp komplementarių bazių (horizontaliai) ir polinukleotidinės grandinės bazių plokštumų sąveika dėl van der Waals jėgų (vertikaliai) suteikia DNR molekulei papildomą stabilizavimą erdvėje.
Abiejų grandinių cukraus ir fosfato stuburai yra nukreipti į išorę, o pagrindai į vidų, vienas į kitą. Grandinių kryptis DNR yra antilygiagreti (vienos iš jų kryptis 5 "-> 3", kitos - 3 "-> 5", ty 3 "vienos grandinės galas yra priešais 5" kitos grandinės galą .). Grandinės sudaro dešiniąsias spirales su bendra ašimi. Vienas spiralės apsisukimas yra 10 nukleotidų, ritės dydis yra 3,4 nm, kiekvieno nukleotido aukštis - 0,34 nm, o spiralės skersmuo - 2,0 nm. Dėl vienos grandinės sukimosi aplink kitą susidaro didelis DNR dvigubos spiralės griovelis (apie 20 Å skersmens) ir mažas griovelis (apie 12 Å). Ši Watson-Crick dvigubos spiralės forma vėliau buvo vadinama B forma. Ląstelėse DNR dažniausiai yra B formos, kuri yra stabiliausia.
DNR funkcijos
Siūlomas modelis paaiškino daugybę biologinių dezoksiribonukleorūgšties savybių, įskaitant genetinės informacijos saugojimą ir genų įvairovę, kurią teikia įvairios nuoseklios 4 nukleotidų kombinacijos, ir genetinio kodo egzistavimo faktą, galimybę savarankiškai daugintis. ir perduoti genetinę informaciją, gaunamą replikacijos procese, ir genetinės informacijos įgyvendinimą baltymų pavidalu, taip pat bet kokius kitus junginius, susidarančius fermentų baltymų pagalba.
Pagrindinės DNR funkcijos.
- DNR yra genetinės informacijos nešėja, kurią užtikrina genetinio kodo egzistavimo faktas.
- Genetinės informacijos dauginimas ir perdavimas ląstelių ir organizmų kartomis. Šią funkciją užtikrina replikacijos procesas.
- Genetinės informacijos realizavimas baltymų pavidalu, taip pat bet kokie kiti junginiai, susidarantys baltymų fermentų pagalba. Šią funkciją užtikrina transkripcijos ir vertimo procesai.

Dvigrandės DNR organizavimo formos
DNR gali sudaryti kelių tipų dvigubas spirales (4 pav.). Šiuo metu jau žinomos šešios formos (nuo A iki E ir Z formos).
Struktūrinės DNR formos, kaip nustatė Rosalind Franklin, priklauso nuo vandens prisotinimo nukleorūgšties molekulėje. Atliekant DNR skaidulų tyrimus naudojant rentgeno struktūrinę analizę, buvo įrodyta, kad rentgeno spindulių difrakcijos vaizdas radikaliai priklauso nuo to, prie kokio santykinio drėgnumo, kokio vandens prisotinimo laipsnio šio pluošto vyksta eksperimentas. Jei pluoštas buvo pakankamai prisotintas vandens, tada buvo gauta viena rentgeno nuotrauka. Džiovinant atsirado visiškai kitoks rentgeno vaizdas, labai skiriasi nuo didelės drėgmės pluošto rentgeno paveikslo.
Didelės drėgmės DNR molekulė vadinama B forma... Fiziologinėmis sąlygomis (maža druskos koncentracija, didelis hidratacijos laipsnis) dominuojantis struktūrinis DNR tipas yra B forma (pagrindinė dvigrandės DNR forma yra Watson-Crick modelis). Tokios molekulės spiralės žingsnis yra 3,4 nm. Viename posūkyje yra 10 papildomų porų susuktų „monetų“ – azoto bazių – pavidalu. Krūvos yra laikomos vandenilinėmis jungtimis tarp dviejų priešingų rietuvės „monetų“ ir yra „apvyniotos“ dviem fosfodiesterio skeleto juostelėmis, susuktomis į dešinę spiralę. Azotinių bazių plokštumos yra statmenos spiralės ašiai. Gretimos papildomos poros yra pasuktos viena kitos atžvilgiu 36 °. Sraigės skersmuo yra 20 Å, purino nukleotidas yra 12 Å, o pirimidino nukleotidas - 8 Å.
Mažesnės drėgmės DNR molekulė vadinama A forma... A forma susidaro mažiau hidratacijos sąlygomis ir esant didesniam Na + arba K + jonų kiekiui. Ši platesnė dešiniarankė turi 11 bazinių porų viename posūkyje. Azotinių bazių plokštumos yra labiau pasvirusios į spiralės ašį, jos nuo normalios iki spiralės ašies nukrypusios 20°. Taigi atsiranda vidinė tuštuma, kurios skersmuo yra 5 Å. Atstumas tarp gretimų nukleotidų yra 0,23 nm, ritės ilgis – 2,5 nm, spiralės skersmuo – 2,3 nm.
Iš pradžių buvo manoma, kad DNR A forma buvo mažiau svarbi. Tačiau vėliau paaiškėjo, kad DNR A forma, kaip ir B forma, turi didžiulę biologinę reikšmę. RNR-DNR spiralė šablono ir pradmenų komplekse turi A formą, taip pat RNR-RNR spiralę ir RNR plaukų segtuko struktūras (ribozės 2'-hidroksilo grupė neleidžia RNR molekulėms sudaryti B formos) . DNR A forma buvo rasta ginčų. Nustatyta, kad DNR A forma yra 10 kartų atsparesnė UV spinduliams nei B forma.
A formos ir B formos vadinamos kanoninės formos DNR.
Formos C-E taip pat dešiniarankiai, jų formavimąsi galima stebėti tik specialių eksperimentų metu ir, matyt, in vivo jų nėra. C formos DNR struktūra panaši į B-DNR. Bazinių porų skaičius viename posūkyje yra 9,33, spiralės ilgis yra 3,1 nm. Bazinės poros pasvirusios 8 laipsnių kampu statmenos padėties ašiai atžvilgiu. Grioveliai yra panašaus dydžio kaip B-DNR grioveliai. Šiuo atveju pagrindinis griovelis yra šiek tiek seklesnis, o mažasis griovelis yra gilesnis. Natūralūs ir sintetiniai DNR polinukleotidai gali pereiti į C formą.
| 1 lentelė. Kai kurių tipų DNR struktūrų charakteristikos | |||
| Spiralinis tipas | A | B | Z |
| Spiralinis žingsnis | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiralinis sukimas | Teisingai | Teisingai | Kairė |
| Bazinės poros vienam posūkiui | 11 | 10 | 12 |
| Atstumas tarp bazinių plokštumų | 0,256 nm | 0,338 nm | 0,371 nm |
| Glikozidinio ryšio konformacija | anti | anti | anti-C sin-g |
| Furanozės ciklo konformacija | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-c |
| Griovelio plotis mažas / didelis | 1,11 / 0,22 nm | 0,57 / 1,17 nm | 0,2 / 0,88 nm |
| Griovelio gylis, mažas / didelis | 0,26 / 1,30 nm | 0,82 / 0,85 nm | 1,38 / 0,37 nm |
| Spiralės skersmuo | 2,3 nm | 2,0 nm | 1,8 nm |
Struktūriniai DNR elementai
(nekanoninės DNR struktūros)
Struktūriniai DNR elementai apima neįprastas struktūras, kurias riboja kai kurios specialios sekos:
|

Z formos DNR buvo atrastas 1979 m., tiriant heksanukleotidą d (CG) 3 -. Jį atrado MIT profesorius Aleksandras Richas ir jo kolegos. Z forma tapo vienu iš svarbiausių struktūrinių DNR elementų dėl to, kad jos formavimasis buvo stebimas DNR srityse, kur purinai kaitaliojasi su pirimidinais (pavyzdžiui, 5'-HCGCH-3'), arba 5' pasikartojimuose. -CHCH-3', turintis metilinto citozino. Esminė Z-DNR susidarymo ir stabilizavimo sąlyga buvo purino nukleotidų buvimas sin-konformacijoje, pakaitomis su pirimidino bazėmis antikonformacijoje.
Natūralios DNR molekulės paprastai egzistuoja tinkamos B formos, jei jose nėra (CH) n tipo sekų. Tačiau jei tokios sekos yra įtrauktos į DNR, tai šios sritys, pasikeitus tirpalo joniniam stiprumui ar katijonams, neutralizuojantiems neigiamą krūvį ant fosfodiesterio pagrindo, gali transformuotis į Z formą, o kitos grandinės DNR sritys išlieka klasikinė B forma. Tokio perėjimo galimybė rodo, kad dvi DNR dvigubos spiralės grandinės yra dinamiškos būsenos ir gali išsivynioti viena kitos atžvilgiu, pereidamos iš dešinės formos į kairę ir atvirkščiai. Tokio labilumo, leidžiančio DNR struktūros konformacines transformacijas, biologinės pasekmės dar nėra visiškai suprantamos. Manoma, kad Z-DNR sritys vaidina tam tikrą vaidmenį reguliuojant kai kurių genų ekspresiją ir dalyvauja genetinėje rekombinacijoje.
DNR Z forma yra kairioji dviguba spiralė, kurioje fosfodiesterio stuburas yra zigzago būdu išilgai molekulės ašies. Iš čia ir kilo molekulės pavadinimas (zigzagas) -DHK. Z-DNR yra mažiausiai susisukusi (12 bazinių porų viename posūkyje) ir ploniausia žinoma gamtoje. Atstumas tarp gretimų nukleotidų yra 0,38 nm, ritės ilgis – 4,56 nm, o Z-DNR skersmuo – 1,8 nm. Be to, išvaizdaši DNR molekulė išsiskiria tuo, kad yra vienas griovelis.
DNR Z forma buvo rasta prokariotinėse ir eukariotinėse ląstelėse. Šiuo metu yra gauti antikūnai, galintys atskirti DNR Z formą nuo B formos. Šie antikūnai jungiasi prie specifinių Drosophila (Dr. melanogaster) seilių liaukų ląstelių milžiniškų chromosomų sričių. Ryšio reakciją lengva sekti dėl neįprastos šių chromosomų struktūros, kai tankesnės sritys (diskai) kontrastuoja su mažiau tankiomis sritimis (tarpdiskiais). Z-DNR regionai yra tarpjuostose. Iš to išplaukia, kad Z forma iš tikrųjų egzistuoja natūraliomis sąlygomis, nors atskirų Z formos atkarpų dydžiai vis dar nežinomi.

(shifters) yra labiausiai žinomos ir dažniausiai randamos bazinės sekos DNR. Palindromas yra žodis ar frazė, skaitoma iš kairės į dešinę ir atvirkščiai tuo pačiu būdu. Tokių žodžių ar frazių pavyzdžiai: SHALASH, KAZAK, POTOP, IR ROSE FALLED ANT AZOR'S Paw. Taikant DNR sritims, šis terminas (palindromas) reiškia tą patį nukleotidų kaitą grandinėje iš dešinės į kairę ir iš kairės į dešinę (kaip ir raidės žodyje "namelis" ir kt.).
Palindromui būdingi apverstų bazinių sekų pasikartojimai, turintys antros eilės simetriją dviejų DNR grandžių atžvilgiu. Tokios sekos dėl visiškai suprantamos priežasties yra viena kitą papildančios ir linkusios formuoti plaukų segtines arba kryžmines struktūras (pav.). Plaukų segtukai padeda reguliuojantiems baltymams atpažinti vietą, kur nurašytas genetinis chromosomos DNR tekstas.
Tais atvejais, kai toje pačioje DNR grandinėje yra apverstas pasikartojimas, ši seka vadinama veidrodiniu pakartojimu. Veidrodiniai pasikartojimai neturi savęs papildymo savybių, todėl negali sudaryti plaukų segtukų ar kryžminių struktūrų. Tokio tipo sekos randamos praktiškai visose didelėse DNR molekulėse ir gali svyruoti nuo vos kelių bazinių porų iki kelių tūkstančių bazinių porų.
Palindromų buvimas kryžminių struktūrų pavidalu eukariotinėse ląstelėse nebuvo įrodytas, nors E. coli ląstelėse in vivo buvo rasta nemažai kryžminių struktūrų. Savarankiškai papildančių sekų buvimas RNR arba viengrandėje DNR yra pagrindinė priežastis, dėl kurios nukleino grandinė tirpaluose susilanksto į tam tikrą erdvinę struktūrą, kuriai būdingas daugybės „plaukų segtukų“ susidarymas.

H formos DNR yra trijų DNR grandžių sudaryta spiralė – DNR triguba spiralė. Tai yra Watson-Crick dvigubos spiralės kompleksas su trečiąja viengrande DNR grandine, kuri telpa į didelį jos griovelį, susidarius vadinamajai Hoogsteen porai.
Toks tripleksas susidaro dėl DNR dvigubos spiralės sulankstymo taip, kad pusė jos atkarpos lieka dvigubos spiralės pavidalu, o kita pusė yra atjungta. Šiuo atveju viena iš atjungtų spiralių suformuoja naują struktūrą su pirmąja dvigubos spiralės puse - triguba spirale, o antroji pasirodo esanti nestruktūrizuota, vienos grandinės sekcijos pavidalu. Šio struktūrinio perėjimo ypatybė – ryški priklausomybė nuo terpės pH, kurios protonai stabilizuoja naują struktūrą. Dėl šios savybės nauja struktūra buvo pavadinta DNR H forma, kurios susidarymas buvo nustatytas superspiralinėse plazmidėse, turinčiose homopurino-homopirimidino sritis, kurios yra veidrodinis pasikartojimas.
Atliekant tolesnius tyrimus, kai kurių homopurino-homopirimidino dvigrandžių polinukleotidų struktūrinio perėjimo galimybė susidarant trijų grandžių struktūrai, kurią sudaro:
- viena homopurino ir dvi homopirimidino gijos ( Py-Pu-Py triplex) [Hoogsteen sąveika].
Py-Pu-Py triplekso sudedamosios dalys yra kanoninės triados, izomorfinės CGC + ir TAT. Triplekso stabilizavimui reikalingas CGC + triados protonavimas, todėl šie tripleksai priklauso nuo tirpalo pH.
- viena homopirimidino ir dvi homopurino gijos ( Py-Pu-Pu triplex) [atvirkštinė Hoogsteen sąveika].
Py-Pu-Pu triplekso sudedamosios dalys yra kanoninės izomorfinės CGG ir TAA triadoms. Esminė Py-Pu-Pu tripleksų savybė yra jų stabilumo priklausomybė nuo dvigubo krūvio jonų buvimo, o skirtingų sekų tripleksams stabilizuoti reikalingi skirtingi jonai. Kadangi Py-Pu-Pu tripleksams susidaryti nereikia protonuoti juos sudarančių nukleotidų, tokie tripleksai gali egzistuoti esant neutraliam pH.
Pastaba: tiesioginė ir atvirkštinė Hoogsteen sąveika paaiškinama 1-metiltimino simetrija: 180 ° sukimas lemia tai, kad O4 atomas pakeičiamas O2 atomu, o vandenilio jungčių sistema išsaugoma.
Yra dviejų tipų trigubos spiralės:
- lygiagrečios trigubos spiralės, kuriose trečiosios grandinės poliškumas sutampa su Watson-Crick duplekso homopurino grandinės poliškumu
- antiparalelinės trigubos spiralės, kuriose trečiosios ir homopurino grandinių poliškumas yra priešingas.

G-quadruplex- 4 grandžių DNR. Tokia struktūra susidaro, jei yra keturi guaninai, kurie sudaro vadinamąjį G-kvadrupleksą – apvalų keturių guaninų šokį.
Pirmosios užuominos apie tokių struktūrų susidarymo galimybę buvo gautos dar gerokai prieš Watsono ir Cricko proveržį – dar 1910 m. Tada vokiečių chemikas Ivaras Bangas išsiaiškino, kad vienas iš DNR komponentų – guanozės rūgštis – didelėmis koncentracijomis formuoja gelius, o kiti DNR komponentai šios savybės neturi.
1962 m., naudojant rentgeno spindulių difrakcijos metodą, pavyko nustatyti šio gelio ląstelių struktūrą. Paaiškėjo, kad jį sudaro keturios guanino liekanos, jungiančios viena kitą ratu ir sudarančios būdingą kvadratą. Centre jungtis palaiko metalo jonas (Na, K, Mg). Tokios pat struktūros gali susidaryti ir DNR, jei joje yra daug guanino. Šie plokšti kvadratai (G-kvartetai) yra sukrauti, kad sudarytų gana stabilias, tankias struktūras (G-kvadrupleksus).
Keturios atskiros DNR grandinės gali būti susipynusios į keturių grandžių kompleksus, tačiau tai veikiau išimtis. Dažniau viena nukleino rūgšties grandinė tiesiog surišama į mazgą, suformuojant būdingus sustorėjimus (pavyzdžiui, chromosomų galuose), arba dvigrandė DNR suformuoja vietinį kvadrupleksą kuriame nors guanino turtingame regione.
Labiausiai ištirtas yra kvadrupleksų buvimas chromosomų galuose – telomeruose ir onkopromotoriuose. Tačiau iki šiol pilnas supratimas apie tokios DNR lokalizaciją žmogaus chromosomose nėra žinomas.
Visos šios neįprastos linijinės formos DNR struktūros yra nestabilios, palyginti su DNR B forma. Tačiau DNR dažnai egzistuoja apskrito topologinio streso pavidalu, kai joje yra vadinamasis superspiritas. Tokiomis sąlygomis nesunkiai susidaro nekanoninės DNR struktūros: Z formos, kryžiai ir plaukų segtukai, H formos, guanino kvadrupleksai ir i motyvas.
- Superspiralinė forma – pastebima, kai ji išskiriama iš ląstelės branduolio nepažeidžiant pentozės-fosfato stuburo. Jis turi labai susuktų uždarų žiedų formą. Superspiralėje DNR dviguba spiralė yra bent kartą „susisukta ant savęs“, tai yra, joje yra bent viena superspiralė (aštuonetuko pavidalu).
- Atsipalaidavusi DNR būsena stebima su viena pertrauka (vienos grandinės nutrūkimu). Tokiu atveju superspiralės išnyksta ir DNR įgauna uždaro žiedo formą.
- Linijinė DNR forma – stebima, kai nutrūksta dvi dvigubos spiralės gijos.
DNR tretinė struktūra
DNR tretinė struktūra susidaro dėl papildomo dvigrandės molekulės susisukimo erdvėje – jos superspiralizavimo. DNR molekulės superspiralizacija eukariotinėse ląstelėse, priešingai nei prokariotuose, atliekama kompleksų su baltymais pavidalu.
Beveik visa eukariotinė DNR randama branduolių chromosomose, tik nedidelis jos kiekis yra mitochondrijose, augaluose ir plastidėse. Pagrindinė eukariotinių ląstelių (įskaitant žmogaus chromosomas) chromosomų medžiaga yra chromatinas, susidedantis iš dvigrandžių DNR, histono ir nehistoninių baltymų.
Chromatino histono baltymai
Histonai yra paprasti baltymai, kurie sudaro iki 50% chromatino. Visose tirtose gyvūnų ir augalų ląstelėse buvo aptiktos penkios pagrindinės histonų klasės: H1, H2A, H2B, H3, H4, kurios skiriasi dydžiu, aminorūgščių sudėtimi ir krūvio reikšme (visada teigiama).
Žinduolių histonas H1 susideda iš vienos polipeptidinės grandinės, susidedančios iš maždaug 215 aminorūgščių; kitų histonų dydžiai svyruoja nuo 100 iki 135 aminorūgščių. Visi jie yra spiralizuoti ir susukti į maždaug 2,5 nm skersmens rutuliuką, turi neįprastai daug teigiamo krūvio aminorūgščių lizino ir arginino. Histonai gali būti acetilinti, metilinti, fosforilinti, poli (ADP) -ribozilinti, o histonai H2A ir H2B yra kovalentiškai susieti su ubikvitinu. Koks yra tokių modifikacijų vaidmuo formuojant struktūrą ir atliekant histonų funkcijas, dar nėra iki galo išaiškinta. Daroma prielaida, kad tai yra jų gebėjimas sąveikauti su DNR ir užtikrinti vieną iš genų veikimo reguliavimo mechanizmų.
Histonai sąveikauja su DNR daugiausia per joninius ryšius (druskų tiltelius), susidariusius tarp neigiamą krūvį turinčių DNR fosfatų grupių ir teigiamai įkrautų histonų lizino bei arginino liekanų.
Ne histoniniai chromatino baltymai
Skirtingai nuo histonų, nehistoniniai baltymai yra labai įvairūs. Išskirta iki 590 skirtingų DNR surišančių nehistoninių baltymų frakcijų. Jie taip pat vadinami rūgštiniais baltymais, nes jų struktūroje vyrauja rūgštinės aminorūgštys (tai yra polianijonai). Specifinis chromatino aktyvumo reguliavimas yra susijęs su įvairiais ne histono baltymais. Pavyzdžiui, fermentai, reikalingi DNR replikacijai ir ekspresijai, gali laikinai prisijungti prie chromatino. Kiti baltymai, pavyzdžiui, dalyvaujantys įvairiuose reguliavimo procesuose, prie DNR jungiasi tik specifiniuose audiniuose arba tam tikrose diferenciacijos stadijose. Kiekvienas baltymas yra papildomas tam tikrai DNR nukleotidų sekai (DNR vietai). Į šią grupę įeina:
- vietos specifinių cinko pirštų baltymų šeima. Kiekvienas cinko pirštas atpažįsta konkrečią vietą, susidedančią iš 5 nukleotidų porų.
- vietai būdingų baltymų šeima – homodimerai. Tokio baltymo fragmentas, besiliečiantis su DNR, turi spiralės posūkio-spiralės struktūrą.
- didelio mobilumo gelio baltymai (HMG baltymai) yra struktūrinių ir reguliuojančių baltymų grupė, kuri nuolat yra susijusi su chromatinu. Jų molekulinė masė mažesnė nei 30 kDa ir pasižymi dideliu įkrautų aminorūgščių kiekiu. Dėl mažos molekulinės masės HMG baltymai yra labai mobilūs poliakrilamido gelio elektroforezės metu.
- replikacijos, transkripcijos ir taisymo fermentai.
Dalyvaujant struktūriniams, reguliuojantiems baltymams ir fermentams, dalyvaujantiems DNR ir RNR sintezėje, nukleozomų grandinė paverčiama labai kondensuotu baltymų ir DNR kompleksu. Gauta struktūra yra 10 000 kartų trumpesnė už pradinę DNR molekulę.

Chromatinas
Chromatinas yra baltymų kompleksas su branduoline DNR ir neorganinių medžiagų... Didžioji chromatino dalis yra neaktyvi. Jame yra sandariai supakuota, kondensuota DNR. Tai heterochromatinas. Atskirkite konstitucinį, genetiškai neaktyvų chromatiną (palydovinę DNR), susidedantį iš neišreikštų regionų, ir pasirenkamą - neaktyvų keliose kartose, bet tam tikromis aplinkybėmis galintį išreikšti.
Aktyvusis chromatinas (euchromatinas) yra nekondensuotas, t.y. supakuotas ne taip sandariai. Skirtingose ląstelėse jo kiekis svyruoja nuo 2 iki 11%. Smegenų ląstelėse jo daugiausia - 10-11%, kepenų ląstelėse - 3-4 ir inkstų ląstelėse - 2-3%. Pastebėta aktyvi euchromatino transkripcija. Tuo pačiu metu jo struktūrinė struktūra leidžia specializuotose ląstelėse įvairiais būdais panaudoti vieną ir tą pačią DNR genetinę informaciją, būdingą tam tikram organizmo tipui.
Elektroniniame mikroskope chromatino vaizdas primena karoliuką: maždaug 10 nm dydžio sferiniai sustorėjimai, atskirti siūliniais tilteliais. Šie rutuliniai sustorėjimai vadinami nukleozomomis. Nukleosoma yra struktūrinis chromatino vienetas. Kiekvienoje nukleozomoje yra superspiralinis DNR segmentas, kurio ilgis yra 146 bazių poros, suvyniotas 1,75 kairiojo posūkio nukleosomos šerdyje. Nukleosomų šerdis yra histono oktameras, susidedantis iš histonų H2A, H2B, H3 ir H4, dviejų kiekvieno tipo molekulių (9 pav.), kuris atrodo kaip 11 nm skersmens ir 5,7 nm storio diskas. Penktasis histonas H1 nėra nukleosominės šerdies dalis ir nedalyvauja DNR vyniojimosi procese ant histono oktamero. Jis susisiekia su DNR ten, kur dviguba spiralė patenka ir išeina iš nukleosominės šerdies. Tai tarpžievės (linkerio) DNR sritys, kurių ilgis priklauso nuo ląstelės tipo nuo 40 iki 50 bazinių porų. Dėl to kinta ir DNR fragmento, įtraukto į nukleozomą, ilgis (nuo 186 iki 196 nukleotidų porų).
Nukleosomoje yra apie 90% DNR, likusi dalis yra jungtis. Manoma, kad nukleosomos yra „tylaus“ chromatino fragmentai, o jungtis yra aktyvi. Tačiau nukleosomos gali išsiskleisti ir tapti linijinės. Išsiskleidusios nukleosomos jau yra aktyvus chromatinas. Taip aiškiai pasireiškia funkcijos priklausomybė nuo struktūros. Galima daryti prielaidą, kad kuo daugiau chromatino yra rutulinių nukleozomų sudėtyje, tuo jis mažiau aktyvus. Akivaizdu, kad skirtingose ląstelėse nevienoda ramybės būsenos chromatino dalis yra susijusi su tokių nukleozomų skaičiumi.
Elektroninėse mikroskopinėse nuotraukose, priklausomai nuo izoliacijos sąlygų ir tempimo laipsnio, chromatinas gali atrodyti ne tik kaip ilgas siūlas su sustorėjimais – nukleozomų „karoliukais“, bet ir kaip trumpesnė ir tankesnė fibrilė (pluoštas), kurio skersmuo 30 nm, kurio susidarymas stebimas sąveikaujant histonui H1, susietam su DNR ir histono H3 jungiklio sritimi, dėl ko papildomai pasisuka šešių nukleozomų spiralė kiekviename posūkyje ir susidaro 30 nm skersmens solenoidas. Šiuo atveju histono baltymas gali trukdyti daugelio genų transkripcijai ir taip reguliuoti jų veiklą.
Dėl aukščiau aprašytų DNR sąveikų su histonais DNR dvigubos spiralės segmentas, sudarytas iš 186 bazių porų, kurių vidutinis skersmuo yra 2 nm, o ilgis 57 nm, virsta spirale, kurios skersmuo 10 nm. ilgis 5 nm. Vėliau suspaudus šią spiralę iki 30 nm skersmens pluošto, kondensacijos laipsnis padidėja šešis kartus.
Galiausiai supakavus DNR dupleksą su penkiais histonais, DNR kondensuojasi 50 kartų. Tačiau net ir toks didelis kondensacijos laipsnis negali paaiškinti beveik 50 000 – 100 000 kartų DNR tankėjimo metafazėje chromosomoje. Deja, tolimesnio chromatino pakavimo iki metafazės chromosomos detalės dar nėra žinomos, todėl galima atsižvelgti tik į bendrus šio proceso bruožus.
DNR sutankinimo lygiai chromosomose
Kiekviena DNR molekulė yra supakuota į atskirą chromosomą. Žmogaus diploidinėse ląstelėse yra 46 chromosomos, kurios yra ląstelės branduolyje. Bendras visų ląstelės chromosomų DNR ilgis yra 1,74 m, tačiau branduolio, kuriame supakuotos chromosomos, skersmuo yra milijonus kartų mažesnis. Tokį kompaktišką DNR susikaupimą chromosomose ir chromosomose ląstelės branduolyje užtikrina įvairūs histoniniai ir nehistoniniai baltymai, kurie tam tikra seka sąveikauja su DNR (žr. aukščiau). DNR sutankinimas chromosomose leidžia sumažinti jos linijinius matmenis maždaug 10 000 kartų – įprastai nuo 5 cm iki 5 mikronų. Yra keli sutankinimo lygiai (10 pav.).


- DNR dviguba spiralė yra neigiamo krūvio molekulė, kurios skersmuo yra 2 nm, o ilgis - keli cm.
- nukleosominis lygis– chromatinas elektroniniame mikroskope atrodo kaip „karoliukų“ – nukleozomų – „ant stygos“ grandinėlė. Nukleosoma yra universalus struktūrinis vienetas, randamas tiek euchromatine, tiek heterochromatine, tarpfaziniame branduolyje ir metafazės chromosomose.
Nukleosominį tankinimo lygį užtikrina specialūs baltymai – histonai. Aštuoni teigiamai įkrauti histono domenai sudaro nukleosomos šerdį (šerdį), aplink kurią suvyniota neigiamo krūvio DNR molekulė. Dėl to sutrumpėja 7 kartus, o skersmuo padidėja nuo 2 iki 11 nm.
- solenoido lygis
Solenoidiniam chromosomų organizavimo lygiui būdingas nukleosominio siūlelio susisukimas ir iš jo formuojasi storesnės 20-35 nm skersmens fibrilės - solenoidai arba superbidai. Solenoido žingsnis yra 11 nm, viename posūkyje yra apie 6-10 nukleozomų. Solenoidinis įpakavimas laikomas labiau tikėtinu nei superbid, pagal kurį 20–35 nm skersmens chromatino fibrilė yra granulių arba superbidų grandinė, kurių kiekviena susideda iš aštuonių nukleozomų. Solenoido lygyje linijinis DNR dydis sumažėja 6-10 kartų, skersmuo padidėja iki 30 nm.
- kilpos lygis
Kilpų lygį užtikrina ne histono vietai specifiniai DNR surišantys baltymai, kurie atpažįsta specifines DNR sekas ir prie jų prisijungia, sudarydami maždaug 30–300 kb kilpas. Kilpa suteikia genų ekspresiją, ty E. kilpa yra ne tik struktūrinis, bet ir funkcinis darinys. Šiame lygyje sutrumpėja 20-30 kartų. Skersmuo padidėja iki 300 nm. Ant citologinių preparatų galima pastebėti į kilpą panašias „lempos šepetėlio“ tipo struktūras varliagyvių oocituose. Šios kilpos yra akivaizdžiai susuktos ir atstovauja DNR domenams, kurie tikriausiai atitinka chromatino transkripcijos ir replikacijos vienetus. Specifiniai baltymai fiksuoja kilpų pagrindus ir, galbūt, kai kuriuos jų vidinius regionus. Kilpos formos domeno organizacija skatina chromatino sulankstymą metafazės chromosomose į aukštesnės eilės spiralines struktūras.
- domeno lygiu
Chromosomų organizavimo srities lygis buvo nepakankamai ištirtas. Šiame lygyje pastebimas kilpų domenų susidarymas - 25-30 nm storio siūlų (fibrilių), kuriose yra 60% baltymų, 35% DNR ir 5% RNR, struktūros praktiškai nematomos visose ląstelės fazėse. ciklas, išskyrus mitozę, ir yra šiek tiek atsitiktinai pasiskirstę ląstelės branduolyje. Ant citologinių preparatų galima pastebėti į kilpą panašias „lempos šepetėlio“ tipo struktūras varliagyvių oocituose.
Kilpų domenai prijungia savo bazes prie intrabranduolinio baltymo matricos vadinamosiose įterptosiose prisijungimo vietose, dažnai vadinamose MAR / SAR sekomis (MAR, iš angliško matricos asocijuoto regiono; SAR, iš angliško karkaso prijungimo regionų) - DNR fragmentai keli. šimto ilgio bazių porų, kurioms būdingas didelis kiekis (> 65%) A/T bazių porų. Atrodo, kad kiekvienas domenas turi vieną replikacijos pradžią ir veikia kaip savarankiškas superspiralinis vienetas. Bet kuriame kilpiniame domene yra daug transkripcijos vienetų, kurių veikimas tikriausiai yra koordinuotas – visas domenas yra aktyvios arba neaktyvios būsenos.
Domeno lygiu dėl nuoseklaus chromatino pakavimo linijiniai DNR matmenys sumažėja maždaug 200 kartų (700 nm).
- chromosomų lygis
Chromosomų lygyje profazės chromosomos kondensacija į metafazę įvyksta sutankinant kilpinius domenus aplink ašinį ne histoninių baltymų karkasą. Šį superspiralizavimą lydi visų ląstelėje esančių H1 molekulių fosforilinimas. Dėl to metafazinė chromosoma gali būti pavaizduota kaip sandariai supakuotos solenoidinės kilpos, susuktos į tankią spiralę. Įprastoje žmogaus chromosomoje gali būti iki 2600 kilpų. Tokios struktūros storis siekia 1400 nm (dvi chromatidės), o DNR molekulė sutrumpėja 104 kartus, t.y. su 5 cm ištemptos DNR iki 5 μm.
Chromosomų funkcijos
Sąveikaujant su ekstrachromosominiais mechanizmais, chromosomos suteikia
- paveldimos informacijos saugojimas
- naudojant šią informaciją korinio ryšio organizacijai kurti ir palaikyti
- paveldimos informacijos skaitymo reguliavimas
- savaiminis genetinės medžiagos padvigubėjimas
- genetinės medžiagos perkėlimas iš motininės ląstelės dukrai.
Yra duomenų, kad aktyvavus chromatino sritį, t.y. transkripcijos metu iš jo pirmiausia grįžtamai pašalinamas histonas H1, o paskui histono oktetas. Tai sukelia chromatino dekondensaciją, nuoseklų 30 nanometrų chromatino fibrilės perėjimą į 10 nanometrų siūlą ir tolesnį jo išsiskleidimą į laisvos DNR sritis, t.y. nukleosominės struktūros praradimas.
Visi žinome, kad žmogaus išvaizda, kai kurie įpročiai ir net ligos yra paveldimi. Visa ši informacija apie gyvą būtybę yra užkoduota genuose. Taigi, kaip atrodo šie patarlių genai, kaip jie veikia ir kur jie yra?
Taigi, visų bet kurio žmogaus ar gyvūno genų nešėjas yra DNR. Šį junginį 1869 metais atrado Johanas Friedrichas Miescheris. Chemiškai DNR yra dezoksiribonukleino rūgštis. Ką tai reiškia? Kaip ši rūgštis turi visos mūsų planetos gyvybės genetinį kodą?
Pradėkime nuo to, kur yra DNR. Žmogaus ląstelėje yra daug organelių, atliekančių įvairias funkcijas. DNR yra branduolyje. Branduolys yra maža organelė, kurią supa speciali membrana, kurioje saugoma visa genetinė medžiaga – DNR.
Kokia yra DNR molekulės struktūra?
Pirmiausia pažiūrėkime, kas yra DNR. DNR yra labai ilga molekulė, sudaryta iš statybinių blokų – nukleotidų. Yra 4 nukleotidų tipai – adeninas (A), timinas (T), guaninas (G) ir citozinas (C). Nukleotidų grandinė schematiškai atrodo taip: GGAATCTAAG... Tai nukleotidų seka, kuri yra DNR grandinė.Pirmą kartą DNR struktūrą 1953 metais iššifravo Jamesas Watsonas ir Francisas Crickas.
Vienoje DNR molekulėje yra dvi nukleotidų grandinės, kurios spirališkai susuktos viena aplink kitą. Kaip šios nukleotidų grandinės sulimpa ir susisuka į spiralę? Šis reiškinys atsiranda dėl komplementarumo savybės. Komplementarumas reiškia, kad tik tam tikri nukleotidai (komplementarūs) gali būti išdėstyti vienas priešais kitą dviejose grandinėse. Taigi, priešingai nei adeninas, visada yra timinas, o priešingai guaninui visada yra tik citozinas. Taigi guaninas yra komplementarus citozinui, o adeninas – timinui.Tokios viena kitai priešingos nukleotidų poros skirtingose grandinėse dar vadinamos komplementariomis.
 Jį galima schematiškai pavaizduoti taip:
Jį galima schematiškai pavaizduoti taip:
G-C
T-A
T-A
C-G
Šios papildomos poros A - T ir G - C susidaro cheminis ryšys tarp poros nukleotidų, o ryšys tarp G ir C yra stipresnis nei tarp A ir T. Ryšys susidaro griežtai tarp komplementarių bazių, tai yra, ryšio tarp nekomplementarių G ir A susidarymas neįmanomas.
DNR pakuotė, kaip DNR grandinė tampa chromosoma?
Kodėl šios DNR nukleotidų grandinės taip pat sukasi viena aplink kitą? Kam to reikia? Faktas yra tas, kad nukleotidų skaičius yra didžiulis ir reikia daug vietos, kad tilptų tokias ilgas grandines. Dėl šios priežasties dvi DNR grandinės sukasi spirale aplink kitą. Šis reiškinys vadinama spiralizacija. Dėl spiralizacijos DNR grandinės sutrumpėja 5-6 kartus.Kai kurias DNR molekules organizmas naudoja aktyviai, o kitas retai. Tokios retai naudojamos DNR molekulės, be spiralizacijos, patiria dar kompaktiškesnį „pakavimą“. Šis kompaktiškas paketas vadinamas superspiralizavimu ir sutrumpina DNR grandinę 25-30 kartų!
Kaip vyksta DNR grandinės pakavimas?
 Superspiralizavimui naudojami histono baltymai, kurie turi strypo arba sriegio ritės išvaizdą ir struktūrą. Ant šių „ritių“ – histono baltymų – suvyniotos spiralizuotos DNR grandinės. Taigi ilgas siūlas tampa labai kompaktiškai supakuotas ir užima labai mažai vietos.
Superspiralizavimui naudojami histono baltymai, kurie turi strypo arba sriegio ritės išvaizdą ir struktūrą. Ant šių „ritių“ – histono baltymų – suvyniotos spiralizuotos DNR grandinės. Taigi ilgas siūlas tampa labai kompaktiškai supakuotas ir užima labai mažai vietos. Jei reikia panaudoti tą ar kitą DNR molekulę, vyksta „išvyniojimo“ procesas, tai yra „išvyniojama“ iš „ritės“ – histono baltymo (jei jis buvo ant jos suvyniotas) ir išvyniojama DNR grandinė. spiralė į dvi lygiagrečias grandines. O kai DNR molekulė yra tokioje nesusuktoje būsenoje, tuomet iš jos galima nuskaityti reikiamą genetinę informaciją. Be to, genetinė informacija nuskaitoma tik iš nesusuktų DNR gijų!
Superspiralinių chromosomų rinkinys vadinamas heterochromatinas, ir chromosomos, prieinamos informacijai skaityti - euchromatinas.
Kas yra genai, koks jų ryšys su DNR?
Dabar pažiūrėkime, kas yra genai. Yra žinoma, kad yra genų, kurie lemia kraujo grupę, akių spalvą, plaukus, odą ir daugybę kitų mūsų organizmo savybių. Genas yra griežtai apibrėžta DNR dalis, susidedanti iš tam tikro skaičiaus nukleotidų, esančių griežtai apibrėžtoje kombinacijoje. Vieta griežtai apibrėžtoje DNR srityje reiškia, kad tam tikram genui buvo priskirta vieta, ir šios vietos pakeisti neįmanoma. Tikslinga atlikti tokį palyginimą: žmogus gyvena tam tikroje gatvėje, tam tikrame name ir bute, o žmogus negali savavališkai persikelti į kitą namą, butą ar į kitą gatvę. Tam tikras nukleotidų skaičius gene reiškia, kad kiekvienas genas turi tam tikrą nukleotidų skaičių ir negali tapti daugiau ar mažiau. Pavyzdžiui, insulino gamybos genas yra 60 bazinių porų ilgio; genas, koduojantis hormono oksitocino gamybą – 370 bazinių porų. Griežta nukleotidų seka yra unikali kiekvienam genui ir yra griežtai apibrėžta. Pavyzdžiui, AATTAATA seka yra geno, koduojančio insulino gamybą, fragmentas. Norint gauti insuliną, naudojama būtent tokia seka, o, pavyzdžiui, adrenalinui gauti, naudojamas kitoks nukleotidų derinys. Svarbu suprasti, kad tik tam tikra nukleotidų kombinacija koduoja tam tikrą „produktą“ (adrenaliną, insuliną ir kt.). Toks yra unikalus tam tikro skaičiaus nukleotidų derinys, stovintis „savo vietoje“ – tai yra genas.
Be genų, DNR grandinėje yra ir vadinamosios „nekoduojančios sekos“. Tokios nekoduojančios nukleotidų sekos reguliuoja genų darbą, padeda spiralizuotis chromosomoms, žymi geno pradžią ir pabaigą. Tačiau iki šiol daugumos nekoduojančių sekų vaidmuo lieka neaiškus.
Kas yra chromosoma? Lytinės chromosomos
Individo genų rinkinys vadinamas genomu. Natūralu, kad neįmanoma sutalpinti viso genomo į vieną DNR. Genomas yra suskaidytas į 46 poras DNR molekulių. Viena DNR molekulių pora vadinama chromosoma. Taigi būtent šias chromosomas žmogus turi 46 vienetus. Kiekviena chromosoma turi griežtai apibrėžtą genų rinkinį, pavyzdžiui, 18 chromosomoje yra genų, koduojančių akių spalvą ir pan. Chromosomos skiriasi viena nuo kitos ilgiu ir forma. Dažniausiai pasitaikančios formos yra X arba Y, tačiau yra ir kitų. Žmogus turi dvi tos pačios formos chromosomas, kurios vadinamos poromis (poromis). Dėl tokių skirtumų visos suporuotos chromosomos yra sunumeruotos – jų yra 23 poros. Tai reiškia, kad yra chromosomų pora # 1, pora # 2, # 3 ir kt. Kiekvienas genas, atsakingas už tam tikrą požymį, yra toje pačioje chromosomoje. Šiuolaikinėse rekomendacijose specialistams geno lokalizaciją galima nurodyti, pavyzdžiui, taip: 22 chromosoma, ilga ranka.Kuo skiriasi chromosomos?
 Kuo dar skiriasi chromosomos? Ką reiškia žodis ilgas petys? Paimkite X formos chromosomas. DNR grandinių susikirtimas gali vykti griežtai viduryje (X), arba gali būti ir ne centre. Kai toks DNR grandžių susikirtimas nevyksta centralizuotai, tada sankryžos taško atžvilgiu vieni galai yra ilgesni, kiti – atitinkamai trumpesni. Tokie ilgi galai dažniausiai vadinami ilga chromosomos ranka, o trumpi atitinkamai trumpąja ranka. Y formos chromosomose daugumą jų užima ilgi pečiai, o trumpi – labai maži (schemame paveikslėlyje jie net nenurodyti).
Kuo dar skiriasi chromosomos? Ką reiškia žodis ilgas petys? Paimkite X formos chromosomas. DNR grandinių susikirtimas gali vykti griežtai viduryje (X), arba gali būti ir ne centre. Kai toks DNR grandžių susikirtimas nevyksta centralizuotai, tada sankryžos taško atžvilgiu vieni galai yra ilgesni, kiti – atitinkamai trumpesni. Tokie ilgi galai dažniausiai vadinami ilga chromosomos ranka, o trumpi atitinkamai trumpąja ranka. Y formos chromosomose daugumą jų užima ilgi pečiai, o trumpi – labai maži (schemame paveikslėlyje jie net nenurodyti). Chromosomų dydis skiriasi: didžiausios yra 1 ir 3 porų chromosomos, mažiausios yra 17, 19 porų chromosomos.
Be formos ir dydžio, chromosomos skiriasi ir savo funkcijomis. Iš 23 porų 22 yra somatinės, o 1 – seksualios. Ką tai reiškia? Somatinės chromosomos lemia visus išorinius individo požymius, ypač jo elgesio reakcijos, paveldimas psichotipas, tai yra visi kiekvieno individualaus žmogaus bruožai ir savybės. Lytinių chromosomų pora lemia žmogaus lytį: vyro ar moters. Yra dviejų tipų žmogaus lytinės chromosomos – X (X) ir Y (Y). Jei jie yra sujungti kaip XX (X - X) - tai yra moteris, o jei XY (X - Y) - priešais mus yra vyras.
Paveldimos ligos ir chromosomų pažeidimai
Tačiau genomo „skilimų“ pasitaiko, tada žmonėms nustatomos genetinės ligos. Pavyzdžiui, kai 21 chromosomų poroje vietoj dviejų yra trys chromosomos, žmogus gimsta su Dauno sindromu.Yra daug mažesnių genetinės medžiagos „skilimų“, kurie nelemia ligos atsiradimo, o, priešingai, suteikia gerų savybių. Visi genetinės medžiagos „skilimai“ vadinami mutacijomis. Mutacijos, sukeliančios ligą ar organizmo savybių pablogėjimą, laikomos neigiamomis, o mutacijos, dėl kurių susidaro naujos naudingos savybės – teigiamos.
Tačiau, kalbant apie daugumą ligų, kuriomis šiandien serga žmonės, tai nėra paveldima liga, o tik polinkis. Pavyzdžiui, cukrų vaiko tėvas pasisavina lėtai. Tai nereiškia, kad vaikas gims sergantis diabetu, tačiau vaikas turės polinkį. Tai reiškia, kad jei vaikas piktnaudžiauja saldumynais ir miltiniais gaminiais, jam išsivystys cukrinis diabetas.
Šiandien vadinamasis predikatyvus vaistas. Šios medicinos praktikos metu nustatomi asmens polinkiai (remiantis atitinkamų genų identifikavimu), o tada jam pateikiamos rekomendacijos - kokios dietos laikytis, kaip teisingai kaitalioti darbo ir poilsio režimą, kad kad nesusirgtų.
Kaip skaityti informaciją, užkoduotą DNR?
Kaip galite perskaityti DNR esančią informaciją? Kaip jos kūnas tuo naudojasi? Pati DNR yra savotiška matrica, bet ne paprasta, o užkoduota. Norint nuskaityti informaciją iš DNR matricos, ji pirmiausia perkeliama į specialų nešiklį – RNR. RNR yra chemiškai ribonukleino rūgštis. Nuo DNR ji skiriasi tuo, kad pro branduolio membraną gali prasiskverbti į ląstelę, o DNR neturi šio gebėjimo (gali būti tik branduolyje). Užkoduota informacija naudojama pačioje ląstelėje. Taigi, RNR yra užkoduotos informacijos nešėja iš branduolio į ląstelę.Kaip sintetinama RNR, kaip RNR pagalba sintetinami baltymai?
 DNR gijos, iš kurių reikia „skaityti“ informaciją, išsivynioja, prie jų privažiuoja specialus fermentas – „statybininkas“ ir lygiagrečiai DNR grandinei susintetina papildomą RNR grandinę. RNR molekulė taip pat susideda iš 4 tipų nukleotidų – adenino (A), uracilo (U), guanino (G) ir citozino (C). Šiuo atveju šios poros yra viena kitą papildančios: adeninas - uracilas, guaninas - citozinas. Kaip matote, skirtingai nei DNR, RNR vietoj timino naudoja uracilą. Tai yra, fermentas „statybininkas“ veikia taip: jei jis mato A DNR grandinėje, tada jis prijungia Y prie RNR grandinės, jei G, tada prijungia C ir kt. Taigi iš kiekvieno aktyvaus geno transkripcijos metu susidaro šablonas – RNR kopija, kuri gali praeiti pro branduolio membraną.
DNR gijos, iš kurių reikia „skaityti“ informaciją, išsivynioja, prie jų privažiuoja specialus fermentas – „statybininkas“ ir lygiagrečiai DNR grandinei susintetina papildomą RNR grandinę. RNR molekulė taip pat susideda iš 4 tipų nukleotidų – adenino (A), uracilo (U), guanino (G) ir citozino (C). Šiuo atveju šios poros yra viena kitą papildančios: adeninas - uracilas, guaninas - citozinas. Kaip matote, skirtingai nei DNR, RNR vietoj timino naudoja uracilą. Tai yra, fermentas „statybininkas“ veikia taip: jei jis mato A DNR grandinėje, tada jis prijungia Y prie RNR grandinės, jei G, tada prijungia C ir kt. Taigi iš kiekvieno aktyvaus geno transkripcijos metu susidaro šablonas – RNR kopija, kuri gali praeiti pro branduolio membraną. Kaip vyksta specifinio geno koduoto baltymo sintezė?
Išėjus iš branduolio, RNR patenka į citoplazmą. Jau citoplazmoje RNR, kaip matrica, gali būti įterpta į specialias fermentų sistemas (ribosomas), kurios, vadovaudamosi RNR informacija, gali sintetinti atitinkamą baltymo aminorūgščių seką. Kaip žinote, baltymų molekulė susideda iš aminorūgščių. Kaip ribosomai pavyksta išsiaiškinti, kurią aminorūgštį reikia prijungti prie augančios baltymų grandinės? Tai daroma remiantis tripleto kodu. Tripleto kodas reiškia, kad trijų RNR grandinės nukleotidų seka ( trynukas, pavyzdžiui, HGH) koduoja vieną aminorūgštį (šiuo atveju gliciną). Kiekvieną aminorūgštį koduoja specifinis tripletas. Taigi, ribosoma „skaito“ tripletą ir nustato, kuri aminorūgštis turėtų būti prijungta toliau, nes ji nuskaito informaciją RNR. Susidarius aminorūgščių grandinei, ji įgauna tam tikrą erdvinę formą ir tampa baltymu, galinčiu atlikti jai priskirtas fermentines, statybines, hormonines ir kitas funkcijas.Bet kurio gyvo organizmo baltymas yra geno produktas. Būtent baltymai lemia visas įvairias genų savybes, savybes ir išorines apraiškas.
Dešinėje yra didžiausia žmogaus DNR spiralė, pastatyta iš žmonių Varnos (Bulgarija) paplūdimyje, kuri 2016 m. balandžio 23 d. pateko į Gineso rekordų knygą.
Deoksiribonukleorūgštis. Bendra informacija
DNR (dezoksiribonukleino rūgštis) yra tam tikras gyvenimo planas, sudėtingas kodas, kuriame yra duomenų apie paveldimą informaciją. Ši sudėtinga makromolekulė gali saugoti ir perduoti paveldimą genetinę informaciją iš kartos į kartą. DNR lemia tokias bet kurio gyvo organizmo savybes kaip paveldimumas ir kintamumas. Jame užkoduota informacija nustato visą bet kurio gyvo organizmo vystymosi programą. Genetiškai įgimti veiksniai nulemia visą žmogaus ir bet kurio kito organizmo gyvenimo eigą. Dirbtinis arba natūralus išorinės aplinkos poveikis gali tik nežymiai paveikti bendrą atskirų genetinių savybių sunkumą arba paveikti užprogramuotų procesų raidą.
Deoksiribonukleorūgštis(DNR) – makromolekulė (viena iš trijų pagrindinių, kitos dvi – RNR ir baltymai), kuri užtikrina saugojimą, perdavimą iš kartos į kartą ir gyvų organizmų vystymosi ir funkcionavimo genetinės programos įgyvendinimą. DNR yra informacijos apie įvairių tipų RNR ir baltymų struktūrą.
Eukariotinėse ląstelėse (gyvūnuose, augaluose ir grybuose) DNR randama ląstelės branduolyje kaip chromosomų dalis, taip pat kai kuriose ląstelių organelėse (mitochondrijose ir plastidėse). Prokariotinių organizmų (bakterijų ir archajų) ląstelėse iš vidaus prie ląstelės membranos yra prijungta apskrita arba linijinė DNR molekulė, vadinamasis nukleoidas. Jie ir žemesni eukariotai (pavyzdžiui, mielės) taip pat turi mažas, autonomines, daugiausia žiedines DNR molekules, vadinamas plazmidėmis.
Cheminiu požiūriu DNR yra ilga polimero molekulė, sudaryta iš pasikartojančių blokų – nukleotidų. Kiekvienas nukleotidas sudarytas iš azoto bazės, cukraus (dezoksiribozės) ir fosfato grupės. Ryšiai tarp nukleotidų grandinėje susidaro dėl dezoksiribozės ( SU) ir fosfatas ( F) grupės (fosfodiesteriniai ryšiai).

Ryžiai. 2. Nukletidas susideda iš azoto bazės, cukraus (dezoksiribozės) ir fosfatų grupės
Daugeliu atvejų (išskyrus kai kuriuos virusus, turinčius vienos grandinės DNR), DNR makromolekulę sudaro dvi grandinės, orientuotos viena į kitą azotinėmis bazėmis. Ši dvigrandė molekulė yra susukta spiraline linija.
DNR yra keturių tipų azoto bazės (adeninas, guaninas, timinas ir citozinas). Vienos grandinės azotinės bazės yra sujungtos su kitos grandinės azotinėmis bazėmis vandeniliniais ryšiais pagal komplementarumo principą: adeninas jungiasi tik su timinu ( AT), guaninas – tik su citozinu ( G-C). Būtent šios poros sudaro spiralinių DNR „laiptų“ „skersinius“ (žr. 2, 3 ir 4 pav.).

Ryžiai. 2. Azoto bazės
Nukleotidų seka leidžia „užkoduoti“ informaciją apie skirtingi tipai RNR, iš kurių svarbiausios yra informacinės, arba pasiuntinio (mRNR), ribosominės (rRNR) ir transportinės (tRNR). Visi šie RNR tipai yra sintetinami DNR šablone, kopijuojant DNR seką į transkripcijos proceso metu susintetintą RNR seką ir dalyvauja baltymų biosintezėje (vertimo procese). Be koduojančių sekų, ląstelės DNR yra sekų, kurios atlieka reguliavimo ir struktūrines funkcijas.

Ryžiai. 3. DNR replikacija
Pagrindinių cheminių DNR junginių derinių išsidėstymas ir kiekybiniai šių derinių ryšiai užtikrina paveldimos informacijos kodavimą.
Išsilavinimas nauja DNR (replikacija)
- Replikacijos procesas: DNR dvigubos spiralės išvyniojimas - komplementarių grandinių sintezė naudojant DNR polimerazę - dviejų DNR molekulių susidarymas iš vienos.
- Dviguba spiralė „atsiriša“ į dvi šakas, kai fermentai nutraukia ryšį tarp cheminių junginių bazinių porų.
- Kiekviena šaka yra naujos DNR elementas. Naujos bazių poros sujungiamos ta pačia seka kaip ir pirminėje šakoje.
Pasibaigus dubliavimui, susidaro dvi nepriklausomos spiralės, sukurtos iš pirminės DNR cheminių junginių ir turinčios tą patį genetinį kodą. Tokiu būdu DNR gali virškinti informaciją iš ląstelės į ląstelę.
Išsamesnė informacija:
NULEORŪGŠČIŲ STRUKTŪRA

Ryžiai. 4 . Azoto bazės: adeninas, guaninas, citozinas, timinas
Deoksiribonukleorūgštis(DNR) reiškia nukleino rūgštis. Nukleino rūgštys yra netaisyklingų biopolimerų klasė, kurios monomerai yra nukleotidai.
NUKLEOTIDAI susideda iš azoto bazė kartu su penkių angliavandenių angliavandeniais (pentoze) - dezoksiribozė(DNR atveju) arba ribozė(RNR atveju), kuri jungiasi su fosforo rūgšties liekana (H 2 PO 3 -).
Azoto bazės yra dviejų tipų: pirimidino bazės – uracilas (tik RNR), citozinas ir timinas, purino bazės – adeninas ir guaninas.

Ryžiai. 5. Nukleotidų struktūra (kairėje), nukleotido vieta DNR (apačioje) ir azotinių bazių tipai (dešinėje): pirimidinas ir purinas

Anglies atomai pentozės molekulėje sunumeruoti nuo 1 iki 5. Fosfatas jungiasi su trečiuoju ir penktuoju anglies atomais. Taip nukleotidai susijungia ir sudaro nukleorūgščių grandinę. Taigi galime išskirti 3 ir 5 DNR grandinės galus:

Ryžiai. 6. DNR grandinės 3 'ir 5' galų išskyrimas
Susidaro dvi DNR grandinės dviguba spiralė... Šios grandinės spirale yra nukreiptos priešingomis kryptimis. Skirtingose DNR grandinėse azoto bazės yra tarpusavyje sujungtos vandeniliniai ryšiai... Adeninas visada jungiasi su timinu, o citozinas su guaninu. Tai vadinama papildomumo taisyklė(cm. papildomumo principas).
Komplementarumo taisyklė:
| A-T G-C |
Pavyzdžiui, jei mums duota DNR grandinė su seka
3'- ATGTCCTAGCTGCTCG - 5',
tada antroji grandinė papildys ją ir nukreipta priešinga kryptimi - nuo 5' galo iki 3' galo:
5'-TACAGGATCGACGAGC-3'.

Ryžiai. 7. DNR molekulės grandinių kryptis ir azoto bazių sujungimas naudojant vandenilinius ryšius
DNR REPLIKACIJA
DNR replikacija yra DNR molekulės padvigubinimo procesas matricos sintezės būdu. Daugeliu atvejų natūrali DNR replikacijagruntasnes DNR sintezė yra trumpas fragmentas (atkurta iš naujo). Tokį ribonukleotidinį pradmenį sukuria fermentas primazė (prokariotuose – DNR primazė, eukariotuose – DNR polimerazė), o vėliau pakeičiamas dezoksiribonukleotidų polimeraze, kuri paprastai atlieka taisymo funkcijas (koreguoja cheminius pažeidimus ir DNR molekulės lūžius).
Replikacija vyksta pusiau konservatyviu mechanizmu. Tai reiškia, kad dviguba DNR spiralė išsivynioja ir pagal komplementarumo principą kiekvienoje jos grandinėje užbaigiama nauja grandinė. Todėl dukterinėje DNR molekulėje yra viena grandinė iš pagrindinės molekulės ir viena naujai susintetinta. Replikacija vyksta kryptimi nuo 3 "iki 5" pagrindinės grandinės galo.

Ryžiai. 8. DNR molekulės replikacija (dvigubėjimas).
DNR sintezė- tai nėra toks sudėtingas procesas, kaip gali pasirodyti iš pirmo žvilgsnio. Jei galvojate apie tai, pirmiausia turite išsiaiškinti, kas yra sintezė. Tai procesas, kai ką nors sujungiame. Naujos DNR molekulės formavimas vyksta keliais etapais:
1) DNR topoizomerazė, esanti prieš replikacijos šakutę, supjausto DNR, kad palengvintų jos išsivyniojimą ir išsivyniojimą.
2) DNR helikazė, eidama po topoizomerazės, įtakoja DNR spiralės „atsisukimo“ procesą.
3) DNR surišantys baltymai atlieka DNR grandinių surišimą, taip pat atlieka jų stabilizavimą, neleisdami joms prilipti vienas prie kito.
4) DNR polimerazė δ(delta) , suderintas su replikacinės šakutės judėjimo greičiu, atlieka sintezępirmaujantisgrandines dukterinė įmonė DNR šablone 5 "→ 3" kryptimi motiniškas DNR grandinė kryptimi nuo 3 "galo iki 5" galo (greitis iki 100 bazinių porų per sekundę). Šie įvykiai apie tai motiniškas DNR grandinės yra ribotos.

Ryžiai. 9. Scheminis DNR replikacijos proceso vaizdavimas: (1) atsiliekanti grandinė (atsilikusi grandinė), (2) pirmaujanti grandinė (pirmaujanti grandinė), (3) DNR polimerazė α (Polα), (4) DNR ligazė, (5) RNR -pradmenys, (6) primazė, (7) Okazaki fragmentas, (8) DNR polimerazė δ (Polδ), (9) helikazė, (10) viengrandžiai DNR surišantys baltymai, (11) topoizomerazė.
Toliau aprašoma atsiliekančios dukterinės DNR grandinės sintezė (žr. Schema replikacijos šakutė ir replikacijos fermento funkcija)
Norėdami gauti vizualesnį DNR replikacijos paaiškinimą, žr
5) Iš karto po kitos pagrindinės molekulės gijos išvyniojimo ir stabilizavimo,DNR polimerazė α(alfa)ir 5 "→ 3" kryptimi sintezuoja pradmenį (RNR pradmenį) - RNR seką ant DNR šablono nuo 10 iki 200 nukleotidų ilgio. Po to fermentaspašalinama iš DNR grandinės.
Vietoj DNR polimerazėα
pritvirtinamas prie 3 colių grunto galo DNR polimerazėε
.
6)
DNR polimerazėε
(epsilonas) tarsi ir toliau ilgina gruntą, bet kaip substratą įsitaisodezoksiribonukleotidai(150-200 nukleotidų kiekiu). Dėl to iš dviejų dalių susidaro tvirtas siūlas -RNR(t.y. gruntas) ir DNR.
DNR polimerazė εveikia tol, kol atitinka ankstesnį gruntąOkazaki fragmentas(sintetintas šiek tiek anksčiau). Tada šis fermentas pašalinamas iš grandinės.
7) DNR polimerazė β(beta) atsikeliaDNR polimerazė ε,juda ta pačia kryptimi (5 "→ 3") ir pašalina pradmenų ribonukleotidus, o į jų vietą įterpia dezoksiribonukleotidus. Fermentas veikia iki visiško grunto pašalinimo, t.y. iki dezoksiribonukleotido (dar anksčiau susintetintoDNR polimerazė ε). Fermentas nesugeba sujungti savo darbo rezultato ir prieš jį esančios DNR, todėl palieka grandinę.
Dėl to ant motininės gijos matricos „guli“ dukterinės DNR fragmentas. Tai vadinamaOkazaki fragmentas.
8) DNR ligazė susiuva du gretimus Okazaki fragmentai , t.y. Susintetintas 5 colių segmento galasDNR polimerazė ε,ir 3 colių grandinės galas, įmontuotasDNR polimerazėβ .
RNR STRUKTŪRA
Ribonukleino rūgštis(RNR) yra viena iš trijų pagrindinių makromolekulių (kitos dvi yra DNR ir baltymai), randamų visų gyvų organizmų ląstelėse.
Kaip ir DNR, RNR sudaro ilga grandinė, kurioje kiekviena grandis vadinama nukleotidas... Kiekvienas nukleotidas susideda iš azoto bazės, ribozės cukraus ir fosfatų grupės. Tačiau, skirtingai nei DNR, RNR paprastai turi ne dvi, o vieną grandinę. Pentozę RNR vaizduoja ribozė, o ne dezoksiribozė (antrame angliavandenių atome ribozė turi papildomą hidroksilo grupę). Galiausiai DNR nuo RNR skiriasi azoto bazių sudėtimi: vietoj timino ( T) uracilas ( U) kuris taip pat papildo adeniną.
Nukleotidų seka leidžia RNR koduoti genetinę informaciją. Visi ląsteliniai organizmai baltymų sintezei programuoti naudoja RNR (mRNR).
Ląstelinės RNR gaminamos procesu, vadinamu transkripcija , tai yra, RNR sintezė ant DNR matricos, atliekama specialiais fermentais - RNR polimerazės.
Tada pasiuntiniai RNR (mRNR) dalyvauja procese, vadinamame transliacija, tie. baltymų sintezė mRNR matricoje dalyvaujant ribosomoms. Kitos RNR po transkripcijos patiria cheminių modifikacijų, o susidariusios antrinėms ir tretinėms struktūroms atlieka funkcijas, priklausomai nuo RNR tipo.

Ryžiai. 10. Skirtumas tarp DNR ir RNR azoto bazėje: vietoj timino (T), RNR yra uracilas (U), kuris taip pat yra adenino komplementas.
PRANEŠIMAS
Tai RNR sintezės DNR šablone procesas. DNR išsivynioja vienoje iš vietų. Vienoje iš gijų yra informacijos, kurią reikia nukopijuoti į RNR molekulę – ši grandinė vadinama koduojančia grandine. Antroji DNR grandinė, papildanti koduojančiąją, vadinama šablonu. Transkripcijos procese šabloninėje grandinėje 3 '- 5' kryptimi (išilgai DNR grandinės), susintetinama papildoma RNR grandinė. Taigi sukuriama koduojančios grandinės RNR kopija.
![]()
Ryžiai. 11. Scheminis transkripcijos vaizdavimas
Pavyzdžiui, jei mums pateikiama koduojančios grandinės seka
3'- ATGTCCTAGCTGCTCG - 5',
tada pagal komplementarumo taisyklę matricos grandinė neš seką
5'- TACAGGATCGACGAGC- 3',
o iš jos susintetinta RNR yra seka
Transliacija
Apsvarstykite mechanizmą baltymų sintezė ant RNR matricos, taip pat genetinis kodas ir jo savybės. Be to, siekiant aiškumo, naudojant žemiau esančią nuorodą, rekomenduojame pažiūrėti trumpą vaizdo įrašą apie transkripcijos ir vertimo procesus, vykstančius gyvoje ląstelėje:

Ryžiai. 12. Baltymų sintezės procesas: DNR koduoja RNR, RNR koduoja baltymus
GENETINIS KODAS
Genetinis kodas- baltymų aminorūgščių sekos kodavimo būdas naudojant nukleotidų seką. Kiekvieną aminorūgštį koduoja trijų nukleotidų seka – kodonas arba tripletas.
Genetinis kodas, būdingas daugumai pro- ir eukariotų. Lentelėje išvardyti visi 64 kodonai ir nurodytos atitinkamos aminorūgštys. Bazinė tvarka yra nuo 5 "iki 3" mRNR galo.
1 lentelė. Standartinis genetinis kodas
|
1-oji ne |
2-oji bazė |
3 ne |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe / F) |
U C U |
(Ser/S) |
U A U |
(Tyr / Y) |
U G U |
(Cys / C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Lė / L) |
U C A |
U A A |
Sustabdyti kodoną** |
U G A |
Sustabdyti kodoną** |
A |
||
|
U U G |
U C G |
U A G |
Sustabdyti kodoną** |
U G G |
(Trp / W) |
G |
|||
|
C |
C U U |
C C U |
(už / P) |
C A U |
(Jo / H) |
C G U |
(Arg / R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln / Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile / I) |
A C U |
(Thr / T) |
A A U |
(Asn / N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys / K) |
A G A |
A |
||||
|
A U G |
(Met / M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val / V) |
G C U |
(Ala / A) |
G A U |
(Asp / D) |
G G U |
(Gly / G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu / E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Tarp trynukų yra 4 specialios sekos, kurios veikia kaip „skyrybos ženklai“:
- *Trejetas RUG, taip pat koduojantis metioniną, vadinamas pradžios kodonas... Nuo šio kodono prasideda baltymo molekulės sintezė. Taigi baltymų sintezės metu pirmoji aminorūgštis sekoje visada bus metioninas.
- ** Trynukai UAA, UAG ir UGA yra vadinami stop kodonai ir neužkoduoja nei vienos aminorūgšties. Šiose sekose baltymų sintezė sustoja.
Genetinio kodo savybės
1. Trijulė... Kiekvieną aminorūgštį koduoja trijų nukleotidų seka – tripletas arba kodonas.
2. Tęstinumas... Tarp tripletų papildomų nukleotidų nėra, informacija skaitoma nuolat.
3. Nepersidengimas... Vienas nukleotidas negali vienu metu patekti į du tripletus.
4. Vienareikšmiškumas... Vienas kodonas gali koduoti tik vieną aminorūgštį.

5. Degeneracija... Viena aminorūgštis gali būti koduota keliais skirtingais kodonais.

6. Universalumas... Genetinis kodas yra vienodas visiems gyviems organizmams.
Pavyzdys. Mums suteikiama kodavimo grandinės seka:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Matricos grandinės seka bus tokia:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Dabar mes „sintetiname“ informacinę RNR iš šios grandinės:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Baltymų sintezė vyksta 5 '→ 3' kryptimi, todėl, norėdami "skaityti" genetinį kodą, turime apversti seką:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
Dabar suraskime AUG pradžios kodoną:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
Padalinkime seką į tripletus:
skamba taip: informacija iš DNR perkeliama į RNR (transkripcija), iš RNR – į baltymą (vertimas). DNR gali būti dubliuojama ir replikacijos būdu, galimas ir atvirkštinės transkripcijos procesas, kai DNR sintetinama iš RNR šablono, tačiau šis procesas daugiausia būdingas virusams.

Ryžiai. 13. Centrinė dogma molekulinė biologija
GENOMAS: GENAI ir CHROMOSOMOS
(bendrosios sąvokos)
Genomas – visų organizmo genų visuma; visas jo chromosomų rinkinys.
Terminą „genomas“ 1920 metais pasiūlė G. Winkleris, kad apibūdintų genų rinkinį, esantį haploidiniame vienos biologinės rūšies organizmų chromosomų rinkinyje. Pirminė šio termino reikšmė nurodė, kad genomo samprata, priešingai nei genotipas, yra visos rūšies, o ne atskiro individo, genetinė savybė. Tobulėjant molekulinei genetikai šio termino reikšmė pasikeitė. Yra žinoma, kad DNR, kuri yra daugumos organizmų genetinės informacijos nešėja ir todėl yra genomo pagrindas, apima ne tik genus šiuolaikine šio žodžio prasme. Dauguma Eukariotinių ląstelių DNR atstovauja nekoduojančios ("perteklinės") nukleotidų sekos, kuriose nėra informacijos apie baltymus ir nukleino rūgštis. Taigi pagrindinė bet kurio organizmo genomo dalis yra visa jo haploidinio chromosomų rinkinio DNR.
Genai yra DNR molekulių sekcijos, koduojančios polipeptidus ir RNR molekules
Per pastarąjį šimtmetį mūsų supratimas apie genus labai pasikeitė. Anksčiau genomu buvo vadinama chromosomos dalis, kuri koduoja arba nustato vieną požymį arba fenotipinis(matoma) savybė, pvz., akių spalva.
1940 m. George'as Beadle'as ir Edwardas Tatemas pasiūlė molekulinį geno apibrėžimą. Mokslininkai gydė grybelių sporas Neurospora crassa Rentgeno spinduliai ir kitos medžiagos, sukeliančios DNR sekos pokyčius ( mutacijos) ir aptiktos mutantinės grybelio padermės, praradusios kai kuriuos specifinius fermentus, o tai kai kuriais atvejais sutrikdė visą medžiagų apykaitos kelią. Beadle ir Tatem padarė išvadą, kad genas yra genetinės medžiagos dalis, kuri apibrėžia arba koduoja vieną fermentą. Taip atsirado hipotezė „Vienas genas – vienas fermentas“... Vėliau ši sąvoka buvo išplėsta siekiant apibrėžti „Vienas genas – vienas polipeptidas“, nes daugelis genų koduoja baltymus, kurie nėra fermentai, o polipeptidas gali būti sudėtingo baltymų komplekso subvienetas.
Fig. 14 yra diagrama, kaip DNR nukleotidų tripletai nustato polipeptidą, baltymo aminorūgščių seką, kurią tarpininkauja mRNR. Viena iš DNR grandžių atlieka mRNR sintezės šablono vaidmenį, kurio nukleotidų tripletai (kodonai) yra komplementarūs DNR tripletams. Kai kuriose bakterijose ir daugelyje eukariotų koduojančias sekas pertraukia nekoduojantys regionai (vadinamieji). intronai).
Šiuolaikinis biocheminis genų apibrėžimas dar konkrečiau. Genai yra visos DNR sekcijos, koduojančios pirminę galutinių produktų seką, įskaitant polipeptidus arba RNR, turinčius struktūrinę arba katalizinę funkciją.
Kartu su genais DNR yra ir kitų sekų, kurios atlieka išskirtinai reguliavimo funkciją. Reguliavimo sekos gali žymėti genų pradžią arba pabaigą, paveikti transkripciją arba nurodyti replikacijos ar rekombinacijos pradžios vietą. Kai kurie genai gali būti išreikšti skirtingais būdais, o ta pati DNR dalis yra skirtingų produktų formavimo šablonas.
Galime apytiksliai apskaičiuoti minimalus geno dydis koduoja vidutinį baltymą. Kiekviena aminorūgštis polipeptidinėje grandinėje yra užkoduota kaip trijų nukleotidų seka; šių tripletų (kodonų) sekos atitinka duoto geno koduojamą aminorūgščių grandinę polipeptide. 350 aminorūgščių liekanų polipeptidinė grandinė (vidutinė grandinė) atitinka 1050 bp seką. ( bazinės poros). Tačiau daugelis eukariotų genų ir kai kurių prokariotų genų yra pertraukiami DNR segmentų, kurie nėra informacijos nešėjai apie baltymą, todėl pasirodo esąs daug ilgesnis, nei rodo paprastas skaičiavimas.
Kiek genų yra vienoje chromosomoje?
 Ryžiai. 15. Chromosomų vaizdas prokaritinėse (kairėje) ir eukariotinėse ląstelėse. Histonai yra plati branduolinių baltymų klasė, atliekanti dvi pagrindines funkcijas: jie dalyvauja DNR grandinių pakavime branduolyje ir epigenetiniame branduolinių procesų, tokių kaip transkripcija, replikacija ir taisymas, reguliavime.
Ryžiai. 15. Chromosomų vaizdas prokaritinėse (kairėje) ir eukariotinėse ląstelėse. Histonai yra plati branduolinių baltymų klasė, atliekanti dvi pagrindines funkcijas: jie dalyvauja DNR grandinių pakavime branduolyje ir epigenetiniame branduolinių procesų, tokių kaip transkripcija, replikacija ir taisymas, reguliavime.
Prokariotų DNR yra paprastesnė: jų ląstelės neturi branduolio, todėl DNR yra tiesiai citoplazmoje nukleoido pavidalu.
 Kaip žinote, bakterijų ląstelės turi chromosomą DNR grandinės pavidalu, supakuotą į kompaktišką struktūrą – nukleoidą. Prokarioto chromosoma Escherichia coli, kurio genomas buvo visiškai iššifruotas, yra apskrita DNR molekulė (iš tikrųjų taip nėra teisingas ratas, o kilpa be pradžios ir pabaigos), susidedanti iš 4 639 675 bp. Šioje sekoje yra maždaug 4300 genų baltymams ir 157 genai stabilioms RNR molekulėms. V žmogaus genomas maždaug 3,1 milijardo bazinių porų, atitinkančių beveik 29 000 genų, esančių 24 skirtingose chromosomose.
Kaip žinote, bakterijų ląstelės turi chromosomą DNR grandinės pavidalu, supakuotą į kompaktišką struktūrą – nukleoidą. Prokarioto chromosoma Escherichia coli, kurio genomas buvo visiškai iššifruotas, yra apskrita DNR molekulė (iš tikrųjų taip nėra teisingas ratas, o kilpa be pradžios ir pabaigos), susidedanti iš 4 639 675 bp. Šioje sekoje yra maždaug 4300 genų baltymams ir 157 genai stabilioms RNR molekulėms. V žmogaus genomas maždaug 3,1 milijardo bazinių porų, atitinkančių beveik 29 000 genų, esančių 24 skirtingose chromosomose.
Prokariotai (bakterijos).
 Bakterija E. coli turi vieną dvigrandę žiedinę DNR molekulę. Jį sudaro 4 639 675 bp. ir pasiekia apie 1,7 mm ilgį, o tai viršija pačios ląstelės ilgį E. coli maždaug 850 kartų. Be didelės žiedinės chromosomos nukleoide, daugelis bakterijų turi vieną ar daugiau mažų žiedinių DNR molekulių, kurios laisvai išsidėsčiusios citozolyje. Tokie ekstrachromosominiai elementai vadinami plazmidės(16 pav.).
Bakterija E. coli turi vieną dvigrandę žiedinę DNR molekulę. Jį sudaro 4 639 675 bp. ir pasiekia apie 1,7 mm ilgį, o tai viršija pačios ląstelės ilgį E. coli maždaug 850 kartų. Be didelės žiedinės chromosomos nukleoide, daugelis bakterijų turi vieną ar daugiau mažų žiedinių DNR molekulių, kurios laisvai išsidėsčiusios citozolyje. Tokie ekstrachromosominiai elementai vadinami plazmidės(16 pav.).
Daugumą plazmidžių sudaro tik keli tūkstančiai bazinių porų, kai kurios turi daugiau nei 10 000 bp. Jie neša genetinę informaciją ir dauginasi susidarant dukterinėms plazmidėms, kurios patenka į dukterines ląsteles dalijantis pirminei ląstelei. Plazmidžių yra ne tik bakterijose, bet ir mielėse bei kituose grybuose. Daugeliu atvejų plazmidės nesuteikia jokio pranašumo ląstelėms šeimininkėms, o vienintelė jų užduotis yra daugintis savarankiškai. Tačiau kai kurios plazmidės turi šeimininkui naudingus genus. Pavyzdžiui, plazmidėse esantys genai gali suteikti bakterijų ląstelėms atsparumą antibakterinėms medžiagoms. Plazmidės, turinčios β-laktamazės geną, suteikia atsparumą β-laktaminiams antibiotikams, tokiems kaip penicilinas ir amoksicilinas. Plazmidės gali būti perkeltos iš antibiotikams atsparių ląstelių į kitas tos pačios arba skirtingų rūšių bakterijų ląsteles, todėl šios ląstelės taip pat tampa atsparios. Intensyvus antibiotikų vartojimas yra galingas selektyvus veiksnys, skatinantis plazmidžių, koduojančių atsparumą antibiotikams (taip pat transpozonų, koduojančių panašius genus), plitimą tarp patogeninių bakterijų ir sukeliančių bakterijų padermių, atsparių keliems antibiotikams, atsiradimą. Gydytojai pradeda suprasti plataus antibiotikų vartojimo pavojus ir skiria juos tik tada, kai reikia skubiai. Dėl panašių priežasčių plačiai paplitęs antibiotikų naudojimas ūkinių gyvūnų gydymui yra ribotas.
Taip pat žiūrėkite: Ravinas N.V., Šestakovas S.V. Prokariotų genomas // Vavilov Journal of Genetics and Selection, 2013. V. 17. Nr. 4/2. S. 972-984.
Eukariotai.
2 lentelė. Kai kurių organizmų DNR, genai ir chromosomos
|
Bendra DNR, p.n. |
Chromosomų skaičius* |
Apytikslis genų skaičius |
|
|
Escherichia coli(bakterija) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(mielės) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegantiškas(nematodas) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(augalas) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(vaisinė musė) |
120 367 260 |
20 000 |
|
|
Oryza sativa(ryžiai) |
480 000 000 |
57 000 |
|
|
Musculus(pelė) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens(Žmogus) |
3 070 128 600 |
29 000 |
Pastaba. Informacija nuolat atnaujinama; Norėdami gauti daugiau naujausios informacijos, žr. svetaines, skirtas atskiriems genominiams projektams
* Visiems eukariotams, išskyrus mieles, pateikiamas diploidinis chromosomų rinkinys. Diploidas rinkinys chromosomos (iš graikų kalbos diploos - double ir eidos - rūšys) - dvigubas chromosomų rinkinys(2n), kurių kiekvienas turi homologinį.
** Haploidinis rinkinys. Laukinių mielių padermės paprastai turi aštuonis (oktaploidinius) ar daugiau tokių chromosomų rinkinių.
*** Moterims, turinčioms dvi X chromosomas. Vyrai turi X chromosomą, bet neturi Y, tai yra, yra tik 11 chromosomų.
Mielių ląstelė, viena iš mažiausių eukariotų, turi 2,6 karto daugiau DNR nei ląstelė E. coli(2 lentelė). Vaisinės muselės ląstelės Drosophila, klasikinis genetinių tyrimų objektas, turi 35 kartus daugiau DNR, o žmogaus ląstelėse – apie 700 kartų daugiau DNR nei ląstelėse. E. coli. Daugelyje augalų ir varliagyvių yra dar daugiau DNR. Eukariotinių ląstelių genetinė medžiaga yra organizuota chromosomų pavidalu. Diploidinis chromosomų rinkinys (2 n) priklauso nuo organizmo tipo (2 lentelė).
Pavyzdžiui, žmogaus somatinėje ląstelėje yra 46 chromosomos ( ryžių. 17). Kiekviena eukariotinės ląstelės chromosoma, kaip parodyta Fig. 17, a, yra viena labai didelė dvigrandė DNR molekulė. Dvidešimt keturios žmogaus chromosomos (22 suporuotos chromosomos ir dvi lytinės chromosomos X ir Y) skiriasi savo ilgiu daugiau nei 25 kartus. Kiekvienoje eukariotų chromosomoje yra tam tikras genų rinkinys.

Ryžiai. 17. Eukariotų chromosomos.a- susietų ir kondensuotų seserinių chromatidžių pora iš žmogaus chromosomos. Šioje formoje eukariotinės chromosomos išlieka po replikacijos ir metafazėje mitozės metu. b- visas chromosomų rinkinys iš vieno iš knygos autorių leukocitų. Kiekvienoje normalioje žmogaus somatinėje ląstelėje yra 46 chromosomos.

DNR, kaip paveldimos medžiagos saugojimo ir perdavimo matricos, dydis ir funkcija paaiškina specialių struktūrinių elementų buvimą šios molekulės organizacijoje. Aukštesniuose organizmuose DNR pasiskirsto tarp chromosomų.
Organizmo DNR (chromosomų) rinkinys vadinamas genomu. Chromosomos randamos ląstelės branduolyje ir sudaro struktūrą, vadinamą chromatinu. Chromatinas yra DNR ir bazinių baltymų (histonų) kompleksas santykiu 1:1. DNR ilgis paprastai matuojamas komplementarių nukleotidų porų (bp) skaičiumi. Pavyzdžiui, 3-oji chromosoma yra žmogausamžiuje yra DNR molekulė, kurios dydis yra 160 milijonų bp. Išskirta tiesinė DNR, kurios dydis yra 3 * 10 6 bp. ilgis yra apie 1 mm, todėl tiesinė 3-osios žmogaus chromosomos molekulė būtų 5 mm ilgio, o visų 23 haploido chromosomų (~ 3 * 10 9 bp, MR = 1,8 * 10 12) DNR ląstelė – kiaušialąstė arba spermatozoidas – tiesine forma būtų 1 m. Išskyrus lytines ląsteles, visose žmogaus kūno ląstelėse (jų yra apie 1013) yra dvigubas chromosomų rinkinys. Ląstelių dalijimosi metu visos 46 DNR molekulės yra replikuojamos ir pertvarkomos į 46 chromosomas.
Jei sujungsite žmogaus genomo DNR molekules (22 chromosomos ir chromosomos X ir Y arba X ir X), gausite maždaug vieno metro ilgio seką. Pastaba: Visi žinduoliai ir kiti organizmai, turintys heterogametinę vyriškąją lytį, patelės turi dvi X chromosomas (XX), o patinai turi vieną X chromosomą ir vieną Y chromosomą (XY).
Todėl daugumos žmogaus ląstelių bendras tokių ląstelių DNR ilgis yra apie 2 m. Suaugęs žmogus turi maždaug 10 14 ląstelių, todėl bendras visų DNR molekulių ilgis yra 2 ・ 10 11 km. Palyginimui, Žemės perimetras yra 4 ・ 10 4 km, o atstumas nuo Žemės iki Saulės yra 1,5 ・ 10 8 km. Štai kaip stebėtinai kompaktiškai supakuota DNR mūsų ląstelėse!
Eukariotinėse ląstelėse yra ir kitų DNR turinčių organelių – mitochondrijų ir chloroplastų. Buvo iškelta daug hipotezių dėl mitochondrijų ir chloroplastų DNR kilmės. Šiandien visuotinai priimtas požiūris yra tas, kad tai yra senovės bakterijų chromosomų užuomazgos, kurios pateko į šeimininkų ląstelių citoplazmą ir tapo šių organelių pirmtakais. Mitochondrijų DNR koduoja mitochondrijų tRNR ir rRNR, taip pat keletą mitochondrijų baltymų. Daugiau nei 95% mitochondrijų baltymų koduoja branduolio DNR.
GENŲ STRUKTŪRA
Apsvarstykite prokariotų ir eukariotų geno struktūrą, jų panašumus ir skirtumus. Nepaisant to, kad genas yra DNR dalis, koduojanti tik vieną baltymą arba RNR, be tiesiogiai koduojančios dalies, jis apima ir reguliuojančius bei kitus struktūrinius elementus, kurių prokariotuose ir eukariotuose yra skirtinga struktūra.
Kodavimo seka- pagrindinis struktūrinis ir funkcinis geno vienetas, jame yra nukleotidų tripletai, koduojantysaminorūgščių seka. Jis prasideda pradžios kodonu ir baigiasi stop kodonu.
Prieš ir po kodavimo sekos yra neišverstos 5' ir 3' sekos... Jie atlieka reguliavimo ir pagalbines funkcijas, pavyzdžiui, užtikrina ribosomos nusileidimą ant m-RNR.
Neverstos ir koduojančios sekos sudaro transkripcijos vienetą – transkribuotą DNR sekciją, tai yra DNR sekciją, iš kurios sintetinama m-RNR.
Terminatorius- netranskribuota DNR sritis geno gale, kurioje sustoja RNR sintezė.
Geno pradžioje yra reguliavimo sritisįskaitant propaguotojas ir operatorius.
Rėmėjas- seka, prie kurios polimerazė prisijungia transkripcijos inicijavimo metu. operatorius yra sritis, prie kurios gali prisijungti ypatingi baltymai - represorių, kuris gali sumažinti RNR sintezės iš šio geno aktyvumą – kitaip tariant, sumažinti išraiška.
Genų struktūra prokariotuose
Bendra prokariotų ir eukariotų genų struktūra nesiskiria – jie abu turi reguliavimo sritį su promotoriumi ir operatoriumi, transkripcijos vienetą su koduojančiomis ir neverčiamomis sekomis bei terminatorių. Tačiau prokariotų ir eukariotų genų organizacija skiriasi.

Ryžiai. 18. Prokariotų (bakterijų) geno sandaros schema -vaizdas padidintas
Operono pradžioje ir pabaigoje yra bendri kelių struktūrinių genų reguliavimo regionai. Viena mRNR molekulė nuskaitoma iš transkribuotos operono srities, kurioje yra kelios koduojančios sekos, kurių kiekviena turi savo pradžios ir pabaigos kodonus. Iš kiekvienos iš šių svetainių suvienas baltymas nutrūksta. Šiuo būdu, iš vienos i-RNR molekulės sintetinamos kelios baltymo molekulės.
Prokariotams būdinga sujungti kelis genus į vieną funkcinį vienetą - operonas... Operono darbą gali reguliuoti kiti genai, kurie gali būti pastebimai nutolę nuo paties operono - reguliatoriai... Iš šio geno išverstas baltymas vadinamas represorius... Jis jungiasi su operono operatoriumi, reguliuodamas visų jame esančių genų ekspresiją vienu metu.
Šis reiškinys būdingas ir prokariotams transkripcijos ir vertimo susiejimas.
![]()
Ryžiai. 19 Prokariotų transkripcijos ir vertimo konjugacijos reiškinys - vaizdas padidintas
Tokia konjugacija eukariotuose nevyksta dėl branduolio apvalkalo, kuris atskiria citoplazmą, kurioje vyksta transliacija, nuo genetinės medžiagos, ant kurios vyksta transkripcija. Prokariotuose RNR sintezės metu DNR šablone ribosoma gali iš karto prisijungti prie susintetintos RNR molekulės. Taigi, vertimas pradedamas dar nebaigus transkripcijos. Be to, kelios ribosomos vienu metu gali prisijungti prie vienos RNR molekulės, vienu metu sintetindamos kelias vieno baltymo molekules.
Genų struktūra eukariotuose
Eukariotų genai ir chromosomos yra labai sudėtingai organizuoti
Daugelis bakterijų rūšių turi tik vieną chromosomą ir beveik visais atvejais kiekvienoje chromosomoje yra po vieną kiekvieno geno kopiją. Tik keli genai, tokie kaip rRNR genai, yra keliose kopijose. Genai ir reguliavimo sekos sudaro beveik visą prokariotų genomą. Be to, beveik kiekvienas genas griežtai atitinka aminorūgščių seką (arba RNR seką), kurią jis koduoja (14 pav.).
Eukariotų genų struktūrinė ir funkcinė organizacija yra daug sudėtingesnė. Eukariotų chromosomų tyrimas, o vėliau ir pilnų eukariotų genomų sekų sekos nustatymas atnešė daug netikėtumų. Daugelis, jei ne dauguma, eukariotų genų turi įdomią savybę: jų nukleotidų sekose yra viena ar daugiau DNR sričių, kuriose nėra užkoduota polipeptidinio produkto aminorūgščių seka. Tokie neverčiami intarpai nutraukia tiesioginį geno nukleotidų sekos ir koduojamo polipeptido aminorūgščių sekos atitikimą. Šie neišversti genų segmentai vadinami intronai, arba įdėta sekos o kodavimo segmentai yra egzonai... Prokariotuose tik keli genai turi intronų.
Taigi eukariotuose praktiškai nėra genų derinio į operonus, o koduojanti eukarioto geno seka dažniausiai skirstoma į išverstas sritis. - egzonai, ir neišverstos skiltys – intronai.
Daugeliu atvejų intronų funkcija nebuvo nustatyta. Apskritai tik apie 1,5% žmogaus DNR yra „koduojama“, tai yra, jose yra informacija apie baltymus arba RNR. Tačiau, atsižvelgiant į didelius intronus, paaiškėja, kad 30% žmogaus DNR sudaro genai. Kadangi genai sudaro palyginti nedidelę žmogaus genomo dalį, didelė DNR dalis lieka neapskaityta.

Ryžiai. 16. Eukariotų geno sandaros schema - vaizdas padidintas
Iš kiekvieno geno pirmiausia susintetinama nesubrendusi arba pre-RNR, kurioje yra ir intronų, ir egzonų.
Po to vyksta susijungimo procesas, kurio metu išpjaunamos intronų sritys ir susidaro subrendusi iRNR, iš kurios galima susintetinti baltymą.

Ryžiai. 20. Alternatyvaus sujungimo procesas - vaizdas padidintas
Toks genų organizavimas leidžia, pavyzdžiui, suvokti, kada iš vieno geno gali būti susintetintos skirtingos baltymo formos dėl to, kad susijungimo procese egzonai gali būti susiuvami į skirtingas sekas.

Ryžiai. 21. Prokariotų ir eukariotų genų sandaros skirtumai - vaizdas padidintas
MUTACIJOS IR MUTAGENĖ
Mutacija vadinamas nuolatiniu genotipo pasikeitimu, tai yra nukleotidų sekos pasikeitimu.
Procesas, vedantis į mutacijų atsiradimą, vadinamas mutagenezė ir organizmas, visi kurių ląstelės turi tą pačią mutaciją - mutantas.
Mutacijų teorija pirmą kartą suformulavo Hugo de Vries 1903 m. Šiuolaikinėje jo versijoje yra šios nuostatos:
1. Mutacijos atsiranda staiga, staigiai.
2. Mutacijos perduodamos iš kartos į kartą.
3. Mutacijos gali būti naudingos, žalingos arba neutralios, dominuojančios arba recesyvinės.
4. Tikimybė aptikti mutacijas priklauso nuo tiriamų individų skaičiaus.
5. Panašios mutacijos gali kartotis.
6. Mutacijos nėra tikslinės.
Mutacijos gali atsirasti dėl įvairių veiksnių. Atskirkite mutacijas, atsiradusias veikiant mutageninis poveikio: fizinė (pavyzdžiui, ultravioletinė arba spinduliuotė), cheminė (pavyzdžiui, kolchicinas arba reaktyviosios deguonies rūšys) ir biologinė (pavyzdžiui, virusai). Taip pat mutacijas gali sukelti replikacijos klaidos.
Priklausomai nuo išvaizdos sąlygų, mutacijos skirstomos į spontaniškas- tai yra mutacijos, atsiradusios normaliomis sąlygomis, ir sukeltas- tai yra mutacijos, atsiradusios ypatingomis sąlygomis.
Mutacijos gali atsirasti ne tik branduolinėje DNR, bet ir, pavyzdžiui, mitochondrijų ar plastidžių DNR. Atitinkamai galime atskirti branduolinis ir citoplazminis mutacijos.
Dėl mutacijų dažnai gali atsirasti naujų alelių. Jei mutantinis alelis slopina normalaus alelio veikimą, vadinama mutacija dominuojantis... Jei normalus alelis slopina mutantą, tokia mutacija vadinama recesyvinis... Dauguma mutacijų, dėl kurių atsiranda naujų alelių, yra recesyvinės.
Pagal poveikį išskiriamos mutacijos prisitaikantis dėl to organizmas geriau prisitaiko prie aplinkos, neutralus kurie neturi įtakos išgyvenimui, kenksmingas kurie mažina organizmų prisitaikymą prie aplinkos sąlygų ir mirtinas sukelianti organizmo mirtį ankstyvosiose vystymosi stadijose.
Pagal pasekmes išskiriamos mutacijos, vedančios į baltymų funkcijos praradimas, mutacijos, sukeliančios atsiradimas baltymas atlieka naują funkciją, taip pat mutacijos, kurios pakeisti geno dozę, ir atitinkamai iš jo susintetinto baltymo dozė.
Mutacija gali atsirasti bet kurioje kūno ląstelėje. Jei lytinėje ląstelėje įvyksta mutacija, ji vadinama gemalinė(germinalinis arba generatyvinis). Tokios mutacijos neatsiranda tame organizme, kuriame jos atsirado, o veda prie mutantų atsiradimo palikuoniuose ir yra paveldimos, todėl yra svarbios genetikai ir evoliucijai. Jei mutacija įvyksta bet kurioje kitoje ląstelėje, ji vadinama somatinės... Tokia mutacija vienu ar kitu laipsniu gali pasireikšti organizme, kuriame ji atsirado, pavyzdžiui, sukelti vėžinių navikų susidarymą. Tačiau ši mutacija nėra paveldima ir nepaveikia palikuonių.
Mutacijos gali paveikti įvairaus dydžio genomo sritis. Paskirstyti genas, chromosominės ir genominis mutacijos.
Genų mutacijos
Mutacijos, kurios įvyksta mažesniu nei vieno geno mastu, vadinamos genetinė, arba taškas (taškas)... Tokios mutacijos lemia vieno ar daugiau sekos nukleotidų pasikeitimą. Tarp genų mutacijų yrapakaitalai dėl kurio vienas nukleotidas pakeičiamas kitu,ištrynimai dėl kurio prarandamas vienas iš nukleotidų,intarpai dėl to į seką pridedamas papildomas nukleotidas.

Ryžiai. 23. Genų (taškų) mutacijos
Pagal baltymų veikimo mechanizmą genų mutacijos skirstomos į:sinonimas, kurios (dėl genetinio kodo degeneracijos) nekeičia baltyminio produkto aminorūgščių sudėties,missense mutacijos, dėl kurių viena aminorūgštis pakeičiama kita ir gali turėti įtakos sintezuojamo baltymo struktūrai, nors dažnai pasirodo, kad jos yra nereikšmingos,nesąmonės mutacijos dėl ko koduojantis kodonas pakeičiamas stop kodonu,mutacijos, sukeliančios sujungimo sutrikimas:

Ryžiai. 24. Mutacijų schemos
Be to, pagal baltymo veikimo mechanizmą išskiriamos mutacijos, dėl kurių atsiranda rėmo poslinkis parodymai pavyzdžiui, įterpimai ir ištrynimai. Tokios mutacijos, kaip ir beprasmiškos mutacijos, nors ir įvyksta viename geno taške, dažnai paveikia visą baltymo struktūrą, o tai gali lemti visišką jo struktūros pasikeitimą. kai chromosomos dalis pasukama 180 laipsnių kampu, Ryžiai. 28. Translokacija
kai chromosomos dalis pasukama 180 laipsnių kampu, Ryžiai. 28. Translokacija
Ryžiai. 29. Chromosoma prieš ir po dubliavimosi
Genominės mutacijos
Pagaliau, genominės mutacijos paveikti visą genomą kaip visumą, tai yra, keičiasi chromosomų skaičius. Paskirstykite poliploidiją - ląstelių ploidijos padidėjimą ir aneuploidiją, tai yra chromosomų skaičiaus pasikeitimą, pavyzdžiui, trisomija (papildomo homologo buvimas vienoje iš chromosomų) ir monosomija (homologo nebuvimas chromosoma).
DNR vaizdo įrašai
DNR REPLIKACIJA, RNR KODAVIMAS, BALTYMŲ SINTEZĖ
(Jei vaizdo įrašas nerodomas, jis pasiekiamas per
Pagal savo cheminę struktūrą DNR ( Deoksiribonukleorūgštis) yra biopolimeras, kurio monomerai yra nukleotidai... Tai yra, DNR yra polinukleotidas... Be to, DNR molekulė paprastai susideda iš dviejų grandinių, susuktų viena kitos atžvilgiu išilgai spiralinės linijos (dažnai vadinamos „spirališkai susukta“) ir sujungtų vandeniliniais ryšiais.
Grandines galima susukti tiek į kairę, tiek į dešinę (dažniausiai) pusę.
Kai kurie virusai turi vieną DNR grandinę.
Kiekvienas DNR nukleotidas susideda iš 1) azoto bazės, 2) dezoksiribozės, 3) fosforo rūgšties liekanos.
Dviguba dešinės rankos DNR spiralė
DNR apima: adeninas, guaninas, timinas ir citozinas... Adeninas ir guaninas priklauso purinamas, o timinas ir citozinas – į pirimidinas... Kartais DNR yra uracilo, kuris dažniausiai būdingas RNR, kur jis pakeičia timiną.
Vienos DNR molekulės grandinės azotinės bazės yra sujungtos su kitos azotinėmis bazėmis griežtai pagal komplementarumo principą: adeninas tik su timinu (sudaro du vandenilinius ryšius tarpusavyje), o guaninas tik su citozinu (trys ryšiai) .
Azoto bazė pačiame nukleotide yra prijungta prie pirmojo ciklinės formos anglies atomo dezoksiribozė kuri yra pentozė (angliavandenis, turintis penkis anglies atomus). Ryšys yra kovalentinis, glikozidinis (C-N). Skirtingai nuo ribozės, dezoksiribozėje nėra vienos iš hidroksilo grupių. Dezoksiribozės žiedą sudaro keturi anglies atomai ir vienas deguonies atomas. Penktasis anglies atomas yra už žiedo ribų ir per deguonies atomą yra sujungtas su likusia fosforo rūgšties dalimi. Taip pat per deguonies atomą prie trečiojo anglies atomo prisijungia gretimo nukleotido fosforo rūgšties liekana.
Taigi vienoje DNR grandinėje gretimi nukleotidai yra susieti vienas su kitu. kovalentiniai ryšiai tarp dezoksiribozės ir fosforo rūgšties (fosfodiesterio jungtis). Susidaro fosfato-dezoksiribozės stuburas. Jai statmenai į kitą DNR grandinę nukreiptos azotinės bazės, kurios vandeniliniais ryšiais sujungtos su antrosios grandinės bazėmis.
DNR struktūra tokia, kad vandenilinių grandinių stuburai yra nukreipti skirtingomis kryptimis (sakoma „daugiakryptis“, „antiparalelinis“). Šone, kur vienas baigiasi fosforo rūgštimi, susieta su penktuoju dezoksiribozės anglies atomu, kitas baigiasi „laisvu“ trečiuoju anglies atomu. Tai yra, vienos grandinės skeletas apverstas aukštyn kojomis kitos atžvilgiu. Taigi DNR grandinių struktūroje išskiriami 5 „galai ir 3“ galai.
Replikuojant (dvigubinant) DNR, naujų grandinių sintezė visada vyksta nuo 5-ojo galo iki trečiojo, nes nauji nukleotidai gali prisijungti tik prie laisvo trečiojo galo.
Galiausiai (netiesiogiai per RNR) kas trys iš eilės DNR grandinės nukleotidai koduoja vieną baltymo aminorūgštį.
DNR molekulės sandaros atradimas įvyko 1953 metais F. Cricko ir D. Watsono darbų dėka (tam padėjo ir ankstyvi kitų mokslininkų darbai). Nors kaip Cheminė medžiaga DNR buvo žinoma jau XIX a. XX amžiaus 40-aisiais tapo aišku, kad genetinės informacijos nešėja yra DNR.
Dviguba spiralė laikoma antrine DNR molekulės struktūra. Eukariotinėse ląstelėse didžioji dalis DNR yra chromosomose, kur ji yra susieta su baltymais ir kitomis medžiagomis, taip pat yra tankesnė.
Deoksiribonukleorūgštis arba DNR yra genetinės informacijos nešėjas. Didžioji dalis ląstelėse esančios DNR yra sutelkta branduolyje. Tai yra pagrindinis chromosomų komponentas. Eukariotuose DNR taip pat randama mitochondrijose ir plastiduose. DNR sudaro mononukleotidai, kovalentiškai sujungti vienas su kitu, atstovaujantys ilgą, neišsišakojusį polimerą. Mononukleotidai, sudarantys DNR, sudaryti iš dezoksiribozės, vienos iš 4 azoto bazių (adenino, guanino, citozino ir timino) ir fosforo rūgšties liekanos. Šių mononukleotidų skaičius yra labai didelis. Pavyzdžiui, prokariotinėse ląstelėse, kuriose yra viena chromosoma, DNR yra viena makromolekulė, kurios molekulinė masė didesnė nei 2 x 10 9.
Vienos DNR grandinės mononukleotidai dėl susidarymo yra sujungti vienas su kitu nuosekliai kovalentiniai fosfodiesterio ryšiai tarp vieno mononukleotido dezoksiribozės OH grupės ir kito fosforo rūgšties liekanos. Vienoje suformuoto vienos DNR grandinės stuburo pusėje yra azoto bazės. Juos galima palyginti su keturiais skirtingais karoliukais, dėvimais ant vieno siūlo, nes atrodo, kad jie suverti ant cukraus-fosfato grandinės.
Kyla klausimas, kaip ši ilga polinukleotidų grandinė gali užkoduoti ląstelės ar net viso organizmo vystymosi programą? Atsakymą į šį klausimą galima gauti supratus, kaip formuojasi DNR erdvinė struktūra. Šios molekulės struktūrą iššifravo ir aprašė J. Watson ir F. Crick 1953 m.
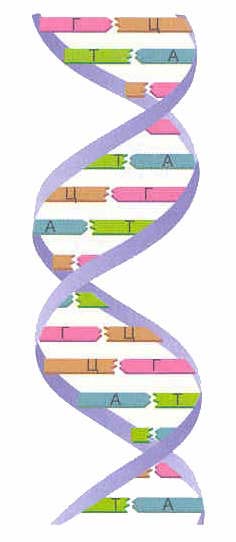
DNR molekulės yra dvi grandinės, kurios eina lygiagrečiai viena kitai ir susidaro dešiniarankė spiralė
... Šios spiralės plotis yra apie 2 nm, tačiau jos ilgis gali siekti šimtus tūkstančių nanometrų. Watsonas ir Crickas pasiūlė DNR modelį, pagal kurį visos DNR bazės yra spiralės viduje, o išorėje yra cukraus-fosfato pagrindas. Taigi vienos grandinės pagrindai yra kuo arčiau kitos grandinės pagrindų,
todėl tarp jų susidaro vandeniliniai ryšiai. DNR spiralės struktūra yra tokia, kad ją sudarančios polinukleotidinės grandinės gali būti atskirtos tik ją išvyniojus.
Dėl maksimalaus dviejų DNR grandžių artumo jo sudėtyje yra vienodas kiekis vieno tipo azoto bazių (adenino ir guanino) ir kito tipo azoto bazių (timino ir citozino), t. y. formulė galioja: A + G = T + C... Taip yra dėl azoto bazių dydžio, ty struktūrų, susidarančių dėl vandenilio jungties tarp adenino-timino ir guanino-citozino porų, ilgis yra maždaug 1,1 nm. Bendri šių porų dydžiai atitinka vidinės DNR spiralės dalies dydžius. Kad susidarytų spiralė pora C-T būtų per mažas ir pora A-G priešingai – jis per didelis. Tai yra, pirmosios DNR grandinės azotinė bazė apibrėžia bazę, esančią toje pačioje vietoje ant kitos DNR grandinės. Buvo vadinamas griežtas nukleotidų, esančių DNR molekulėje, porinėse grandinėse, lygiagrečiose viena kitai atitikmuo. papildomumo (papildomumas). Tikslus atgaminimas arba replikacija genetinė informacija yra įmanoma būtent dėl šios DNR molekulės ypatybės.
DNR biologinė informacija įrašoma taip, kad ją būtų galima tiksliai nukopijuoti ir perduoti palikuonims. Prieš joje vykstant ląstelių dalijimuisi replikacija (savęs padvigubėjimas ) DNR. Kadangi kiekvienoje grandinėje yra nukleotidų seka, kuri papildo partnerių grandinės seką, jie iš tikrųjų turi tą pačią genetinę informaciją. Jei atskirsite sruogas ir naudosite kiekvieną iš jų kaip šabloną (šabloną), kad sukurtumėte antrąją grandinę, gausite dvi naujas identiškas DNR grandines. Taip ląstelėje vyksta DNR dubliavimasis.


