რისგან შედგება დნმ. ნუკლეოტიდი - რა არის ეს? ნუკლეოტიდების შემადგენლობა, სტრუქტურა, რაოდენობა და თანმიმდევრობა დნმ-ის ჯაჭვში დნმ-ის ორმაგი სპირალში სტაბილიზირებულია იონებით.
ნუკლეინის მჟავები არის მაკრომოლეკულური ნივთიერებები, რომლებიც შედგება მონონუკლეოტიდებისგან, რომლებიც დაკავშირებულია ერთმანეთთან პოლიმერული ჯაჭვით 3", 5" - ფოსფოდიესტერული ბმების გამოყენებით და გარკვეული გზით შეფუთულია უჯრედებში.
ნუკლეინის მჟავები არის ორი ჯიშის ბიოპოლიმერები: რიბონუკლეინის მჟავა (რნმ) და დეზოქსირიბონუკლეინის მჟავა (დნმ). თითოეული ბიოპოლიმერი შედგება ნუკლეოტიდებისგან, რომლებიც განსხვავდებიან ნახშირწყლების ნარჩენებით (რიბოზა, დეზოქსირიბოზა) და ერთ-ერთი აზოტოვანი ბაზა (ურაცილი, თიმინი). შესაბამისად, ნუკლეინის მჟავებმა მიიღო სახელი.
დეზოქსირიბონუკლეინის მჟავის სტრუქტურა
ნუკლეინის მჟავებს აქვთ პირველადი, მეორადი და მესამეული სტრუქტურები.
დნმ-ის პირველადი სტრუქტურა
დნმ-ის პირველადი სტრუქტურა არის ხაზოვანი პოლინუკლეოტიდური ჯაჭვი, რომელშიც მონონუკლეოტიდები დაკავშირებულია 3", 5" ფოსფოდიესტერული ბმებით. უჯრედში ნუკლეინის მჟავის ჯაჭვის აწყობის საწყისი მასალაა ნუკლეოზიდი 5'-ტრიფოსფატი, რომელსაც ფოსფორმჟავას β და γ ნარჩენების მოცილების შედეგად შეუძლია სხვა ნუკლეოზიდის 3'-ნახშირბადის ატომის მიმაგრება. . ამრიგად, ერთი დეზოქსირიბოზის 3" ნახშირბადის ატომი კოვალენტურად უერთდება მეორე დეზოქსირიბოზის 5" ნახშირბადის ატომს ერთი ფოსფორის მჟავის ნარჩენის მეშვეობით და ქმნის ნუკლეინის მჟავას ხაზოვან პოლინუკლეოტიდურ ჯაჭვს. აქედან მოდის სახელწოდება: 3", 5"-ფოსფოდიესტერული ბმები. აზოტოვანი ფუძეები არ მონაწილეობენ ერთი ჯაჭვის ნუკლეოტიდების შეერთებაში (ნახ. 1.).
ასეთი კავშირი, ერთი ნუკლეოტიდის ფოსფორმჟავას ნარჩენებსა და მეორის ნახშირწყლებს შორის, იწვევს პოლინუკლეოტიდის მოლეკულის პენტოზა-ფოსფატის ხერხემლის წარმოქმნას, რომელზედაც გვერდიდან ერთმანეთის მიყოლებით ემატება აზოტოვანი ფუძეები. მათი თანმიმდევრობა ნუკლეინის მჟავას მოლეკულების ჯაჭვებში მკაცრად სპეციფიკურია სხვადასხვა ორგანიზმის უჯრედებისთვის, ე.ი. აქვს სპეციფიკური ხასიათი (ჩარგაფის წესი).
ხაზოვანი დნმ-ის ჯაჭვს, რომლის სიგრძე დამოკიდებულია ჯაჭვში შემავალი ნუკლეოტიდების რაოდენობაზე, აქვს ორი ბოლო: ერთს ეწოდება მე-3 ბოლო და შეიცავს თავისუფალ ჰიდროქსილს, ხოლო მეორე, მეხუთე ბოლო შეიცავს ფოსფორის მჟავას. ნარჩენი. წრე პოლარულია და შეიძლება იყოს 5"->3" და 3"->5". გამონაკლისი არის წრიული დნმ.
დნმ-ის გენეტიკური „ტექსტი“ შედგება კოდი „სიტყვებისგან“ – ნუკლეოტიდების სამეულისგან, რომელსაც კოდონები ეწოდება. დნმ-ის სეგმენტებს, რომლებიც შეიცავს ინფორმაციას ყველა ტიპის რნმ-ის პირველადი სტრუქტურის შესახებ, ეწოდება სტრუქტურული გენები.
პოლინუკლეოდიტური დნმ-ის ჯაჭვები აღწევს გიგანტურ ზომებს, ამიტომ ისინი გარკვეულწილად შეფუთულია უჯრედში.
დნმ-ის შემადგენლობის შესწავლისას ჩარგაფმა (1949) დაადგინა მნიშვნელოვანი კანონზომიერებები ცალკეული დნმ-ის ბაზების შემცველობასთან დაკავშირებით. მათ დაეხმარნენ დნმ-ის მეორადი სტრუქტურის აღმოჩენაში. ამ შაბლონებს ჩარგაფის წესებს უწოდებენ. ჩარგაფის წესები
ეს წესები ამბობს, რომ დნმ-ის აგებისას საკმაოდ მკაცრი კორესპონდენცია (დაწყვილება) უნდა იყოს დაცული არა ზოგადად პურინისა და პირიმიდინის ბაზებისთვის, არამედ კონკრეტულად თიმინისთვის ადენინთან და ციტოზინთან გუანინთან. ამ წესების საფუძველზე, სხვა საკითხებთან ერთად, 1953 წელს უოტსონმა და კრიკმა შემოგვთავაზეს დნმ-ის მეორადი სტრუქტურის მოდელი, რომელსაც ეწოდება ორმაგი სპირალი (ნახ.). |

დნმ-ის მეორადი სტრუქტურა
დნმ-ის მეორადი სტრუქტურა არის ორმაგი სპირალი, რომლის მოდელი შემოგვთავაზეს დ. უოტსონმა და ფ. კრიკმა 1953 წელს.
დნმ მოდელის შექმნის წინაპირობები
პირველადი ანალიზების შედეგად, იდეა იყო, რომ ნებისმიერი წარმოშობის დნმ შეიცავს ოთხივე ნუკლეოტიდს თანაბარი მოლარული რაოდენობით. თუმცა 1940-იან წლებში ე.ჩარგაფმა და მისმა კოლეგებმა, სხვადასხვა ორგანიზმისგან იზოლირებული დნმ-ის ანალიზის შედეგად, ნათლად აჩვენეს, რომ მათში აზოტოვანი ფუძეები სხვადასხვა რაოდენობრივი თანაფარდობითაა შეტანილი. ჩარგაფმა აღმოაჩინა, რომ მიუხედავად იმისა, რომ ეს თანაფარდობები დნმ-ისთვის ერთნაირია ერთი და იგივე სახეობის ორგანიზმების ყველა უჯრედიდან, სხვადასხვა სახეობის დნმ შეიძლება მკვეთრად განსხვავდებოდეს გარკვეული ნუკლეოტიდების შემცველობით. ეს ვარაუდობს, რომ აზოტოვანი ფუძეების თანაფარდობის განსხვავებები შესაძლოა დაკავშირებული იყოს ზოგიერთ ბიოლოგიურ კოდთან. მიუხედავად იმისა, რომ დნმ-ის სხვადასხვა ნიმუშებში ცალკეული პურინისა და პირიმიდინის ბაზების თანაფარდობა არ იყო იგივე, ანალიზის შედეგების შედარებისას გამოვლინდა გარკვეული ნიმუში: ყველა ნიმუშში პურინების მთლიანი რაოდენობა უდრიდა პირიმიდინების მთლიან რაოდენობას ( A + G = T + C), ადენინის რაოდენობა უდრიდა თიმინის რაოდენობას (A = T), ხოლო გუანინის რაოდენობა - ციტოზინის რაოდენობას (G = C). ძუძუმწოვრების უჯრედებიდან იზოლირებული დნმ ზოგადად უფრო მდიდარი იყო ადენინით და თიმინით და შედარებით ღარიბი გუანინითა და ციტოზინით, ხოლო ბაქტერიების დნმ უფრო მდიდარი იყო გუანინით და ციტოზინით და შედარებით ღარიბი ადენინითა და თიმინით. ეს მონაცემები შეადგენდა ფაქტობრივი მასალის მნიშვნელოვან ნაწილს, რომლის საფუძველზეც მოგვიანებით აშენდა უოტსონ-კრიკის დნმ-ის სტრუქტურის მოდელი.
დნმ-ის შესაძლო სტრუქტურის კიდევ ერთი მნიშვნელოვანი არაპირდაპირი მითითება იყო ლ. პაულინგის მონაცემები ცილის მოლეკულების სტრუქტურის შესახებ. პაულინგმა აჩვენა, რომ ცილის მოლეკულაში შესაძლებელია ამინომჟავის ჯაჭვის რამდენიმე განსხვავებული სტაბილური კონფიგურაცია. პეპტიდური ჯაჭვის ერთ-ერთი საერთო კონფიგურაცია - α-ჰელიქსი - არის რეგულარული ხვეული სტრუქტურა. ასეთი სტრუქტურით შესაძლებელია წყალბადის ბმების ფორმირება ამინომჟავებს შორის, რომლებიც მდებარეობს ჯაჭვის მიმდებარე მოხვევებზე. პაულინგმა აღწერა პოლიპეპტიდური ჯაჭვის α-სპირალური კონფიგურაცია 1950 წელს და ვარაუდობს, რომ დნმ-ის მოლეკულებს ასევე აქვთ სპირალური სტრუქტურა, რომელიც ფიქსირდება წყალბადის ბმებით.
თუმცა, ყველაზე ღირებული ინფორმაცია დნმ-ის მოლეკულის სტრუქტურის შესახებ რენტგენის დიფრაქციული ანალიზის შედეგებმა მოგვაწოდა. რენტგენის სხივები, რომლებიც გადის დნმ-ის კრისტალში, განიცდის დიფრაქციას, ანუ ისინი გადახრილია გარკვეული მიმართულებით. სხივების გადახრის ხარისხი და ბუნება დამოკიდებულია თავად მოლეკულების სტრუქტურაზე. რენტგენის დიფრაქციის ნიმუში (ნახ. 3) გამოცდილ თვალს აძლევს რიგ ირიბ მითითებებს შესასწავლი ნივთიერების მოლეკულების სტრუქტურასთან დაკავშირებით. დნმ-ის რენტგენის დიფრაქციული შაბლონების ანალიზმა მიგვიყვანა დასკვნამდე, რომ აზოტოვანი ფუძეები (ბრტყელი ფორმის მქონე) დაწყობილია როგორც ფირფიტების დასტა. რენტგენის ნიმუშებმა შესაძლებელი გახადა კრისტალური დნმ-ის სტრუქტურაში სამი ძირითადი პერიოდის იდენტიფიცირება: 0.34, 2 და 3.4 ნმ.

უოტსონ-კრიკის დნმ მოდელი
ჩარგაფის ანალიტიკური მონაცემებიდან დაწყებული, უილკინსის რენტგენი და ქიმიკოსის კვლევა, რომელიც გვაწვდიდა ინფორმაციას მოლეკულაში ატომებს შორის ზუსტი მანძილის, მოცემული ატომის ობლიგაციებს შორის კუთხეების და ატომების ზომის შესახებ, უოტსონი და კრიკი. დაიწყო დნმ-ის მოლეკულის ცალკეული კომპონენტების ფიზიკური მოდელების აგება გარკვეული მასშტაბით და „მორგება“ ერთმანეთზე ისე, რომ შედეგად მიღებული სისტემა შეესაბამებოდეს სხვადასხვა ექსპერიმენტულ მონაცემებს. [ჩვენება] .
ადრეც ცნობილი იყო, რომ დნმ-ის ჯაჭვში მიმდებარე ნუკლეოტიდები დაკავშირებულია ფოსფოდიესტერის ხიდებით, რომლებიც აკავშირებენ ერთი ნუკლეოტიდის დეზოქსირიბოზის 5'-ნახშირბადის ატომს მომდევნო ნუკლეოტიდის დეზოქსირიბოზის 3'-ნახშირბადის ატომთან. უოტსონსა და კრიკს ეჭვი არ ეპარებოდათ, რომ 0,34 ნმ პერიოდი შეესაბამება დნმ-ის ჯაჭვში თანმიმდევრულ ნუკლეოტიდებს შორის მანძილს. გარდა ამისა, შეიძლება ვივარაუდოთ, რომ 2 ნმ პერიოდი შეესაბამება ჯაჭვის სისქეს. და იმისთვის, რომ აეხსნათ რა რეალურ სტრუქტურას შეესაბამება 3,4 ნმ პერიოდი, უოტსონმა და კრიკმა, ისევე როგორც პაულინგმა, ადრე ჩათვალეს, რომ ჯაჭვი ხვეულია სპირალის სახით (ან, უფრო ზუსტად, ქმნის სპირალს, ვინაიდან სპირალი, ამის მკაცრი გაგებით, სიტყვა მიიღება მაშინ, როდესაც შემობრუნებები ქმნიან კონუსურ და არა ცილინდრულ ზედაპირს სივრცეში). მაშინ პერიოდი 3,4 ნმ შეესაბამება ამ სპირალის თანმიმდევრულ შემობრუნებებს შორის მანძილს. ასეთი სპირალი შეიძლება იყოს ძალიან მკვრივი ან გარკვეულწილად დაჭიმული, ანუ მისი მოხვევები შეიძლება იყოს ბრტყელი ან ციცაბო. ვინაიდან 3,4 ნმ პერიოდი ზუსტად 10-ჯერ აღემატება თანმიმდევრულ ნუკლეოტიდებს შორის მანძილს (0,34 ნმ), ცხადია, რომ სპირალის ყოველი სრული შემობრუნება შეიცავს 10 ნუკლეოტიდს. ამ მონაცემებიდან უოტსონმა და კრიკმა შეძლეს გამოთვალონ 2 ნმ დიამეტრის სპირალში გადაბმული პოლინუკლეოტიდური ჯაჭვის სიმკვრივე, მოხვევებს შორის მანძილი 3,4 ნმ უდრის. გაირკვა, რომ ასეთ ჯაჭვს ექნება დნმ-ის რეალური სიმკვრივის ნახევარი, რაც უკვე ცნობილი იყო. უნდა ვივარაუდო, რომ დნმ-ის მოლეკულა შედგება ორი ჯაჭვისგან – რომ ეს არის ნუკლეოტიდების ორმაგი სპირალი.
შემდეგი ამოცანა, რა თქმა უნდა, იყო სივრცითი ურთიერთობის გარკვევა ორ ძაფს შორის, რომლებიც ქმნიან ორმაგ სპირალს. უოტსონმა და კრიკმა თავიანთ ფიზიკურ მოდელზე სცადეს რიგი სტრიქონების მოწყობა, აღმოაჩინეს, რომ ყველა ხელმისაწვდომი მონაცემისთვის საუკეთესოა ის, რომელშიც ორი პოლინუკლეოტიდური სპირალი ეშვება საპირისპირო მიმართულებით; ამ შემთხვევაში, შაქრისა და ფოსფატის ნარჩენებისგან შემდგარი ჯაჭვები ქმნიან ორმაგი სპირალის ზედაპირს, ხოლო შიგნით განლაგებულია პურინები და პირიმიდინები. ერთმანეთის მოპირდაპირედ განლაგებული ფუძეები, რომლებიც მიეკუთვნება ორ ჯაჭვს, წყვილ-წყვილად არის დაკავშირებული წყალბადის ბმებით; ეს არის წყალბადის ბმები, რომლებიც ატარებენ ჯაჭვებს ერთად, რითაც აფიქსირებს მოლეკულის საერთო კონფიგურაციას.
დნმ-ის ორმაგი სპირალი შეიძლება მივიჩნიოთ, როგორც სპირალური თოკის კიბე, საფეხურები რჩება ჰორიზონტალურად. შემდეგ ორი გრძივი თოკი შეესაბამება შაქრისა და ფოსფატის ნარჩენების ჯაჭვებს, ხოლო ჯვარედინი ზოლები შეესაბამება წყალბადის ბმებით დაკავშირებულ აზოტოვან ფუძეებს.
შესაძლო მოდელების შემდგომი შესწავლის შედეგად, უოტსონმა და კრიკმა მივიდნენ დასკვნამდე, რომ თითოეული „ჯვარედინი ზოლი“ უნდა შედგებოდეს ერთი პურინისა და ერთი პირიმიდინისგან; 2 ნმ პერიოდის განმავლობაში (შეესაბამება ორმაგი სპირალის დიამეტრს), არ იქნებოდა საკმარისი ადგილი ორი პურინისთვის და ორი პირიმიდინი ვერ იქნებოდა ერთმანეთთან საკმარისად ახლოს, რომ შექმნან სათანადო წყალბადის ბმები. დეტალური მოდელის სიღრმისეულმა შესწავლამ აჩვენა, რომ ადენინი და ციტოზინი, რომლებიც ქმნიან სწორი ზომის კომბინაციას, მაინც ვერ მდებარეობენ ისე, რომ წყალბადის ბმები წარმოიქმნას მათ შორის. მსგავსმა მოხსენებებმა ასევე აიძულა გუანინ-თიმინის კომბინაცია გამორიცხულიყო, ხოლო ადენინ-თიმინისა და გუანინ-ციტოზინის კომბინაციები საკმაოდ მისაღები აღმოჩნდა. წყალბადის ბმების ბუნება ისეთია, რომ ადენინი წყვილდება თიმინთან, ხოლო გუანინი ციტოზინთან. სპეციფიური ბაზის დაწყვილების კონცეფციამ შესაძლებელი გახადა აეხსნა „ჩარგაფის წესი“, რომლის მიხედვითაც დნმ-ის ნებისმიერ მოლეკულაში ადენინის რაოდენობა ყოველთვის ტოლია თიმინის შემცველობასთან, ხოლო გუანინის რაოდენობა ყოველთვის ციტოზინის ტოლია. . ორი წყალბადის ბმა იქმნება ადენინსა და თიმინს შორის, ხოლო სამი გუანინსა და ციტოზინს შორის. ამ სპეციფიკის გამო ერთ ჯაჭვში თითოეული ადენინის მიმართ წყალბადური ბმების წარმოქმნაში, თიმინი მეორეშია; ანალოგიურად, მხოლოდ ციტოზინი შეიძლება განთავსდეს თითოეული გუანინის წინააღმდეგ. ამრიგად, ჯაჭვები ერთმანეთს ავსებენ, ანუ ნუკლეოტიდების თანმიმდევრობა ერთ ჯაჭვში ცალსახად განსაზღვრავს მათ თანმიმდევრობას მეორეში. ორი ჯაჭვი ეშვება საპირისპირო მიმართულებით და მათი ფოსფატის ბოლო ჯგუფები ორმაგი სპირალის საპირისპირო ბოლოებზეა.
მათი კვლევის შედეგად, 1953 წელს უოტსონმა და კრიკმა შემოგვთავაზეს დნმ-ის მოლეკულის სტრუქტურის მოდელი (ნახ. 3), რომელიც აქტუალურია დღემდე. მოდელის მიხედვით, დნმ-ის მოლეკულა შედგება ორი დამატებითი პოლინუკლეოტიდური ჯაჭვისგან. დნმ-ის თითოეული ჯაჭვი არის პოლინუკლეოტიდი, რომელიც შედგება რამდენიმე ათი ათასი ნუკლეოტიდისგან. მასში მეზობელი ნუკლეოტიდები ქმნიან რეგულარულ პენტოზა-ფოსფატის ხერხემალს ფოსფორის მჟავის ნარჩენისა და დეზოქსირიბოზის ძლიერი კოვალენტური ბმის კომბინაციის გამო. ერთი პოლინუკლეოტიდური ჯაჭვის აზოტოვანი ფუძეები განლაგებულია მკაცრად განსაზღვრული თანმიმდევრობით მეორის აზოტოვანი ფუძეების წინააღმდეგ. პოლინუკლეოტიდურ ჯაჭვში აზოტოვანი ფუძეების მონაცვლეობა არარეგულარულია.
დნმ-ის ჯაჭვში აზოტოვანი ფუძეების განლაგება ურთიერთშემავსებელია (ბერძნული „კომპლიმენტიდან“ - დამატება), ე.ი. ადენინის წინააღმდეგ (A) ყოველთვის არის თიმინი (T), ხოლო გუანინის (G) წინააღმდეგ - მხოლოდ ციტოზინი (C). ეს აიხსნება იმით, რომ A და T, ისევე როგორც G და C, მკაცრად შეესაბამება ერთმანეთს, ე.ი. ავსებენ ერთმანეთს. ეს შესაბამისობა მოცემულია ფუძეების ქიმიური სტრუქტურით, რაც იძლევა წყალბადის ბმების წარმოქმნის საშუალებას პურინისა და პირიმიდინის წყვილში. A-სა და T-ს შორის არის ორი ბმა, G-სა და C-ს შორის - სამი. ეს ობლიგაციები უზრუნველყოფს დნმ-ის მოლეკულის ნაწილობრივ სტაბილიზაციას სივრცეში. ორმაგი სპირალის სტაბილურობა პირდაპირპროპორციულია G≡C ბმების რაოდენობასთან, რომლებიც უფრო სტაბილურია ვიდრე A=T ბმები.
ნუკლეოტიდების ცნობილი თანმიმდევრობა დნმ-ის ერთ ჯაჭვში შესაძლებელს ხდის სხვა ჯაჭვის ნუკლეოტიდების დადგენას, კომპლემენტარობის პრინციპით.
გარდა ამისა, დადგინდა, რომ არომატული სტრუქტურის მქონე აზოტოვანი ფუძეები განლაგებულია ერთმანეთის ზემოთ წყალხსნარში და ქმნიან, თითქოსდა, მონეტების დასტას. ეს პროცესი ფორმირების stacks of ორგანული მოლეკულებიდაწყობას უწოდებენ. განხილული უოტსონ-კრიკის მოდელის დნმ-ის მოლეკულის პოლინუკლეოტიდურ ჯაჭვებს აქვთ მსგავსი ფიზიკოქიმიური მდგომარეობა, მათი აზოტოვანი ფუძეები განლაგებულია მონეტების დასის სახით, რომელთა სიბრტყეებს შორის ხდება ვან დერ ვაალის ურთიერთქმედება (დაწყობის ურთიერთქმედება).
წყალბადის ბმა კომპლემენტურ ფუძეებს შორის (ჰორიზონტალურად) და პოლინუკლეოტიდურ ჯაჭვში ბაზის სიბრტყეებს შორის დაწყობილი ურთიერთქმედება ვან დერ ვაალის ძალების გამო (ვერტიკალურად) უზრუნველყოფს დნმ-ის მოლეკულას დამატებით სტაბილიზაციას სივრცეში.
ორივე ჯაჭვის შაქრის-ფოსფატის ხერხემალი მობრუნებულია გარედან, ფუძეები კი შიგნით, ერთმანეთისკენ. დნმ-ში ჯაჭვების მიმართულება ანტიპარალელულია (ერთ მათგანს აქვს მიმართულება 5"->3", მეორეს - 3"->5", ანუ ერთი ჯაჭვის 3"-ბოლო მდებარეობს 5"-ბოლოს მოპირდაპირედ. მეორის.). ჯაჭვები ქმნიან მარჯვენა სპირალებს საერთო ღერძით. სპირალის ერთი შემობრუნება არის 10 ნუკლეოტიდი, ბრუნვის ზომაა 3,4 ნმ, თითოეული ნუკლეოტიდის სიმაღლეა 0,34 ნმ, სპირალის დიამეტრი 2,0 ნმ. ერთი ჯაჭვის მეორის გარშემო ბრუნვის შედეგად დნმ-ის ორმაგ სპირალში წარმოიქმნება ძირითადი ღარი (დაახლოებით 20 Å დიამეტრის) და მცირე ღარი (დაახლოებით 12 Å). უოტსონ-კრიკის ორმაგი სპირალის ამ ფორმას მოგვიანებით B- ფორმა ეწოდა. უჯრედებში დნმ ჩვეულებრივ არსებობს B ფორმაში, რომელიც ყველაზე სტაბილურია.
დნმ-ის ფუნქციები
შემოთავაზებულმა მოდელმა ახსნა დეზოქსირიბონუკლეინის მჟავას მრავალი ბიოლოგიური თვისება, მათ შორის გენეტიკური ინფორმაციის შენახვა და გენების მრავალფეროვნება, რაც უზრუნველყოფილია 4 ნუკლეოტიდის თანმიმდევრული კომბინაციით და გენეტიკური კოდის არსებობის ფაქტი, უნარი გენეტიკური ინფორმაციის თვითრეპროდუცირება და გადაცემა, რომელიც მოწოდებულია რეპლიკაციის პროცესით და გენეტიკური ინფორმაციის დანერგვა ცილების სახით, ისევე როგორც ნებისმიერი სხვა ნაერთები, რომლებიც წარმოიქმნება ფერმენტული ცილების დახმარებით.
დნმ-ის ძირითადი ფუნქციები.
- დნმ არის გენეტიკური ინფორმაციის მატარებელი, რასაც გენეტიკური კოდის არსებობის ფაქტი უზრუნველყოფს.
- რეპროდუქცია და გადაცემული გენეტიკური ინფორმაცია უჯრედებისა და ორგანიზმების თაობებში. ეს ფუნქცია უზრუნველყოფილია რეპლიკაციის პროცესით.
- გენეტიკური ინფორმაციის დანერგვა ცილების სახით, ისევე როგორც ნებისმიერი სხვა ნაერთები, რომლებიც წარმოიქმნება ფერმენტის ცილების დახმარებით. ამ ფუნქციას უზრუნველყოფს ტრანსკრიფციის და თარგმანის პროცესები.

ორჯაჭვიანი დნმ-ის ორგანიზაციის ფორმები
დნმ-ს შეუძლია შექმნას რამდენიმე ტიპის ორმაგი სპირალი (ნახ. 4). ამჟამად უკვე ცნობილია ექვსი ფორმა (A-დან E-მდე და Z-ფორმამდე).
როზალინდ ფრანკლინის მიერ დადგენილი დნმ-ის სტრუქტურული ფორმები დამოკიდებულია ნუკლეინის მჟავის მოლეკულის წყლით გაჯერებაზე. რენტგენის დიფრაქციული ანალიზის გამოყენებით დნმ-ის ბოჭკოების კვლევებში აჩვენეს, რომ რენტგენის დიფრაქციის ნიმუში რადიკალურად დამოკიდებულია იმაზე, თუ რა ფარდობით ტენიანობაზე, ამ ბოჭკოს წყლის გაჯერების რა ხარისხზე ტარდება ექსპერიმენტი. თუ ბოჭკო საკმარისად იყო გაჯერებული წყლით, მაშინ მიიღეს ერთი რენტგენოგრაფია. გაშრობისას გამოჩნდა სრულიად განსხვავებული რენტგენის ნიმუში, რომელიც ძალიან განსხვავდება მაღალი ტენიანობის ბოჭკოს რენტგენის ნიმუშისგან.
მაღალი ტენიანობის დნმ-ის მოლეკულას B- ფორმა ეწოდება. ფიზიოლოგიურ პირობებში (მარილების დაბალი კონცენტრაცია, დატენიანების მაღალი ხარისხი) დნმ-ის დომინანტური სტრუქტურული ტიპია B- ფორმა (ორჯაჭვიანი დნმ-ის ძირითადი ფორმა არის Watson-Crick მოდელი). ასეთი მოლეკულის სპირალის სიმაღლეა 3,4 ნმ. თითო ბრუნვაში არის 10 დამატებითი წყვილი "მონეტების" დაგრეხილი დასტაების სახით - აზოტოვანი ფუძეები. დასტაები ერთმანეთთან არის შეკრული წყალბადის ობლიგაციებით ორ საპირისპირო „მონეტას“ შორის და „დახვეულია“ ფოსფოდიესტერის ხერხემლის ორი ლენტით, გადაბმული მარჯვენა სპირალში. აზოტოვანი ფუძეების სიბრტყეები პერპენდიკულარულია სპირალის ღერძის მიმართ. მეზობელი დამატებითი წყვილი ბრუნავს ერთმანეთთან შედარებით 36°-ით. სპირალის დიამეტრი არის 20Å, პურინის ნუკლეოტიდი იკავებს 12Å და პირიმიდინის ნუკლეოტიდს იკავებს 8Å.
ქვედა ტენიანობის დნმ-ის მოლეკულას A- ფორმა ეწოდება. A- ფორმა წარმოიქმნება ნაკლებად მაღალი ჰიდრატაციის პირობებში და Na + ან K + იონების მაღალი შემცველობით. ამ უფრო ფართო მარჯვენა კონფორმაციას აქვს 11 ბაზის წყვილი თითო შემობრუნებაზე. აზოტოვანი ფუძეების სიბრტყეებს უფრო ძლიერი დახრილობა აქვთ სპირალის ღერძის მიმართ, ისინი გადახრილია ნორმალურიდან სპირალის ღერძზე 20°-ით. ეს გულისხმობს შიდა სიცარიელის არსებობას 5 Å დიამეტრით. მიმდებარე ნუკლეოტიდებს შორის მანძილი არის 0,23 ნმ, ხვეულის სიგრძე 2,5 ნმ, ხოლო სპირალის დიამეტრი 2,3 ნმ.
თავდაპირველად, დნმ-ის A-ფორმა ნაკლებად მნიშვნელოვანი იყო. თუმცა, მოგვიანებით გაირკვა, რომ დნმ-ის A-ფორმას, ისევე როგორც B- ფორმას, დიდი ბიოლოგიური მნიშვნელობა აქვს. რნმ-დნმ-ის სპირალს შაბლონ-თესლის კომპლექსში აქვს A-ფორმა, ასევე რნმ-რნმ სპირალი და რნმ თმის სამაგრი სტრუქტურები (რიბოზის 2'-ჰიდროქსილის ჯგუფი არ აძლევს რნმ-ის მოლეკულებს B-ფორმის შექმნის საშუალებას). . დნმ-ის A-ფორმა გვხვდება სპორებში. დადგენილია, რომ დნმ-ის A ფორმა 10-ჯერ უფრო მდგრადია ულტრაიისფერი სხივების მიმართ, ვიდრე B ფორმა.
A-ფორმა და B-ფორმა ეწოდება კანონიკური ფორმებიდნმ.
ფორმები C-Eასევე მემარჯვენეები, მათი ფორმირება მხოლოდ სპეციალურ ექსპერიმენტებში შეიძლება დაფიქსირდეს და, როგორც ჩანს, ისინი არ არსებობენ in vivo. დნმ-ის C- ფორმას აქვს B-დნმ-ის მსგავსი სტრუქტურა. ბაზის წყვილების რაოდენობა ერთ შემობრუნებაზე არის 9,33, ხოლო სპირალის სიგრძე 3,1 ნმ. ბაზის წყვილები დახრილია 8 გრადუსიანი კუთხით ღერძის პერპენდიკულარულ პოზიციასთან შედარებით. ღარები ზომით ახლოსაა B-დნმ-ის ღარებთან. ამ შემთხვევაში, მთავარი ღარი გარკვეულწილად უფრო მცირეა, ხოლო მცირე ღარი უფრო ღრმაა. ბუნებრივი და სინთეზური დნმ-ის პოლინუკლეოტიდები შეიძლება გადავიდნენ C- ფორმაში.
| ცხრილი 1. დნმ-ის ზოგიერთი ტიპის სტრუქტურის მახასიათებლები | |||
| სპირალური ტიპი | ა | ბ | ზ |
| სპირალური მოედანი | 0,32 ნმ | 3,38 ნმ | 4,46 ნმ |
| სპირალის გადახვევა | უფლება | უფლება | მარცხენა |
| ბაზის წყვილების რაოდენობა თითო შემობრუნებაზე | 11 | 10 | 12 |
| მანძილი საბაზისო თვითმფრინავებს შორის | 0,256 ნმ | 0.338 ნმ | 0.371 ნმ |
| გლიკოზიდური ბმის კონფორმაცია | ანტი | ანტი | ანტი-C სინ-გ |
| ფურანოზის რგოლის კონფორმაცია | C3 "-ენდო | C2 "-ენდო | C3 "-endo-G C2 "-endo-C |
| ღარის სიგანე, პატარა/დიდი | 1.11/0.22 ნმ | 0,57/1,17 ნმ | 0.2/0.88 ნმ |
| ღარის სიღრმე, პატარა/დიდი | 0,26/1,30 ნმ | 0,82/0,85 ნმ | 1.38/0.37 ნმ |
| სპირალის დიამეტრი | 2.3 ნმ | 2.0 ნმ | 1.8 ნმ |
დნმ-ის სტრუქტურული ელემენტები
(დნმ-ის არაკანონიკური სტრუქტურები)
დნმ-ის სტრუქტურული ელემენტები მოიცავს უჩვეულო სტრუქტურებს, რომლებიც შემოიფარგლება რამდენიმე სპეციალური თანმიმდევრობით:
|

დნმ-ის Z- ფორმააღმოაჩინეს 1979 წელს ჰექსანუკლეოტიდის d(CG)3 - შესწავლისას. იგი გახსნეს MIT პროფესორმა ალექსანდრე რიჩმა და მისმა თანამშრომლებმა. Z-ფორმა გახდა დნმ-ის ერთ-ერთი ყველაზე მნიშვნელოვანი სტრუქტურული ელემენტი იმის გამო, რომ მისი ფორმირება შეინიშნებოდა დნმ-ის რეგიონებში, სადაც პურინები მონაცვლეობენ პირიმიდინებთან (მაგალითად, 5'-HCHCHC-3'), ან გამეორებით 5'. -CHCHCH-3', რომელიც შეიცავს მეთილირებულ ციტოზინს. Z-დნმ-ის ფორმირებისა და სტაბილიზაციის აუცილებელი პირობა იყო მასში პურინის ნუკლეოტიდების არსებობა სინ-კონფორმაციაში, რომლებიც მონაცვლეობდნენ პირიმიდინის ბაზებთან ანტიკონფორმაციაში.
ბუნებრივი დნმ-ის მოლეკულები ძირითადად არსებობენ მარჯვენა B ფორმაში, თუ ისინი არ შეიცავენ თანმიმდევრობას, როგორიცაა (CG)n. თუმცა, თუ ასეთი თანმიმდევრობები დნმ-ის ნაწილია, მაშინ ეს რეგიონები, როდესაც ხსნარის ან კათიონების იონური სიძლიერე, რომელიც ანეიტრალებს ფოსფოდიესტერის ხერხემალზე უარყოფით მუხტს, შეიძლება გადავიდეს Z- ფორმაში, ხოლო ჯაჭვის სხვა დნმ-ის უბნები დარჩეს. კლასიკური B ფორმაში. ასეთი გადასვლის შესაძლებლობა მიუთითებს იმაზე, რომ დნმ-ის ორმაგი სპირალის ორი ძაფები დინამიურ მდგომარეობაშია და შეუძლიათ ერთმანეთთან შედარებით გადახვევა, მარჯვენა ფორმიდან მარცხენაზე გადასვლა და პირიქით. ამ ლაბილურობის ბიოლოგიური შედეგები, რომელიც იძლევა დნმ-ის სტრუქტურის კონფორმაციული გარდაქმნების საშუალებას, ჯერ ბოლომდე არ არის გასაგები. ითვლება, რომ Z-დნმ რეგიონები თამაშობენ როლს გარკვეული გენების ექსპრესიის რეგულირებაში და მონაწილეობენ გენეტიკურ რეკომბინაციაში.
დნმ-ის Z ფორმა არის მარცხენა ორმაგი სპირალი, რომელშიც ფოსფოდიესტერის ხერხემალი ზიგზაგია მოლეკულის ღერძის გასწვრივ. აქედან მოდის მოლეკულის (ზიგზაგი)-დნმ-ის სახელი. Z-დნმ არის ყველაზე ნაკლებად გრეხილი (12 ბაზის წყვილი თითო ბრუნზე) და ყველაზე თხელი ბუნებაში. მანძილი მეზობელ ნუკლეოტიდებს შორის არის 0,38 ნმ, ხვეულის სიგრძე 4,56 ნმ და Z-დნმ დიამეტრი 1,8 ნმ. გარდა ამისა, გარეგნობადნმ-ის ეს მოლეკულა გამოირჩევა ერთი ღარის არსებობით.
დნმ-ის Z ფორმა ნაპოვნია პროკარიოტულ და ევკარიოტულ უჯრედებში. დღეისათვის მიღებულია ანტისხეულები, რომლებსაც შეუძლიათ განასხვავონ დნმ-ის Z- და B- ფორმა. ეს ანტისხეულები უკავშირდებიან დროზოფილას (Dr. melanogaster) სანერწყვე ჯირკვლის უჯრედების გიგანტური ქრომოსომების სპეციფიკურ უბნებს. შებოჭვის რეაქცია მარტივია ამ ქრომოსომების უჩვეულო სტრუქტურის გამო, რომლებშიც უფრო მკვრივი რეგიონები (დისკები) განსხვავდება ნაკლებად მკვრივი რეგიონებისგან (ინტერდისკები). Z-დნმ რეგიონები განლაგებულია ინტერდისკებში. აქედან გამომდინარეობს, რომ Z- ფორმა რეალურად არსებობს ბუნებრივ პირობებში, თუმცა Z- ფორმის ცალკეული მონაკვეთების ზომები ჯერ არ არის ცნობილი.

(ცვლილები) - დნმ-ში ყველაზე ცნობილი და ხშირად გავრცელებული ბაზის თანმიმდევრობები. პალინდრომი არის სიტყვა ან ფრაზა, რომელიც იკითხება მარცხნიდან მარჯვნივ და პირიქით. ასეთი სიტყვების ან ფრაზების მაგალითებია: ქოხი, კაზაკი, წყალდიდობა და აზორის თათებზე დაცემული ვარდი. როდესაც გამოიყენება დნმ-ის მონაკვეთებზე, ეს ტერმინი (პალინდრომი) ნიშნავს ნუკლეოტიდების იგივე მონაცვლეობას ჯაჭვის გასწვრივ მარჯვნიდან მარცხნივ და მარცხნიდან მარჯვნივ (როგორც ასოები სიტყვაში "ქოხი" და ა.შ.).
პალინდრომს ახასიათებს დნმ-ის ორი ჯაჭვის მიმართ მეორე რიგის სიმეტრიის მქონე ბაზის მიმდევრობების ინვერსიული გამეორებების არსებობა. ასეთი თანმიმდევრობები, გასაგები მიზეზების გამო, თვითშეავსებენ და მიდრეკილნი არიან წარმოქმნან თმის სამაგრი ან ჯვარცმული სტრუქტურები (ნახ.). თმის სამაგრები ეხმარება მარეგულირებელ ცილებს ამოიცნონ ქრომოსომის დნმ-ის გენეტიკური ტექსტის კოპირება.
იმ შემთხვევებში, როდესაც ინვერსიული გამეორება არის იმავე დნმ-ის ჯაჭვში, ასეთ თანმიმდევრობას სარკის გამეორება ეწოდება. სარკის გამეორებას არ გააჩნია თვითშემავსებელი თვისებები და, შესაბამისად, არ შეუძლია შექმნას თმის სამაგრი ან ჯვარცმული სტრუქტურები. ამ ტიპის თანმიმდევრობები გვხვდება დნმ-ის თითქმის ყველა დიდ მოლეკულაში და შეიძლება მერყეობდეს რამდენიმე ბაზის წყვილიდან რამდენიმე ათას ბაზის წყვილამდე.
პალინდრომების არსებობა ჯვარცმული სტრუქტურების სახით ევკარიოტულ უჯრედებში არ არის დადასტურებული, თუმცა მთელი რიგი ჯვარცმული სტრუქტურები ნაპოვნია in vivo E. coli უჯრედებში. რნმ-ში ან ერთჯაჭვიანი დნმ-ში თვითშემავსებელი თანმიმდევრობების არსებობა არის ხსნარებში ნუკლეინის ჯაჭვის დაკეცვის მთავარი მიზეზი გარკვეულ სივრცულ სტრუქტურაში, რაც ხასიათდება მრავალი „თმის სამაგრის“ წარმოქმნით.

დნმ-ის H- ფორმა- ეს არის სპირალი, რომელიც იქმნება დნმ-ის სამი ჯაჭვისგან - დნმ-ის სამმაგი სპირალი. ეს არის უოტსონ-კრიკის ორმაგი სპირალის კომპლექსი მესამე ერთჯაჭვიანი დნმ-ის ჯაჭვით, რომელიც ჯდება მის დიდ ღარში, ე.წ. Hoogsteen წყვილის წარმოქმნით.
ასეთი ტრიპლექსის წარმოქმნა ხდება დნმ-ის ორმაგი სპირალის დამატების შედეგად ისე, რომ მისი მონაკვეთის ნახევარი რჩება ორმაგი სპირალის სახით, ხოლო მეორე ნახევარი გათიშულია. ამ შემთხვევაში, ერთ-ერთი გათიშული სპირალი ორმაგი სპირალის პირველი ნახევრით ქმნის ახალ სტრუქტურას - სამმაგი სპირალი, ხოლო მეორე აღმოჩნდება უსტრუქტურო, ერთძაფის განყოფილების სახით. ამ სტრუქტურული გადასვლის თავისებურებაა მკვეთრი დამოკიდებულება გარემოს pH-ზე, რომლის პროტონები ასტაბილურებენ ახალ სტრუქტურას. ამ მახასიათებლის გამო ახალ სტრუქტურას ეწოდა დნმ-ის H- ფორმა, რომლის ფორმირება აღმოაჩინეს ჰომოპურინ-ჰომოპირიმიდინის უბნების შემცველ ზეხვეულ პლაზმიდებში, რომლებიც სარკისებური გამეორებაა.
შემდგომ კვლევებში დადგინდა ზოგიერთი ჰომოპურინ-ჰომოპირიმიდინის ორჯაჭვიანი პოლინუკლეოტიდის სტრუქტურული გადასვლის შესაძლებლობა სამჯაჭვიანი სტრუქტურის ფორმირებით, რომელიც შეიცავს:
- ერთი ჰომოპურინი და ორი ჰომოპირიმიდინის ჯაჭვი ( Py-Pu-Py ტრიპლექსი) [ჰუგსტინის ურთიერთქმედება].
Py-Pu-Py ტრიპლექსის შემადგენელი ბლოკები არის კანონიკური იზომორფული CGC+ და TAT ტრიადები. ტრიპლექსის სტაბილიზაცია მოითხოვს CGC+ ტრიადის პროტონაციას, ამიტომ ეს ტრიპლექსები დამოკიდებულია ხსნარის pH-ზე.
- ერთი ჰომოპირიმიდინი და ორი ჰომოპურინი ( პი-პუ-პუ ტრიპლექსი) [ჰუგსტინის შებრუნებული ურთიერთქმედება].
Py-Pu-Pu ტრიპლექსის შემადგენელი ბლოკები არის კანონიკური იზომორფული CGG და TAA ტრიადები. Py-Pu-Pu ტრიპლექსების არსებითი თვისებაა მათი სტაბილურობის დამოკიდებულება ორმაგად დამუხტული იონების არსებობაზე და სხვადასხვა იონები საჭიროა სხვადასხვა მიმდევრობის ტრიპლექსების სტაბილიზაციისთვის. ვინაიდან Py-Pu-Pu ტრიპლექსების ფორმირება არ საჭიროებს მათი შემადგენელი ნუკლეოტიდების პროტონაციას, ასეთი ტრიპლექსები შეიძლება არსებობდეს ნეიტრალურ pH-ზე.
შენიშვნა: Hoogsteen-ის პირდაპირი და საპირისპირო ურთიერთქმედება აიხსნება 1-მეთილთიმინის სიმეტრიით: 180 ° ბრუნვა იწვევს იმ ფაქტს, რომ O4 ატომის ადგილი იკავებს O2 ატომს, ხოლო წყალბადის ბმების სისტემა შენარჩუნებულია.
არსებობს სამმაგი სპირალის ორი ტიპი:
- პარალელური სამმაგი სპირალი, რომელშიც მესამე ჯაჭვის პოლარობა იგივეა, რაც უოტსონ-კრიკის დუპლექსის ჰომოპურინის ჯაჭვის.
- ანტიპარალელური სამმაგი სპირალი, რომელშიც მესამე და ჰომოპურინის ჯაჭვების პოლარობები საპირისპიროა.

G- ოთხკუთხედი- 4-ჯაჭვიანი დნმ. ასეთი სტრუქტურა იქმნება, თუ არსებობს ოთხი გუანინი, რომლებიც ქმნიან ეგრეთ წოდებულ G-quadruplex-ს - ოთხი გუანინის მრგვალ ცეკვას.
პირველი მინიშნებები ასეთი სტრუქტურების ფორმირების შესაძლებლობის შესახებ იქნა მიღებული უოტსონისა და კრიკის გარღვევამდე დიდი ხნით ადრე - ჯერ კიდევ 1910 წელს. შემდეგ გერმანელმა ქიმიკოსმა ივარ ბანგმა აღმოაჩინა, რომ დნმ-ის ერთ-ერთი კომპონენტი - გუანოზინის მჟავა - ქმნის გელებს მაღალი კონცენტრაციით, ხოლო დნმ-ის სხვა კომპონენტებს ეს თვისება არ გააჩნიათ.
1962 წელს რენტგენის დიფრაქციული მეთოდის გამოყენებით შესაძლებელი გახდა ამ გელის უჯრედული სტრუქტურის დადგენა. აღმოჩნდა, რომ იგი შედგებოდა ოთხი გუანინის ნარჩენებისგან, რომლებიც ერთმანეთს აკავშირებდნენ წრეში და ქმნიდნენ დამახასიათებელ კვადრატს. ცენტრში ბმა მხარს უჭერს ლითონის იონს (Na, K, Mg). იგივე სტრუქტურები შეიძლება ჩამოყალიბდეს დნმ-ში, თუ ის შეიცავს ბევრ გუანინს. ეს ბრტყელი კვადრატები (G-კვარტეტები) დაწყობილია საკმაოდ სტაბილური, მკვრივი სტრუქტურების შესაქმნელად (G-ოთხკუთხედები).
დნმ-ის ოთხი ცალკეული ჯაჭვი შეიძლება იყოს ნაქსოვი ოთხჯაჭვიან კომპლექსებად, მაგრამ ეს საკმაოდ გამონაკლისია. უფრო ხშირად, ნუკლეინის მჟავის ერთი ჯაჭვი უბრალოდ კვანძად არის მიბმული, რაც ქმნის დამახასიათებელ გასქელებას (მაგალითად, ქრომოსომების ბოლოებზე), ან ორჯაჭვიანი დნმ ქმნის ადგილობრივ ოთხკუთხედს ზოგიერთ გუანინით მდიდარ ადგილას.
ყველაზე შესწავლილი არის ოთხკუთხედების არსებობა ქრომოსომების ბოლოებზე - ტელომერებზე და ონკოპრომოტერებში. თუმცა, ადამიანის ქრომოსომებში ასეთი დნმ-ის ლოკალიზაციის სრული გაგება ჯერ კიდევ არ არის ცნობილი.
დნმ-ის ყველა ეს უჩვეულო სტრუქტურა ხაზოვანი ფორმით არასტაბილურია დნმ-ის B ფორმასთან შედარებით. თუმცა, დნმ ხშირად არსებობს ტოპოლოგიური დაძაბულობის რგოლის სახით, როდესაც მას აქვს ის, რაც ცნობილია, როგორც ზეგადახვევა. ამ პირობებში ადვილად ყალიბდება არაკანონიკური დნმ-ის სტრუქტურები: Z-ფორმები, „ჯვრები“ და „თმები“, H-ფორმები, გუანინის ოთხკუთხედები და i-მოტივი.
- ზეხვეული ფორმა - აღინიშნება უჯრედის ბირთვიდან გათავისუფლებისას პენტოზა-ფოსფატის ხერხემლის დაზიანების გარეშე. მას აქვს სუპერმოგრეხილი დახურული რგოლების ფორმა. ზეგადაბრუნებულ მდგომარეობაში დნმ-ის ორმაგი სპირალი ერთხელ მაინც „მიუგრიხეს თავის თავზე“, ანუ შეიცავს სულ მცირე ერთ სუპერკოჭას (რვა ფიგურის ფორმას იღებს).
- დნმ-ის მოდუნებული მდგომარეობა - შეინიშნება ერთი შესვენებით (ერთი ჯაჭვის გაწყვეტით). ამ შემთხვევაში სუპერხვეულები ქრება და დნმ დახურული რგოლის ფორმას იღებს.
- დნმ-ის წრფივი ფორმა შეინიშნება ორმაგი სპირალის ორი ჯაჭვის გაწყვეტისას.
დნმ-ის მესამეული სტრუქტურა
დნმ-ის მესამეული სტრუქტურაწარმოიქმნება ორჯაჭვიანი მოლეკულის სივრცეში დამატებითი გადახვევის - მისი ზეგადახვევის შედეგად. ეუკარიოტულ უჯრედებში დნმ-ის მოლეკულის ზეგადახვევა, პროკარიოტებისგან განსხვავებით, ხორციელდება ცილებთან კომპლექსების სახით.
თითქმის ყველა ევკარიოტული დნმ მდებარეობს ბირთვების ქრომოსომებში, მისი მხოლოდ მცირე რაოდენობაა მიტოქონდრიებში, მცენარეებში და პლასტიდებში. ევკარიოტული უჯრედების (მათ შორის ადამიანის ქრომოსომების) ქრომოსომების ძირითადი ნივთიერება არის ქრომატინი, რომელიც შედგება ორჯაჭვიანი დნმ-ის, ჰისტონისა და არაჰისტონის ცილებისგან.
ქრომატინის ჰისტონური ცილები
ჰისტონები მარტივი ცილებია, რომლებიც შეადგენენ ქრომატინის 50%-მდე. ცხოველებისა და მცენარეების ყველა შესწავლილ უჯრედში აღმოჩნდა ჰისტონების ხუთი ძირითადი კლასი: H1, H2A, H2B, H3, H4, განსხვავებული ზომით, ამინომჟავის შემადგენლობით და მუხტით (ყოველთვის დადებითი).
ძუძუმწოვრების ჰისტონი H1 შედგება ერთი პოლიპეპტიდური ჯაჭვისგან, რომელიც შეიცავს დაახლოებით 215 ამინომჟავას; სხვა ჰისტონების ზომები მერყეობს 100-დან 135 ამინომჟავამდე. ყველა მათგანი სპირალიზებულია და გრეხილია გლობულად, რომლის დიამეტრი დაახლოებით 2,5 ნმ-ია, შეიცავს უჩვეულოდ დიდი რაოდენობით დადებითად დამუხტულ ამინომჟავებს ლიზინს და არგინინს. ჰისტონები შეიძლება იყოს აცეტილირებული, მეთილირებული, ფოსფორილირებადი, პოლი(ADP)-რიბოზილირებადი, ხოლო ჰისტონები H2A და H2B შეიძლება კოვალენტურად იყოს დაკავშირებული უბიკვიტინთან. რა როლი აქვს ამგვარ მოდიფიკაციებს ჰისტონების სტრუქტურის ფორმირებასა და ფუნქციების შესრულებაში, ჯერ ბოლომდე არ არის განმარტებული. ვარაუდობენ, რომ ეს არის მათი უნარი დნმ-თან ურთიერთობისა და გენების მოქმედების რეგულირების ერთ-ერთი მექანიზმის უზრუნველყოფას.
ჰისტონები ურთიერთქმედებენ დნმ-თან ძირითადად იონური ბმების (მარილის ხიდების) მეშვეობით, რომლებიც წარმოიქმნება დნმ-ის უარყოფითად დამუხტულ ფოსფატურ ჯგუფებსა და ჰისტონების დადებითად დამუხტულ ლიზინისა და არგინინის ნარჩენებს შორის.
ქრომატინის არაჰისტონის ცილები
არაჰისტონის ცილები, ჰისტონებისგან განსხვავებით, ძალიან მრავალფეროვანია. იზოლირებულია დნმ-ის დამაკავშირებელი არაჰისტონის ცილების 590-მდე სხვადასხვა ფრაქცია. მათ ასევე უწოდებენ მჟავე პროტეინებს, რადგან მათ სტრუქტურაში ჭარბობს მჟავე ამინომჟავები (ეს არის პოლიანიონები). ქრომატინის აქტივობის სპეციფიკური რეგულირება დაკავშირებულია სხვადასხვა არაჰისტონის ცილებთან. მაგალითად, ფერმენტები, რომლებიც აუცილებელია დნმ-ის რეპლიკაციისთვის და ექსპრესიისთვის, შეუძლიათ ქრომატინს დროებით დაუკავშირონ. სხვა პროტეინები, რომლებიც მონაწილეობენ სხვადასხვა მარეგულირებელ პროცესებში, უკავშირდებიან დნმ-ს მხოლოდ კონკრეტულ ქსოვილებში ან დიფერენციაციის გარკვეულ ეტაპებზე. თითოეული ცილა ავსებს დნმ-ის ნუკლეოტიდების სპეციფიკურ თანმიმდევრობას (დნმ-ის ადგილი). ამ ჯგუფში შედის:
- საიტის სპეციფიკური თუთიის თითის ცილების ოჯახი. თითოეული „თუთიის თითი“ ცნობს კონკრეტულ ადგილს, რომელიც შედგება 5 ნუკლეოტიდური წყვილისაგან.
- უბნის სპეციფიკური ცილების ოჯახი - ჰომოდიმერები. ასეთი ცილის ფრაგმენტს დნმ-თან კონტაქტში აქვს "სპირალი-ბრუნი-სპირალი" სტრუქტურა.
- მაღალი მობილურობის პროტეინები (HMG proteins - ინგლისურიდან, მაღალი მობილურობის გელი პროტეინები) არის სტრუქტურული და მარეგულირებელი ცილების ჯგუფი, რომლებიც მუდმივად ასოცირდება ქრომატინთან. მათ აქვთ 30 კდ-ზე ნაკლები მოლეკულური წონა და ხასიათდებიან დამუხტული ამინომჟავების მაღალი შემცველობით. მათი დაბალი მოლეკულური წონის გამო, HMG ცილები ძალიან მოძრავია პოლიაკრილამიდის გელის ელექტროფორეზის დროს.
- რეპლიკაციის, ტრანსკრიფციის და აღდგენის ფერმენტები.
დნმ-ისა და რნმ-ის სინთეზში ჩართული სტრუქტურული, მარეგულირებელი ცილების და ფერმენტების მონაწილეობით, ნუკლეოსომის ძაფი გარდაიქმნება ცილებისა და დნმ-ის მაღალ შედედებულ კომპლექსად. შედეგად მიღებული სტრუქტურა 10000-ჯერ უფრო მოკლეა, ვიდრე ორიგინალური დნმ-ის მოლეკულა.

ქრომატინი
ქრომატინი არის ცილების კომპლექსი ბირთვული დნმ-ით და არაორგანული ნივთიერებები. ქრომატინის უმეტესობა არააქტიურია. ის შეიცავს მჭიდროდ შეფუთულ, შედედებულ დნმ-ს. ეს არის ჰეტეროქრომატინი. არსებობს კონსტიტუციური, გენეტიკურად არააქტიური ქრომატინი (სატელიტური დნმ), რომელიც შედგება არაგამოხატული უბნებისგან და ფაკულტატური - არააქტიური რიგ თაობებში, მაგრამ გარკვეულ გარემოებებში შეუძლია გამოხატოს.
აქტიური ქრომატინი (ევქრომატინი) არაკონდენსირებულია, ე.ი. ნაკლებად მჭიდროდ შეფუთული. სხვადასხვა უჯრედებში მისი შემცველობა 2-დან 11%-მდე მერყეობს. თავის ტვინის უჯრედებში ყველაზე მეტია - 10-11%, ღვიძლის უჯრედებში - 3-4 და თირკმელების - 2-3%. არსებობს ევქრომატინის აქტიური ტრანსკრიფცია. ამავდროულად, მისი სტრუქტურული ორგანიზაცია შესაძლებელს ხდის მოცემული ტიპის ორგანიზმისთვის დამახასიათებელი იგივე დნმ-ის გენეტიკური ინფორმაციის გამოყენებას სპეციალიზებულ უჯრედებში სხვადასხვა გზით.
ელექტრონულ მიკროსკოპში ქრომატინის გამოსახულება წააგავს მძივებს: სფერული გასქელება დაახლოებით 10 ნმ ზომით, გამოყოფილი ძაფისებრი ხიდებით. ამ სფერულ გასქელებას ნუკლეოსომებს უწოდებენ. ნუკლეოსომა არის ქრომატინის სტრუქტურული ერთეული. თითოეული ნუკლეოსომა შეიცავს 146 bp სიგრძის ზეგადახვეულ დნმ-ის სეგმენტს, რომელიც ქმნის 1,75 მარცხნივ ბრუნს თითო ნუკლეოსომის ბირთვზე. ნუკლეოსომური ბირთვი არის ჰისტონური ოქტამერი, რომელიც შედგება ჰისტონებისგან H2A, H2B, H3 და H4, თითოეული ტიპის ორი მოლეკულისგან (ნახ. 9), რომელიც ჰგავს დისკს, რომლის დიამეტრი 11 ნმ და სისქეა 5,7 ნმ. მეხუთე ჰისტონი, H1, არ არის ნუკლეოსომური ბირთვის ნაწილი და არ არის ჩართული ჰისტონის ოქტამერის გარშემო დნმ-ის დახვევის პროცესში. ის დაუკავშირდება დნმ-ს იმ წერტილებში, სადაც ორმაგი სპირალი შედის და გამოდის ნუკლეოსომური ბირთვიდან. ეს არის დნმ-ის შუალედური (მაკავშირებელი) სექციები, რომელთა სიგრძე უჯრედის ტიპის მიხედვით იცვლება 40-დან 50 ნუკლეოტიდურ წყვილამდე. შედეგად, დნმ-ის ფრაგმენტის სიგრძეც, რომელიც არის ნუკლეოსომების ნაწილი, ასევე იცვლება (186-დან 196 ნუკლეოტიდის წყვილამდე).
ნუკლეოსომა შეიცავს დნმ-ის დაახლოებით 90%-ს, დანარჩენი ნაწილი არის დამაკავშირებელი. ითვლება, რომ ნუკლეოსომა არის "ჩუმი" ქრომატინის ფრაგმენტები, ხოლო დამაკავშირებელი აქტიურია. თუმცა, ნუკლეოსომები შეიძლება განვითარდეს და გახდეს ხაზოვანი. გაშლილი ნუკლეოსომა უკვე აქტიური ქრომატინია. ეს ნათლად აჩვენებს ფუნქციის დამოკიდებულებას სტრუქტურაზე. შეიძლება ვივარაუდოთ, რომ რაც უფრო მეტია ქრომატინი გლობულური ნუკლეოსომების შემადგენლობაში, მით უფრო ნაკლებად აქტიურია იგი. ცხადია, სხვადასხვა უჯრედებში მოსვენებული ქრომატინის არათანაბარი პროპორცია დაკავშირებულია ასეთი ნუკლეოსომების რაოდენობასთან.
ელექტრონულ მიკროსკოპულ ფოტოებზე, იზოლაციის პირობებიდან და გაჭიმვის ხარისხიდან გამომდინარე, ქრომატინი შეიძლება გამოიყურებოდეს არა მხოლოდ როგორც გრძელი ძაფით გასქელება - ნუკლეოსომების "მძივები", არამედ როგორც უფრო მოკლე და მკვრივი ფიბრილა (ბოჭკო) დიამეტრით. 30 ნმ, რომლის ფორმირება შეინიშნება ჰისტონ H1 ურთიერთქმედების დროს, რომელიც დაკავშირებულია დნმ-ის და ჰისტონ H3-ის დამაკავშირებელ რეგიონთან, რაც იწვევს ექვსი ნუკლეოსომისგან შემდგარი სპირალის დამატებით გადახვევას 30 ნმ დიამეტრის სოლენოიდის წარმოქმნით. . ამ შემთხვევაში ჰისტონის პროტეინს შეუძლია ხელი შეუშალოს რიგი გენების ტრანსკრიფციას და ამით დაარეგულიროს მათი აქტივობა.
ზემოთ აღწერილი დნმ-ის ჰისტონებთან ურთიერთქმედების შედეგად, დნმ-ის ორმაგი სპირალის 186 წყვილის სეგმენტი საშუალო დიამეტრით 2 ნმ და სიგრძით 57 ნმ იქცევა სპირალში 10 ნმ დიამეტრით და სიგრძით. 5 ნმ. ამ სპირალის შემდგომი შეკუმშვით ბოჭკოზე 30 ნმ დიამეტრით, კონდენსაციის ხარისხი კიდევ ექვსჯერ იზრდება.
საბოლოო ჯამში, დნმ-ის დუპლექსის ხუთი ჰისტონებით შეფუთვა იწვევს დნმ-ის 50-ჯერ კონდენსაციას. თუმცა, კონდენსაციის ასეთი მაღალი ხარისხიც კი ვერ ხსნის დნმ-ის თითქმის 50,000-100,000-ჯერ შეკუმშვას მეტაფაზას ქრომოსომაში. სამწუხაროდ, ქრომატინის შემდგომი შეფუთვის დეტალები მეტაფაზა ქრომოსომამდე ჯერ არ არის ცნობილი, ამიტომ ამ პროცესის მხოლოდ ზოგადი მახასიათებლების გათვალისწინება შეიძლება.
დნმ-ის დატკეპნის დონე ქრომოსომებში
თითოეული დნმ-ის მოლეკულა შეფუთულია ცალკეულ ქრომოსომაში. ადამიანის დიპლოიდური უჯრედები შეიცავს 46 ქრომოსომას, რომლებიც განლაგებულია უჯრედის ბირთვში. უჯრედის ყველა ქრომოსომის დნმ-ის მთლიანი სიგრძე 1,74 მ-ია, მაგრამ ბირთვის დიამეტრი, რომელშიც ქრომოსომაა შეფუთული, მილიონჯერ მცირეა. დნმ-ის ასეთ კომპაქტურ შეფუთვას ქრომოსომებსა და ქრომოსომებში უჯრედის ბირთვში უზრუნველყოფს სხვადასხვა ჰისტონური და არაჰისტონის ცილები, რომლებიც ურთიერთქმედებენ დნმ-თან გარკვეული თანმიმდევრობით (იხ. ზემოთ). დნმ-ის დატკეპნა ქრომოსომებში შესაძლებელს ხდის მისი წრფივი ზომების შემცირებას დაახლოებით 10000-ჯერ - პირობითად 5 სმ-დან 5 მიკრონამდე. კომპაქტიზაციის რამდენიმე დონე არსებობს (სურ. 10).


- დნმ-ის ორმაგი სპირალი არის უარყოფითად დამუხტული მოლეკულა 2 ნმ დიამეტრით და რამდენიმე სმ სიგრძით.
- ნუკლეოსომური დონე- ქრომატინი გამოიყურება ელექტრონულ მიკროსკოპში, როგორც "მძივების" - ნუკლეოსომების ჯაჭვი - "ძაფზე". ნუკლეოსომა არის უნივერსალური სტრუქტურული ერთეული, რომელიც გვხვდება როგორც ევქრომატინში, ასევე ჰეტეროქრომატინში, ინტერფაზურ ბირთვში და მეტაფაზურ ქრომოსომებში.
დატკეპნის ნუკლეოსომურ დონეს უზრუნველყოფს სპეციალური ცილები - ჰისტონები. რვა დადებითად დამუხტული ჰისტონის დომენი ქმნის ნუკლეოსომის ბირთვს (ბირთს), რომლის ირგვლივ ჭრია უარყოფითად დამუხტული დნმ-ის მოლეკულა. ეს იძლევა შემცირებას 7-ჯერ, ხოლო დიამეტრი იზრდება 2-დან 11 ნმ-მდე.
- სოლენოიდის დონე
ქრომოსომის ორგანიზაციის სოლენოიდის დონეს ახასიათებს ნუკლეოსომური ძაფის გრეხილი და მისგან 20-35 ნმ დიამეტრის სქელი ფიბრილების - სოლენოიდების ან სუპერბიდების წარმოქმნით. სოლენოიდის სიმაღლეა 11 ნმ, და არის დაახლოებით 6-10 ნუკლეოსომა თითო შემობრუნებაზე. სოლენოიდის შეფუთვა უფრო სავარაუდოა, ვიდრე სუპერბიდ შეფუთვა, რომლის მიხედვითაც ქრომატინის ფიბრილი დიამეტრით 20-35 ნმ არის გრანულების ან სუპერბიდების ჯაჭვი, რომელთაგან თითოეული შედგება რვა ნუკლეოსომისგან. სოლენოიდის დონეზე დნმ-ის ხაზოვანი ზომა მცირდება 6-10-ჯერ, დიამეტრი იზრდება 30 ნმ-მდე.
- მარყუჟის დონე
მარყუჟის დონე უზრუნველყოფილია არაჰისტონის უბნის სპეციფიური დნმ-ის დამაკავშირებელი პროტეინებით, რომლებიც ამოიცნობენ და უკავშირდებიან დნმ-ის სპეციფიკურ თანმიმდევრობებს, ქმნიან მარყუჟებს დაახლოებით 30-300 კბ. მარყუჟი უზრუნველყოფს გენის ექსპრესიას, ე.ი. მარყუჟი არა მხოლოდ სტრუქტურული, არამედ ფუნქციური წარმონაქმნია. ამ დონეზე დაკლება ხდება 20-30-ჯერ. დიამეტრი იზრდება 300 ნმ-მდე. ამფიბიების კვერცხუჯრედებში მარყუჟის მსგავსი „სალამურის“ სტრუქტურები ჩანს ციტოლოგიურ პრეპარატებზე. ეს მარყუჟები, როგორც ჩანს, ზეგადახვეულია და წარმოადგენს დნმ-ის დომენებს, რომლებიც, სავარაუდოდ, შეესაბამება ქრომატინის ტრანსკრიფციისა და რეპლიკაციის ერთეულებს. სპეციფიკური პროტეინები აფიქსირებს მარყუჟების ფუძეებს და, შესაძლოა, მათ ზოგიერთ შიდა რეგიონს. მარყუჟის მსგავსი დომენური ორგანიზაცია ხელს უწყობს ქრომატინის დაკეცვას მეტაფაზურ ქრომოსომებში უფრო მაღალი რიგის სპირალურ სტრუქტურებად.
- დომენის დონე
ქრომოსომის ორგანიზაციის დომენის დონე საკმარისად არ არის შესწავლილი. ამ დონეზე აღინიშნება მარყუჟის დომენების ფორმირება - 25-30 ნმ სისქის ძაფების (ფიბრილების) სტრუქტურები, რომლებიც შეიცავს 60% პროტეინს, 35% დნმ-ს და 5% რნმ-ს, პრაქტიკულად უხილავია უჯრედის ციკლის ყველა ფაზაში. მიტოზის გამონაკლისი და გარკვეულწილად შემთხვევით ნაწილდება უჯრედის ბირთვზე. ამფიბიების კვერცხუჯრედებში მარყუჟის მსგავსი „სალამურის“ სტრუქტურები ჩანს ციტოლოგიურ პრეპარატებზე.
მარყუჟის დომენები მიმაგრებულია მათი ფუძით ინტრაბირთვულ ცილის მატრიქსზე ეგრეთ წოდებული ჩაშენებული მიმაგრების ადგილებში, ხშირად მოხსენიებული, როგორც MAR / SAR თანმიმდევრობები (MAR, ინგლისური მატრიცის ასოცირებული რეგიონიდან; SAR, ინგლისური ხარაჩოების მიმაგრების რეგიონებიდან) - დნმ-ის ფრაგმენტები რამდენიმე ასეული გრძელი ბაზის წყვილით, რომლებიც ხასიათდება A/T ბაზის წყვილების მაღალი შემცველობით (>65%). როგორც ჩანს, თითოეულ დომენს აქვს რეპლიკაციის ერთი საწყისი და ფუნქციონირებს, როგორც ავტონომიური ზეგადახვეული ერთეული. ნებისმიერი მარყუჟის დომენი შეიცავს ბევრ ტრანსკრიფციის ერთეულს, რომელთა ფუნქციონირება სავარაუდოდ კოორდინირებულია - მთელი დომენი არის ან აქტიურ ან არააქტიურ მდგომარეობაში.
დომენის დონეზე, ქრომატინის თანმიმდევრული შეფუთვის შედეგად, დნმ-ის ხაზოვანი ზომები მცირდება დაახლოებით 200-ჯერ (700 ნმ).
- ქრომოსომის დონე
ქრომოსომულ დონეზე პროფაზური ქრომოსომა კონდენსირდება მეტაფაზაში მარყუჟის დომენების დატკეპნით არაჰისტონის ცილების ღერძული ჩარჩოს გარშემო. ამ ზეგადახვევას თან ახლავს უჯრედის ყველა H1 მოლეკულის ფოსფორილირება. შედეგად, მეტაფაზის ქრომოსომა შეიძლება იყოს გამოსახული, როგორც მჭიდროდ შეფუთული სოლენოიდური მარყუჟები, რომლებიც დახვეულია მჭიდრო სპირალში. ტიპიური ადამიანის ქრომოსომა შეიძლება შეიცავდეს 2600-მდე მარყუჟს. ასეთი სტრუქტურის სისქე აღწევს 1400 ნმ-ს (ორი ქრომატიდს), ხოლო დნმ-ის მოლეკულა 104-ჯერ მცირდება, ე.ი. 5 სმ-დან გადაჭიმული დნმ 5 მკმ-მდე.
ქრომოსომების ფუნქციები
ექსტრაქრომოსომულ მექანიზმებთან ურთიერთქმედებისას ქრომოსომები უზრუნველყოფენ
- მემკვიდრეობითი ინფორმაციის შენახვა
- ამ ინფორმაციის გამოყენებით ფიჭური ორგანიზაციის შესაქმნელად და შესანარჩუნებლად
- მემკვიდრეობითი ინფორმაციის წაკითხვის რეგულირება
- გენეტიკური მასალის თვითგამრავლება
- გენეტიკური მასალის გადატანა დედის უჯრედიდან ქალიშვილურ უჯრედებზე.
არსებობს მტკიცებულება, რომ ქრომატინის რეგიონის გააქტიურებისას, ე.ი. ტრანსკრიფციის დროს მისგან ჯერ შექცევადად იხსნება ჰისტონი H1, შემდეგ კი ჰისტონის ოქტეტი. ეს იწვევს ქრომატინის დეკონდენსაციას, 30 ნმ ქრომატინის ფიბრილის თანმიმდევრულ გადასვლას 10 ნმ ძაფში და მის შემდგომ გაშლას დნმ-ის თავისუფალ რეგიონებში, ე.ი. ნუკლეოსომური სტრუქტურის დაკარგვა.
ყველამ ვიცით, რომ ადამიანის გარეგნობა, ზოგიერთი ჩვევა და დაავადებაც კი მემკვიდრეობით მიიღება. მთელი ეს ინფორმაცია ცოცხალი არსების შესახებ დაშიფრულია გენებში. მაშ, რას ჰგავს ეს ცნობილი გენები, როგორ ფუნქციონირებს ისინი და სად მდებარეობს?
ასე რომ, ნებისმიერი ადამიანის ან ცხოველის ყველა გენის მატარებელი არის დნმ. ეს ნაერთი აღმოაჩინა იოჰან ფრიდრიხ მიშერმა 1869 წელს.ქიმიურად დნმ არის დეზოქსირიბონუკლეინის მჟავა. Რას ნიშნავს ეს? როგორ ატარებს ეს მჟავა ჩვენს პლანეტაზე მთელი სიცოცხლის გენეტიკურ კოდს?
დავიწყოთ იმით, თუ სად მდებარეობს დნმ. ადამიანის უჯრედში ბევრი ორგანელაა, რომლებიც ასრულებენ სხვადასხვა ფუნქციებს. დნმ მდებარეობს ბირთვში. ბირთვი არის პატარა ორგანელა, რომელიც გარშემორტყმულია სპეციალური მემბრანით, რომელიც ინახავს მთელ გენეტიკურ მასალას - დნმ.
როგორია დნმ-ის მოლეკულის აგებულება?
ჯერ ვნახოთ, რა არის დნმ. დნმ არის ძალიან გრძელი მოლეკულა, რომელიც შედგება სტრუქტურული ელემენტებისაგან - ნუკლეოტიდებით. არსებობს 4 სახის ნუკლეოტიდი - ადენინი (A), თიმინი (T), გუანინი (G) და ციტოზინი (C). ნუკლეოტიდების ჯაჭვი სქემატურად ასე გამოიყურება: GGAATTSTAAG.... ნუკლეოტიდების ეს თანმიმდევრობა არის დნმ-ის ჯაჭვი.დნმ-ის სტრუქტურა პირველად 1953 წელს გაშიფრეს ჯეიმს უოტსონმა და ფრენსის კრიკმა.
დნმ-ის ერთ მოლეკულაში არის ნუკლეოტიდების ორი ჯაჭვი, რომლებიც სპირალურადაა გადაბმული ერთმანეთის გარშემო. როგორ ეკვრება ეს ნუკლეოტიდური ჯაჭვები ერთმანეთს და ტრიალდება სპირალში? ეს ფენომენი განპირობებულია კომპლემენტარობის თვისებით. კომპლემენტარულობა ნიშნავს, რომ მხოლოდ გარკვეული ნუკლეოტიდები (შემავსებელი) შეიძლება იყოს ერთმანეთის საპირისპიროდ ორ ჯაჭვში. ასე რომ, საპირისპირო ადენინი ყოველთვის არის თიმინი, ხოლო საპირისპირო გუანინი ყოველთვის მხოლოდ ციტოზინია. ამრიგად, გუანინი ავსებს ციტოზინს, ხოლო ადენინი თიმინს.ნუკლეოტიდების ასეთ წყვილებს ერთმანეთის მოპირდაპირე ჯაჭვებში კომპლემენტურსაც უწოდებენ.
 ის სქემატურად შეიძლება წარმოდგენილი იყოს შემდეგნაირად:
ის სქემატურად შეიძლება წარმოდგენილი იყოს შემდეგნაირად:
G - C
თ - ა
თ - ა
გ - გ
ეს დამატებითი წყვილი A - T და G - C ყალიბდება ქიმიური ბმაწყვილის ნუკლეოტიდებს შორის და G-სა და C-ს შორის კავშირი უფრო ძლიერია, ვიდრე A-სა და T-ს შორის. ბმა იქმნება მკაცრად კომპლემენტარულ ფუძეებს შორის, ანუ არაკომპლექტურ G-სა და A-ს შორის კავშირის ფორმირება შეუძლებელია.
დნმ-ის „შეფუთვა“, როგორ ხდება დნმ-ის ჯაჭვი ქრომოსომად?
რატომ ტრიალებს დნმ-ის ეს ნუკლეოტიდური ჯაჭვები ერთმანეთის გარშემო? რატომ არის ეს საჭირო? ფაქტია, რომ ნუკლეოტიდების რაოდენობა უზარმაზარია და ამხელა ჯაჭვების დასაყენებლად დიდი სივრცე გჭირდებათ. ამ მიზეზით, არსებობს დნმ-ის ორი ჯაჭვის სპირალური გადახვევა მეორის გარშემო. ეს ფენომენისპირალიზაციას უწოდებენ. სპირალიზაციის შედეგად დნმ-ის ჯაჭვები 5-6-ჯერ მცირდება.დნმ-ის ზოგიერთ მოლეკულას ორგანიზმი აქტიურად იყენებს, ზოგს კი იშვიათად. ასეთი იშვიათად გამოყენებული დნმ-ის მოლეკულები, გარდა ჰელიკალიზაციისა, ექვემდებარება კიდევ უფრო კომპაქტურ "შეფუთვას". ასეთ კომპაქტურ შეფუთვას სუპერკოილინგი ეწოდება და დნმ-ის ჯაჭვს 25-30-ჯერ ამოკლებს!
როგორ არის შეფუთული დნმ სპირალი?
 ზეგადახვევისთვის გამოიყენება ჰისტონური ცილები, რომლებსაც აქვთ ღეროს ან ძაფის კოჭის გარეგნობა და სტრუქტურა. სპირალიზებული დნმ-ის ძაფები იჭრება ამ „ხვეულებზე“ – ჰისტონურ პროტეინებზე. ამ გზით, გრძელი ძაფი ძალიან კომპაქტურად იკვრება და ძალიან მცირე ადგილს იკავებს.
ზეგადახვევისთვის გამოიყენება ჰისტონური ცილები, რომლებსაც აქვთ ღეროს ან ძაფის კოჭის გარეგნობა და სტრუქტურა. სპირალიზებული დნმ-ის ძაფები იჭრება ამ „ხვეულებზე“ – ჰისტონურ პროტეინებზე. ამ გზით, გრძელი ძაფი ძალიან კომპაქტურად იკვრება და ძალიან მცირე ადგილს იკავებს. თუ საჭიროა ამა თუ იმ დნმ-ის მოლეკულის გამოყენება, ხდება „გახსნის“ პროცესი, ანუ დნმ-ის ძაფი „იხსნება“ „კოჭიდან“ - ჰისტონის ცილა (თუ მასზე იყო დახვეული) და იხსნება. სპირალი ორ პარალელურ ჯაჭვად. და როცა დნმ-ის მოლეკულა ასეთ გაუფუჭებელ მდგომარეობაშია, მაშინ მისგან საჭირო გენეტიკური ინფორმაციის წაკითხვა შეიძლება. უფრო მეტიც, გენეტიკური ინფორმაციის წაკითხვა ხდება მხოლოდ დნმ-ის ძაფებიდან!
ზემოხვეული ქრომოსომების ერთობლიობას ე.წ ჰეტეროქრომატინიდა ინფორმაციის წასაკითხად ხელმისაწვდომი ქრომოსომები - ევქრომატინი.
რა არის გენები, როგორია მათი კავშირი დნმ-თან?
ახლა ვნახოთ რა არის გენები. ცნობილია, რომ არსებობს გენები, რომლებიც განსაზღვრავენ ჩვენი სხეულის სისხლის ჯგუფს, თვალების ფერს, თმას, კანს და სხვა ბევრ თვისებას. გენი არის დნმ-ის მკაცრად განსაზღვრული განყოფილება, რომელიც შედგება გარკვეული რაოდენობის ნუკლეოტიდებისგან, რომლებიც განლაგებულია მკაცრად განსაზღვრულ კომბინაციაში. დნმ-ის მკაცრად განსაზღვრულ მონაკვეთში მდებარეობა ნიშნავს, რომ კონკრეტულ გენს თავისი ადგილი აქვს და ამ ადგილის შეცვლა შეუძლებელია. მიზანშეწონილია ასეთი შედარება: ადამიანი ცხოვრობს გარკვეულ ქუჩაზე, გარკვეულ სახლში და ბინაში და ადამიანი თვითნებურად ვერ გადადის სხვა სახლში, ბინაში ან სხვა ქუჩაზე. ნუკლეოტიდების გარკვეული რაოდენობა გენში ნიშნავს, რომ თითოეულ გენს აქვს ნუკლეოტიდების სპეციფიკური რაოდენობა და არ შეიძლება გახდეს მეტი ან ნაკლები. მაგალითად, ინსულინის გამომუშავების კოდირების გენი 60 ბაზის წყვილია; ჰორმონის ოქსიტოცინის გამომუშავების კოდირების გენი არის 370 bp. მკაცრი ნუკლეოტიდური თანმიმდევრობა უნიკალურია თითოეული გენისთვის და მკაცრად განსაზღვრული. მაგალითად, AATTAATA თანმიმდევრობა არის გენის ფრაგმენტი, რომელიც კოდირებს ინსულინის წარმოებას. ინსულინის მისაღებად გამოიყენება სწორედ ასეთი თანმიმდევრობა; მაგალითად, ადრენალინის მისაღებად გამოიყენება ნუკლეოტიდების განსხვავებული კომბინაცია. მნიშვნელოვანია გვესმოდეს, რომ ნუკლეოტიდების მხოლოდ გარკვეული კომბინაცია აკოდირებს გარკვეულ „პროდუქტს“ (ადრენალინი, ინსულინი და ა.შ.). ნუკლეოტიდების გარკვეული რაოდენობის ასეთი უნიკალური კომბინაცია, რომელიც დგას "თავის ადგილზე" - ეს არის გენი.
გარდა გენებისა, დნმ-ის ჯაჭვში განლაგებულია ე.წ. ასეთი არაკოდირების ნუკლეოტიდური თანმიმდევრობები არეგულირებს გენების ფუნქციონირებას, ხელს უწყობს ქრომოსომის სპირალიზაციას და აღნიშნავს გენის საწყისი და დასასრული წერტილები. თუმცა, დღემდე, არაკოდირების უმეტესობის როლი გაურკვეველი რჩება.
რა არის ქრომოსომა? სქესის ქრომოსომები
ინდივიდის გენების მთლიანობას გენომი ეწოდება. ბუნებრივია, მთელი გენომი არ შეიძლება შეფუთული იყოს ერთ დნმ-ში. გენომი დაყოფილია დნმ-ის 46 წყვილ მოლეკულად. დნმ-ის მოლეკულების ერთ წყვილს ქრომოსომა ეწოდება. ასე რომ, ზუსტად ამ ქრომოსომებს აქვს ადამიანს 46 ცალი. თითოეული ქრომოსომა ატარებს გენების მკაცრად განსაზღვრულ კომპლექტს, მაგალითად, მე-18 ქრომოსომა შეიცავს თვალის ფერის მაკოდირებელ გენებს და ა.შ. ქრომოსომა ერთმანეთისგან განსხვავდება სიგრძით და ფორმით. ყველაზე გავრცელებული ფორმებია X ან Y სახით, მაგრამ არის სხვებიც. ადამიანს აქვს ორი ერთნაირი ფორმის ქრომოსომა, რომლებსაც დაწყვილებულს (წყვილებს) უწოდებენ. ასეთ განსხვავებებთან დაკავშირებით, ყველა დაწყვილებული ქრომოსომა დანომრილია - არის 23 წყვილი. ეს ნიშნავს, რომ არსებობს წყვილი ქრომოსომა #1, წყვილი #2, #3 და ა.შ. თითოეული გენი, რომელიც პასუხისმგებელია კონკრეტულ მახასიათებლებზე, მდებარეობს იმავე ქრომოსომაზე. სპეციალისტების თანამედროვე სახელმძღვანელოებში გენის ლოკალიზაცია შეიძლება მიეთითოს, მაგალითად, შემდეგნაირად: ქრომოსომა 22, გრძელი მკლავი.რა განსხვავებაა ქრომოსომებს შორის?
 კიდევ რით განსხვავდებიან ქრომოსომა ერთმანეთისგან? რას ნიშნავს ტერმინი გრძელი მკლავი? ავიღოთ X- ფორმის ქრომოსომა. დნმ-ის ჯაჭვების გადაკვეთა შეიძლება მოხდეს მკაცრად შუა (X), ან შეიძლება მოხდეს არა ცენტრალურში. როდესაც დნმ-ის ჯაჭვების ასეთი გადაკვეთა არ ხდება ცენტრალურად, მაშინ გადაკვეთის წერტილთან შედარებით, ზოგიერთი ბოლო უფრო გრძელია, სხვები, შესაბამისად, უფრო მოკლე. ასეთ გრძელ ბოლოებს ჩვეულებრივ უწოდებენ ქრომოსომის გრძელ მკლავს და მოკლე ბოლოებს, შესაბამისად, მოკლე მკლავს. Y-ის ფორმის ქრომოსომებს უმეტესად გრძელი მკლავები უკავია, მოკლე კი ძალიან მცირეა (სქემურ სურათზეც კი არ არის მითითებული).
კიდევ რით განსხვავდებიან ქრომოსომა ერთმანეთისგან? რას ნიშნავს ტერმინი გრძელი მკლავი? ავიღოთ X- ფორმის ქრომოსომა. დნმ-ის ჯაჭვების გადაკვეთა შეიძლება მოხდეს მკაცრად შუა (X), ან შეიძლება მოხდეს არა ცენტრალურში. როდესაც დნმ-ის ჯაჭვების ასეთი გადაკვეთა არ ხდება ცენტრალურად, მაშინ გადაკვეთის წერტილთან შედარებით, ზოგიერთი ბოლო უფრო გრძელია, სხვები, შესაბამისად, უფრო მოკლე. ასეთ გრძელ ბოლოებს ჩვეულებრივ უწოდებენ ქრომოსომის გრძელ მკლავს და მოკლე ბოლოებს, შესაბამისად, მოკლე მკლავს. Y-ის ფორმის ქრომოსომებს უმეტესად გრძელი მკლავები უკავია, მოკლე კი ძალიან მცირეა (სქემურ სურათზეც კი არ არის მითითებული). ქრომოსომების ზომა მერყეობს: ყველაზე დიდია No1 და No3 წყვილის ქრომოსომა, უმცირესი ქრომოსომა No17, No19 წყვილების.
ფორმისა და ზომის გარდა, ქრომოსომები განსხვავდებიან თავიანთი ფუნქციებით. 23 წყვილიდან 22 წყვილი სომატურია, 1 წყვილი სექსუალური. Რას ნიშნავს? სომატური ქრომოსომა განსაზღვრავს ინდივიდის ყველა გარეგნულ ნიშანს, მის თვისებებს ქცევითი რეაქციები, მემკვიდრეობითი ფსიქოტიპი, ანუ თითოეული ინდივიდუალური ადამიანის ყველა მახასიათებელი და მახასიათებელი. სქესის ქრომოსომათა წყვილი განსაზღვრავს ადამიანის სქესს: მამრობითი ან მდედრობითი სქესის. ადამიანის სქესის ქრომოსომა ორი ტიპისაა – X (X) და Y (Y). თუ ისინი შერწყმულია როგორც XX (x - x) - ეს არის ქალი, ხოლო თუ XY (x - y) - ჩვენ წინ გვყავს მამაკაცი.
მემკვიდრეობითი დაავადებები და ქრომოსომის დაზიანება
თუმცა ხდება გენომის „დაშლა“, შემდეგ ადამიანებში ვლინდება გენეტიკური დაავადებები. მაგალითად, როდესაც 21 წყვილ ქრომოსომაში სამი ქრომოსომაა ორის ნაცვლად, ადამიანი დაუნის სინდრომით იბადება.არსებობს გენეტიკური მასალის მრავალი უფრო მცირე „დაშლა“, რომელიც არ იწვევს დაავადების დაწყებას, პირიქით, კარგ თვისებებს აძლევს. გენეტიკური მასალის ყველა „დაშლას“ მუტაციას უწოდებენ. ნეგატიურად განიხილება მუტაციები, რომლებიც იწვევს დაავადებას ან ორგანიზმის თვისებების გაუარესებას, ხოლო მუტაციები, რომლებიც იწვევს ახალი სასარგებლო თვისებების წარმოქმნას, დადებითად.
თუმცა, იმ დაავადებების უმეტესობასთან დაკავშირებით, რომლებსაც დღეს ადამიანები აწუხებთ, ეს არ არის მემკვიდრეობითი დაავადება, არამედ მხოლოდ მიდრეკილება. მაგალითად, ბავშვის მამაში შაქარი ნელა შეიწოვება. ეს არ ნიშნავს იმას, რომ ბავშვი დიაბეტით დაიბადება, მაგრამ ბავშვს ექნება მიდრეკილება. ეს ნიშნავს, რომ თუ ბავშვი ბოროტად მოიხმარს ტკბილეულს და ფქვილის პროდუქტებს, მაშინ მას განუვითარდება დიაბეტი.
დღეს ე.წ პრედიკატივიმედიცინა. როგორც ამ სამედიცინო პრაქტიკის ნაწილი, ადამიანში ვლინდება მიდრეკილებები (შესაბამისი გენების იდენტიფიკაციის საფუძველზე), შემდეგ კი მას ეძლევა რეკომენდაციები - რა დიეტა უნდა დაიცვას, როგორ სწორად შეცვალოს სამუშაო და დასვენების რეჟიმები, რომ არ მიიღოს ავადმყოფი.
როგორ წავიკითხოთ დნმ-ში დაშიფრული ინფორმაცია?
მაგრამ როგორ შეგიძლიათ წაიკითხოთ დნმ-ში მოცემული ინფორმაცია? როგორ იყენებს მას საკუთარი სხეული? თავად დნმ არის ერთგვარი მატრიცა, მაგრამ არა მარტივი, არამედ კოდირებული. დნმ-ის მატრიციდან ინფორმაციის წასაკითხად ის ჯერ სპეციალურ მატარებელზე - რნმ-ზე გადადის. რნმ არის ქიმიურად რიბონუკლეინის მჟავა. ის განსხვავდება დნმ-ისგან იმით, რომ მას შეუძლია ბირთვული მემბრანის მეშვეობით უჯრედში გადასვლა და დნმ-ს ეს უნარი აკლია (ის შეიძლება იყოს მხოლოდ ბირთვში). დაშიფრული ინფორმაცია გამოიყენება თავად უჯრედში. ასე რომ, რნმ არის კოდირებული ინფორმაციის გადამტანი ბირთვიდან უჯრედამდე.როგორ ხდება რნმ-ის სინთეზი, როგორ სინთეზირდება ცილა რნმ-ის დახმარებით?
 დნმ-ის ჯაჭვები, რომლებიდანაც ინფორმაცია უნდა „წაკითხულიყო“, იხსნება, სპეციალური ფერმენტი, „მშენებელი“ უახლოვდება მათ და ასინთეზებს დამატებით რნმ-ს ჯაჭვს დნმ-ის ჯაჭვის პარალელურად. რნმ-ის მოლეკულა ასევე შედგება 4 ტიპის ნუკლეოტიდებისგან - ადენინი (A), ურაცილი (U), გუანინი (G) და ციტოზინი (C). ამ შემთხვევაში, შემდეგი წყვილები ერთმანეთს ავსებენ: ადენინი - ურაცილი, გუანინი - ციტოზინი. როგორც ხედავთ, დნმ-ისგან განსხვავებით, რნმ თიმინის ნაცვლად იყენებს ურაცილს. ანუ „მშენებელი“ ფერმენტი მუშაობს შემდეგნაირად: თუ დნმ-ის ჯაჭვში ხედავს A-ს, მაშინ რნმ-ის ჯაჭვს უერთდება Y, თუ G-ს, მაშინ უერთდება C და ა.შ. ამრიგად, ტრანსკრიფციის დროს თითოეული აქტიური გენიდან იქმნება შაბლონი - რნმ-ის ასლი, რომელსაც შეუძლია გაიაროს ბირთვული მემბრანა.
დნმ-ის ჯაჭვები, რომლებიდანაც ინფორმაცია უნდა „წაკითხულიყო“, იხსნება, სპეციალური ფერმენტი, „მშენებელი“ უახლოვდება მათ და ასინთეზებს დამატებით რნმ-ს ჯაჭვს დნმ-ის ჯაჭვის პარალელურად. რნმ-ის მოლეკულა ასევე შედგება 4 ტიპის ნუკლეოტიდებისგან - ადენინი (A), ურაცილი (U), გუანინი (G) და ციტოზინი (C). ამ შემთხვევაში, შემდეგი წყვილები ერთმანეთს ავსებენ: ადენინი - ურაცილი, გუანინი - ციტოზინი. როგორც ხედავთ, დნმ-ისგან განსხვავებით, რნმ თიმინის ნაცვლად იყენებს ურაცილს. ანუ „მშენებელი“ ფერმენტი მუშაობს შემდეგნაირად: თუ დნმ-ის ჯაჭვში ხედავს A-ს, მაშინ რნმ-ის ჯაჭვს უერთდება Y, თუ G-ს, მაშინ უერთდება C და ა.შ. ამრიგად, ტრანსკრიფციის დროს თითოეული აქტიური გენიდან იქმნება შაბლონი - რნმ-ის ასლი, რომელსაც შეუძლია გაიაროს ბირთვული მემბრანა. როგორ ხდება ცილის სინთეზი კოდირებული კონკრეტული გენით?
ბირთვიდან გამოსვლის შემდეგ, რნმ შედის ციტოპლაზმაში. უკვე ციტოპლაზმაში, რნმ შეიძლება იყოს, როგორც მატრიქსი, ჩაშენებული სპეციალურ ფერმენტულ სისტემებში (რიბოსომები), რომლებსაც შეუძლიათ სინთეზირება, რნმ-ის ინფორმაციით, ცილის შესაბამისი ამინომჟავების თანმიმდევრობის სინთეზირება. მოგეხსენებათ, ცილის მოლეკულა შედგება ამინომჟავებისგან. როგორ ახერხებს რიბოსომა გაიგოს, რომელი ამინომჟავა მიამაგროს მზარდ ცილოვან ჯაჭვს? ეს კეთდება სამმაგი კოდის საფუძველზე. სამმაგი კოდი ნიშნავს, რომ რნმ-ის ჯაჭვის სამი ნუკლეოტიდის თანმიმდევრობა ( სამეული,მაგალითად, GGU) კოდი ერთი ამინომჟავისთვის (ამ შემთხვევაში, გლიცინი). თითოეული ამინომჟავა კოდირებულია კონკრეტული სამეულით. ასე რომ, რიბოსომა „კითხულობს“ სამეულს, ადგენს, რომელი ამინომჟავა უნდა დაემატოს რნმ-ში ინფორმაციის წაკითხვისას. როდესაც ამინომჟავების ჯაჭვი იქმნება, ის იღებს გარკვეულ სივრცულ ფორმას და ხდება ცილა, რომელსაც შეუძლია შეასრულოს მისთვის დაკისრებული ფერმენტული, სამშენებლო, ჰორმონალური და სხვა ფუნქციები.ნებისმიერი ცოცხალი ორგანიზმისთვის ცილა არის გენის პროდუქტი. ეს არის ცილები, რომლებიც განსაზღვრავენ გენების ყველა სხვადასხვა თვისებას, თვისებასა და გარეგნულ გამოვლინებას.
მარჯვნივ არის ყველაზე დიდი ადამიანის დნმ-ის სპირალი, რომელიც აშენებულია ადამიანებისგან ვარნას სანაპიროზე (ბულგარეთი), რომელიც გინესის რეკორდების წიგნში შევიდა 2016 წლის 23 აპრილს.
Დეზოქსირიბონუკლეინის მჟავა. Ზოგადი ინფორმაცია
დნმ (დეზოქსირიბონუკლეინის მჟავა) არის სიცოცხლის ერთგვარი გეგმა, რთული კოდი, რომელიც შეიცავს მონაცემებს მემკვიდრეობითი ინფორმაციის შესახებ. ამ რთულ მაკრომოლეკულას შეუძლია შეინახოს და გადასცეს მემკვიდრეობითი გენეტიკური ინფორმაცია თაობიდან თაობას. დნმ განსაზღვრავს ნებისმიერი ცოცხალი ორგანიზმის ისეთ თვისებებს, როგორიცაა მემკვიდრეობა და ცვალებადობა. მასში კოდირებული ინფორმაცია განსაზღვრავს ნებისმიერი ცოცხალი ორგანიზმის განვითარების მთელ პროგრამას. გენეტიკურად ჩადებული ფაქტორები წინასწარ განსაზღვრავს როგორც ადამიანის, ისე ნებისმიერი სხვა ორგანიზმის ცხოვრების მთელ კურსს. გარე გარემოს ხელოვნურმა ან ბუნებრივმა ზემოქმედებამ შეიძლება მხოლოდ ოდნავ იმოქმედოს ცალკეული გენეტიკური თვისებების საერთო სიმძიმეზე ან დაპროგრამებული პროცესების განვითარებაზე.
Დეზოქსირიბონუკლეინის მჟავა(დნმ) არის მაკრომოლეკულა (სამიდან ერთ-ერთი მთავარი, დანარჩენი ორი არის რნმ და ცილა), რომელიც უზრუნველყოფს შენახვას, გადაცემას თაობიდან თაობას და ცოცხალი ორგანიზმების განვითარებისა და ფუნქციონირების გენეტიკური პროგრამის განხორციელებას. დნმ შეიცავს ინფორმაციას სხვადასხვა ტიპის რნმ-ისა და ცილების სტრუქტურის შესახებ.
ევკარიოტულ უჯრედებში (ცხოველები, მცენარეები და სოკოები) დნმ გვხვდება უჯრედის ბირთვში, როგორც ქრომოსომების ნაწილი, ასევე ზოგიერთ უჯრედულ ორგანელებში (მიტოქონდრია და პლასტიდები). პროკარიოტული ორგანიზმების უჯრედებში (ბაქტერიები და არქეები) შიგნიდან უჯრედის მემბრანაზე მიმაგრებულია დნმ-ის წრიული ან ხაზოვანი მოლეკულა, ეგრეთ წოდებული ნუკლეოიდი. მათ და ქვედა ევკარიოტებს (მაგალითად, საფუარს) ასევე აქვთ მცირე ავტონომიური, ძირითადად წრიული დნმ-ის მოლეკულები, რომლებსაც პლაზმიდები ეწოდება.
ქიმიური თვალსაზრისით, დნმ არის გრძელი პოლიმერული მოლეკულა, რომელიც შედგება განმეორებადი ბლოკებისგან - ნუკლეოტიდებისგან. თითოეული ნუკლეოტიდი შედგება აზოტოვანი ფუძისგან, შაქრისგან (დეოქსირიბოზა) და ფოსფატის ჯგუფისგან. ჯაჭვში ნუკლეოტიდებს შორის კავშირები წარმოიქმნება დეზოქსირიბოზას მიერ ( FROM) და ფოსფატი ( ფ) ჯგუფები (ფოსფოდიესტერული ბმები).

ბრინჯი. 2. ნუკლეტიდი შედგება აზოტოვანი ფუძისგან, შაქრისგან (დეზოქსირიბოზა) და ფოსფატის ჯგუფისგან.
შემთხვევების აბსოლუტურ უმრავლესობაში (გარდა ზოგიერთი ვირუსისა, რომელიც შეიცავს ერთჯაჭვიან დნმ-ს), დნმ მაკრომოლეკულა შედგება ორი ჯაჭვისგან, რომლებიც ორიენტირებულია აზოტოვანი ბაზებით ერთმანეთზე. ეს ორჯაჭვიანი მოლეკულა ხვეულია სპირალში.
დნმ-ში ნაპოვნია აზოტოვანი ფუძის ოთხი ტიპი (ადენინი, გუანინი, თიმინი და ციტოზინი). ერთ-ერთი ჯაჭვის აზოტოვანი ფუძეები დაკავშირებულია მეორე ჯაჭვის აზოტოვან ფუძებთან წყალბადური ბმებით კომპლემენტარობის პრინციპის მიხედვით: ადენინი ერწყმის მხოლოდ თიმინს ( A-T), გუანინი - მხოლოდ ციტოზინით ( G-C). სწორედ ეს წყვილები ქმნიან დნმ-ის ხვეული „კიბის“ „საფეხურებს“ (იხ.: სურ. 2, 3 და 4).

ბრინჯი. 2. აზოტოვანი ფუძეები
ნუკლეოტიდების თანმიმდევრობა საშუალებას გაძლევთ „დაშიფვროთ“ ინფორმაცია სხვადასხვა სახისრნმ, რომელთაგან ყველაზე მნიშვნელოვანია ინფორმაცია ან შაბლონი (mRNA), რიბოსომული (rRNA) და ტრანსპორტი (tRNA). ყველა ამ ტიპის რნმ სინთეზირდება დნმ-ის შაბლონზე დნმ-ის თანმიმდევრობის კოპირებით ტრანსკრიპციის დროს სინთეზირებულ რნმ-ის თანმიმდევრობაში და მონაწილეობს ცილების ბიოსინთეზში (თარგმანის პროცესი). გარდა კოდირების თანმიმდევრობისა, უჯრედის დნმ შეიცავს თანმიმდევრობებს, რომლებიც ასრულებენ მარეგულირებელ და სტრუქტურულ ფუნქციებს.

ბრინჯი. 3. დნმ-ის რეპლიკაცია
დნმ-ის ქიმიური ნაერთების ძირითადი კომბინაციების მდებარეობა და ამ კომბინაციებს შორის რაოდენობრივი თანაფარდობა უზრუნველყოფს მემკვიდრეობითი ინფორმაციის დაშიფვრას.
Განათლება ახალი დნმ (რეპლიკაცია)
- რეპლიკაციის პროცესი: დნმ-ის ორმაგი სპირალის გახსნა - დამატებითი ჯაჭვების სინთეზი დნმ პოლიმერაზას მიერ - დნმ-ის ორი მოლეკულის წარმოქმნა ერთიდან.
- ორმაგი სპირალი "იხსნება" ორ ტოტად, როდესაც ფერმენტები არღვევს კავშირს ქიმიური ნაერთების ფუძე წყვილებს შორის.
- თითოეული ტოტი არის ახალი დნმ-ის ელემენტი. ახალი ბაზის წყვილები დაკავშირებულია იმავე თანმიმდევრობით, როგორც მშობლის ფილიალში.
დუბლირების დასრულების შემდეგ წარმოიქმნება ორი დამოუკიდებელი სპირალი, რომლებიც შექმნილია მშობელი დნმ-ის ქიმიური ნაერთებისგან და აქვთ იგივე გენეტიკური კოდი. ამ გზით, დნმ-ს შეუძლია ინფორმაციის ამოღება უჯრედიდან უჯრედში.
უფრო დეტალური ინფორმაცია:
ნუკლეინის მჟავების სტრუქტურა

ბრინჯი. 4 . აზოტოვანი ფუძეები: ადენინი, გუანინი, ციტოზინი, თიმინი
Დეზოქსირიბონუკლეინის მჟავა(დნმ) ეხება ნუკლეინის მჟავებს. Ნუკლეინის მჟავაარის არარეგულარული ბიოპოლიმერების კლასი, რომელთა მონომერები არის ნუკლეოტიდები.
ნუკლეოტიდებიშედგება აზოტოვანი ბაზადაკავშირებულია ხუთნახშირბადიან ნახშირწყალთან (პენტოზა) - დეზოქსირიბოზა(დნმ-ის შემთხვევაში) ან რიბოზა(რნმ-ის შემთხვევაში), რომელიც ერწყმის ფოსფორმჟავას ნარჩენს (H 2 PO 3 -).
აზოტოვანი ფუძეებიარსებობს ორი ტიპი: პირიმიდინის ფუძეები - ურაცილი (მხოლოდ რნმ-ში), ციტოზინი და თიმინი, პურინის ფუძეები - ადენინი და გუანინი.

ბრინჯი. სურ. 5. ნუკლეოტიდების სტრუქტურა (მარცხნივ), ნუკლეოტიდის მდებარეობა დნმ-ში (ქვედა) და აზოტოვანი ფუძეების ტიპები (მარჯვნივ): პირიმიდინი და პურინი

ნახშირბადის ატომები პენტოზას მოლეკულაში დანომრილია 1-დან 5-მდე. ფოსფატი ერწყმის მესამე და მეხუთე ნახშირბადის ატომებს. ასე უკავშირდებიან ნუკლეინის მჟავები და ქმნიან ნუკლეინის მჟავების ჯაჭვს. ამრიგად, ჩვენ შეგვიძლია გამოვყოთ დნმ-ის ჯაჭვის 3' და 5' ბოლოები:

ბრინჯი. 6. დნმ-ის ჯაჭვის 3' და 5' ბოლოების იზოლაცია
დნმ-ის ორი ჯაჭვი იქმნება ორმაგი სპირალი. ეს ჯაჭვები სპირალურად არის ორიენტირებული საპირისპირო მიმართულებით. დნმ-ის სხვადასხვა ჯაჭვებში აზოტოვანი ფუძეები ერთმანეთთან არის დაკავშირებული წყალბადის ბმები. ადენინი ყოველთვის ერწყმის თიმინს, ხოლო ციტოზინი ყოველთვის აერთიანებს გუანინს. მას ეძახიან კომპლემენტარობის წესი(სმ. კომპლემენტარობის პრინციპი).
კომპლემენტარობის წესი:
| A-T G-C |
მაგალითად, თუ გვეძლევა დნმ-ის ჯაჭვი, რომელსაც აქვს თანმიმდევრობა
3'-ATGTCCTAGCTGCTCG - 5',
მაშინ მეორე ჯაჭვი იქნება მისი შემავსებელი და მიმართული საპირისპირო მიმართულებით - 5'-ბოლოდან 3'-ბოლომდე:
5'- TACAGGATCGACGAGC- 3'.

ბრინჯი. 7. დნმ-ის მოლეკულის ჯაჭვების მიმართულება და აზოტოვანი ფუძეების შეერთება წყალბადური ბმების გამოყენებით.
დნმ-ის რეპლიკაცია
დნმ-ის რეპლიკაციაარის დნმ-ის მოლეკულის გაორმაგების პროცესი შაბლონის სინთეზით. დნმ-ის ბუნებრივი რეპლიკაციის უმეტეს შემთხვევაშიპრაიმერიდნმ-ის სინთეზისთვის არის მოკლე ნაწყვეტი (ხელახლა შეიქმნა). ასეთი რიბონუკლეოტიდური პრაიმერი იქმნება ფერმენტ პრიმაზას მიერ (დნმ პრიმაზა პროკარიოტებში, დნმ პოლიმერაზა ევკარიოტებში) და შემდგომში ჩანაცვლებულია დეზოქსირიბონუკლეოტიდის პოლიმერაზათი, რომელიც ჩვეულებრივ ასრულებს აღდგენის ფუნქციებს (ქიმიური დაზიანების გამოსწორება და დნმ-ის მოლეკულაში რღვევები).
რეპლიკაცია ხდება ნახევრად კონსერვატიული გზით. ეს ნიშნავს, რომ დნმ-ის ორმაგი სპირალი იხსნება და მის თითოეულ ჯაჭვზე სრულდება ახალი ჯაჭვი კომპლემენტარობის პრინციპის მიხედვით. ამრიგად, ქალიშვილი დნმ-ის მოლეკულა შეიცავს ერთ ჯაჭვს მშობლის მოლეკულიდან და ერთ ახლად სინთეზირებულს. რეპლიკაცია ხდება მშობელი ჯაჭვის 3'-დან 5'-მდე მიმართულებით.

ბრინჯი. 8. დნმ-ის მოლეკულის რეპლიკაცია (გაორმაგება).
დნმ სინთეზი- ეს არც ისე რთული პროცესია, როგორც ერთი შეხედვით შეიძლება ჩანდეს. თუ დაფიქრდებით, ჯერ უნდა გაარკვიოთ რა არის სინთეზი. ეს არის რაღაცის გაერთიანების პროცესი. ახალი დნმ-ის მოლეკულის ფორმირება ხდება რამდენიმე ეტაპად:
1) დნმ ტოპოიზომერაზა, რომელიც განლაგებულია რეპლიკაციის ჩანგლის წინ, ჭრის დნმ-ს, რათა ხელი შეუწყოს მის გადახვევას და განტვირთვას.
2) დნმ ჰელიკაზა, ტოპოიზომერაზას შემდეგ, მოქმედებს დნმ-ის სპირალის „გახსნის“ პროცესზე.
3) დნმ-ის შემაკავშირებელი პროტეინები ახორციელებენ დნმ-ის ჯაჭვების შეკავშირებას და ასევე ახორციელებენ მათ სტაბილიზაციას, ხელს უშლიან მათ ერთმანეთთან მიბმას.
4) დნმ პოლიმერაზა δ(დელტა) რეპლიკაციის ჩანგლის მოძრაობის სიჩქარესთან კოორდინირებული, ასრულებს სინთეზსწამყვანიჯაჭვებიშვილობილი დნმ მატრიცაზე 5" → 3" მიმართულებითდედობრივი დნმ-ის ჯაჭვები მისი 3" ბოლოდან 5" ბოლომდე (სიჩქარე 100 ბაზის წყვილამდე წამში). ეს მოვლენები ამის შესახებ დედობრივიდნმ-ის ჯაჭვები შეზღუდულია.

ბრინჯი. 9. დნმ-ის რეპლიკაციის პროცესის სქემატური წარმოდგენა: (1) ჩამორჩენილი ჯაჭვი (ლაგ ჯაჭვი), (2) წამყვანი ჯაჭვი (წამყვანი ჯაჭვი), (3) დნმ პოლიმერაზა α (Polα), (4) დნმ ლიგაზა, (5) რნმ -პრაიმერი, (6) პრიმაზა, (7) ოკაზაკის ფრაგმენტი, (8) დნმ პოლიმერაზა δ (Polδ), (9) ჰელიკაზა, (10) ერთჯაჭვიანი დნმ-ის დამაკავშირებელი ცილები, (11) ტოპოიზომერაზა.
ჩამორჩენილი ქალიშვილის დნმ-ის ჯაჭვის სინთეზი აღწერილია ქვემოთ (იხ. ქვემოთ). სქემარეპლიკაციის ჩანგალი და რეპლიკაციის ფერმენტების ფუნქცია)
დნმ-ის რეპლიკაციის შესახებ მეტი ინფორმაციისთვის იხ
5) მშობელი მოლეკულის სხვა ჯაჭვის გახსნისა და სტაბილიზაციისთანავე ის უერთდებადნმ პოლიმერაზა α(ალფა)და მიმართულებით 5 "→3" სინთეზირებს პრაიმერი (რნმ პრაიმერი) - რნმ-ის თანმიმდევრობა დნმ-ის შაბლონზე, რომლის სიგრძეა 10-დან 200 ნუკლეოტიდამდე. ამის შემდეგ ფერმენტიამოღებულია დნმ-ის ჯაჭვიდან.
Იმის მაგივრად დნმ პოლიმერაზაα
მიმაგრებულია პრაიმერის 3" ბოლოზედნმ პოლიმერაზაε
.
6)
დნმ პოლიმერაზაε
(ეპსილონი) თითქოს აგრძელებს პრაიმერის გახანგრძლივებას, მაგრამ როგორც სუბსტრატის ჩაშენებადეზოქსირიბონუკლეოტიდები(150-200 ნუკლეოტიდის ოდენობით). შედეგად, მყარი ძაფი იქმნება ორი ნაწილისგან -რნმ(ანუ პრაიმერი) და დნმ.
დნმ პოლიმერაზა εმუშაობს მანამ, სანამ არ შეხვდება წინა პრაიმერსფრაგმენტი ოკაზაკი(ცოტა ადრე სინთეზირებულია). შემდეგ ეს ფერმენტი ამოღებულია ჯაჭვიდან.
7) დნმ პოლიმერაზა β(ბეტა) დგას ნაცვლადდნმ პოლიმერაზები ε,მოძრაობს იმავე მიმართულებით (5" → 3") და შლის პრაიმერის რიბონუკლეოტიდებს მათ ადგილას დეზოქსირიბონუკლეოტიდების ჩასმისას. ფერმენტი მუშაობს პრაიმერის სრულ მოცილებამდე, ე.ი. დეზოქსირიბონუკლეოტიდამდე (კიდევ უფრო ადრე სინთეზირებულიდნმ პოლიმერაზა ε). ფერმენტს არ შეუძლია დააკავშიროს თავისი მუშაობის შედეგი და წინა დნმ, ამიტომ ის ტოვებს ჯაჭვს.
შედეგად, ქალიშვილის დნმ-ის ფრაგმენტი დედა ძაფის მატრიცაზე „დევს“. მას ეძახიანოკაზაკის ფრაგმენტი.
8) დნმ ლიგაზა აკავშირებს ორ მიმდებარედ ოკაზაკის ფრაგმენტები , ე.ი. 5" - სეგმენტის დასასრული, სინთეზირებულიდნმ პოლიმერაზა ε,და 3" ჯაჭვის ბოლო ჩაშენებულიდნმ პოლიმერაზაβ .
რნმ-ის სტრუქტურა
რიბონუკლეინის მჟავა(რნმ) არის სამი ძირითადი მაკრომოლეკულიდან ერთ-ერთი (დანარჩენი ორი არის დნმ და ცილა), რომლებიც გვხვდება ყველა ცოცხალი ორგანიზმის უჯრედებში.
ისევე როგორც დნმ, რნმ შედგება გრძელი ჯაჭვისაგან, რომელშიც ყოველი რგოლი ეწოდება ნუკლეოტიდი. თითოეული ნუკლეოტიდი შედგება აზოტოვანი ფუძისგან, რიბოზის შაქრისა და ფოსფატის ჯგუფისგან. თუმცა, დნმ-ისგან განსხვავებით, რნმ-ს ჩვეულებრივ აქვს ერთი და არა ორი ჯაჭვი. რნმ-ში პენტოზა წარმოდგენილია რიბოზით და არა დეზოქსირიბოზით (რიბოზას აქვს დამატებითი ჰიდროქსილის ჯგუფი ნახშირწყლების მეორე ატომზე). დაბოლოს, დნმ განსხვავდება რნმ-ისგან აზოტოვანი ბაზების შემადგენლობით: თიმინის ნაცვლად ( თ) ურაცილი არის რნმ-ში ( U) , რომელიც ასევე ავსებს ადენინს.
ნუკლეოტიდების თანმიმდევრობა რნმ-ს გენეტიკური ინფორმაციის კოდირების საშუალებას აძლევს. ყველა ფიჭური ორგანიზმი იყენებს რნმ-ს (mRNA) ცილების სინთეზის დასაპროგრამებლად.
უჯრედული რნმ წარმოიქმნება პროცესში ე.წ ტრანსკრიფცია , ანუ რნმ-ის სინთეზი დნმ-ის შაბლონზე, რომელიც ხორციელდება სპეციალური ფერმენტების მიერ - რნმ პოლიმერაზები.
მესინჯერი რნმ-ები (mRNA) შემდეგ მონაწილეობენ პროცესში ე.წ გადაცემა, იმათ. ცილის სინთეზი mRNA შაბლონზე რიბოზომების მონაწილეობით. სხვა რნმ-ები ტრანსკრიფციის შემდეგ განიცდიან ქიმიურ მოდიფიკაციას და მეორადი და მესამეული სტრუქტურების წარმოქმნის შემდეგ ასრულებენ ფუნქციებს, რომლებიც დამოკიდებულია რნმ-ის ტიპზე.

ბრინჯი. 10. განსხვავება დნმ-სა და რნმ-ს შორის აზოტოვანი ფუძის მიხედვით: თიმინის (T) ნაცვლად რნმ შეიცავს ურაცილს (U), რომელიც ასევე ავსებს ადენინს.
ტრანსკრიპცია
ეს არის რნმ-ის სინთეზის პროცესი დნმ-ის შაბლონზე. დნმ იხსნება ერთ-ერთ ადგილზე. ერთ-ერთი ჯაჭვი შეიცავს ინფორმაციას, რომელიც საჭიროებს კოპირებას რნმ-ის მოლეკულაზე - ამ ჯაჭვს კოდირებას უწოდებენ. დნმ-ის მეორე ჯაჭვს, რომელიც ავსებს კოდირების ჯაჭვს, ეწოდება შაბლონი. შაბლონის ჯაჭვზე ტრანსკრიფციის პროცესში 3'-5' მიმართულებით (დნმ-ის ჯაჭვის გასწვრივ) სინთეზირდება მისი დამატებითი რნმ-ის ჯაჭვი. ამრიგად, იქმნება კოდირების ჯაჭვის რნმ-ის ასლი.
![]()
ბრინჯი. 11. ტრანსკრიფციის სქემატური წარმოდგენა
მაგალითად, თუ გვეძლევა კოდირების სტრიქონის თანმიმდევრობა
3'-ATGTCCTAGCTGCTCG - 5',
შემდეგ, კომპლემენტარობის წესის მიხედვით, მატრიცის ჯაჭვი ატარებს თანმიმდევრობას
5'- TACAGGATCGACGAGC- 3',
და მისგან სინთეზირებული რნმ არის თანმიმდევრობა
გადაცემა
განიხილეთ მექანიზმი ცილის სინთეზირნმ-ის მატრიცაზე, ასევე გენეტიკურ კოდსა და მის თვისებებზე. ასევე, სიცხადისთვის, ქვემოთ მოცემულ ბმულზე, გირჩევთ უყუროთ მოკლე ვიდეოს ცოცხალ უჯრედში მიმდინარე ტრანსკრიფციისა და თარგმნის პროცესების შესახებ:

ბრინჯი. 12. ცილის სინთეზის პროცესი: დნმ კოდირებს რნმ-ს, რნმ კოდირებს ცილას
გენეტიკური კოდი
გენეტიკური კოდი- ცილების ამინომჟავების თანმიმდევრობის კოდირების მეთოდი ნუკლეოტიდების თანმიმდევრობის გამოყენებით. თითოეული ამინომჟავა დაშიფრულია სამი ნუკლეოტიდის თანმიმდევრობით - კოდონი ან ტრიპლეტი.
გენეტიკური კოდი საერთოა პრო- და ევკარიოტების უმეტესობისთვის. ცხრილში მოცემულია 64-ვე კოდონი და ჩამოთვლილია შესაბამისი ამინომჟავები. საბაზისო რიგი არის mRNA-ს 5"-დან 3"-მდე ბოლომდე.
ცხრილი 1. სტანდარტული გენეტიკური კოდი
|
1-ლი არა |
მე-2 ბაზა |
მე-3 არა |
|||||||
|
U |
C |
ა |
გ |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(სერ/ს) |
U A U |
(წ/წ) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(ლეუ/ლ) |
U C A |
U A A |
შეაჩერე კოდონი** |
U G A |
შეაჩერე კოდონი** |
ა |
||
|
U U G |
U C G |
U A G |
შეაჩერე კოდონი** |
U G G |
(Trp/W) |
გ |
|||
|
C |
C U U |
C C U |
(პრო/პ) |
C A U |
(მისი/ჰ) |
C G U |
(არგ/რ) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
CGA |
ა |
||||
|
C U G |
C C G |
C A G |
C G G |
გ |
|||||
|
ა |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(სერ/ს) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
ა |
||||
|
A U G |
(მეტ/მ) |
A C G |
A A G |
A G G |
გ |
||||
|
გ |
G U U |
(Val/V) |
G C U |
(ალა/ა) |
G A U |
(Asp/D) |
გ გ უ |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
გ უ ა |
G C A |
G A A |
(გლუ/ე) |
G G A |
ა |
||||
|
გ უ გ |
G C G |
G A G |
G G G |
გ |
|||||
სამეულებს შორის არის 4 სპეციალური თანმიმდევრობა, რომლებიც მოქმედებს როგორც "პუნქტუაციის ნიშნები":
- *სამმაგი აუგ, ასევე მეთიონინის კოდირებას, ე.წ დაწყების კოდონი. ეს კოდონი იწყებს ცილის მოლეკულის სინთეზს. ამრიგად, ცილის სინთეზის დროს, პირველი ამინომჟავა თანმიმდევრობით ყოველთვის იქნება მეთიონინი.
- ** სამეული UAA, UAGდა UGAდაურეკა შეაჩერე კოდონებიდა არ კოდებს რაიმე ამინომჟავას. ამ თანმიმდევრობებზე ცილის სინთეზი ჩერდება.
გენეტიკური კოდის თვისებები
1. სამეული. თითოეული ამინომჟავა კოდირებულია სამი ნუკლეოტიდის თანმიმდევრობით - ტრიპლეტი ან კოდონი.
2. უწყვეტობა. სამეულებს შორის დამატებითი ნუკლეოტიდები არ არის, ინფორმაცია მუდმივად იკითხება.
3. გადახურვის გარეშე. ერთი ნუკლეოტიდი არ შეიძლება იყოს ერთდროულად ორი სამეულის ნაწილი.
4. უნიკალურობა. ერთ კოდონს შეუძლია მხოლოდ ერთი ამინომჟავის კოდირება.

5. დეგენერაცია. ერთი ამინომჟავა შეიძლება დაშიფრული იყოს რამდენიმე სხვადასხვა კოდონით.

6. მრავალმხრივობა. გენეტიკური კოდი ყველა ცოცხალი ორგანიზმისთვის ერთნაირია.
მაგალითი. ჩვენ მოცემულია კოდირების სტრიქონის თანმიმდევრობა:
3’- CCGATTGCACGTCGATCGTATA- 5’.
მატრიცის ჯაჭვს ექნება თანმიმდევრობა:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
ახლა ჩვენ „ასინთეზირებთ“ ინფორმაციულ რნმ-ს ამ ჯაჭვიდან:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
პროტეინის სინთეზი მიდის 5' → 3' მიმართულებით, ამიტომ, გენეტიკური კოდის "წაკითხვისთვის" უნდა გადავატრიალოთ თანმიმდევრობა:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
ახლა იპოვნეთ საწყისი კოდონი AUG:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
დაყავით თანმიმდევრობა სამეულებად:
ასე ჟღერს: დნმ-დან ინფორმაცია გადადის რნმ-ში (ტრანსკრიფცია), რნმ-დან ცილაში (თარგმანი). დნმ-ის დუბლირება შესაძლებელია რეპლიკაციითაც და საპირისპირო ტრანსკრიფციის პროცესიც შესაძლებელია, როდესაც დნმ სინთეზირდება რნმ-ის შაბლონიდან, მაგრამ ასეთი პროცესი ძირითადად ვირუსებისთვისაა დამახასიათებელი.

ბრინჯი. 13. ცენტრალური დოგმა მოლეკულური ბიოლოგია
გენომი: გენები და ქრომოსომა
(ზოგადი ცნებები)
გენომი - ორგანიზმის ყველა გენის მთლიანობა; მისი სრული ქრომოსომული ნაკრები.
ტერმინი „გენომი“ შემოგვთავაზა გ. ვინკლერმა 1920 წელს იმავე ბიოლოგიური სახეობის ორგანიზმების ქრომოსომების ჰაპლოიდურ კომპლექტში შემავალი გენების მთლიანობის აღსაწერად. ამ ტერმინის თავდაპირველი მნიშვნელობა მიუთითებს იმაზე, რომ გენომის კონცეფცია, გენოტიპისგან განსხვავებით, არის მთლიანი სახეობის გენეტიკური მახასიათებელი და არა ცალკეული. მოლეკულური გენეტიკის განვითარებასთან ერთად, ამ ტერმინის მნიშვნელობა შეიცვალა. ცნობილია, რომ დნმ, რომელიც ორგანიზმების უმეტესობაში გენეტიკური ინფორმაციის მატარებელია და, შესაბამისად, გენომის საფუძველს ქმნის, მოიცავს არა მხოლოდ გენებს ამ სიტყვის თანამედროვე გაგებით. უმეტესობაევკარიოტული უჯრედების დნმ წარმოდგენილია არაკოდიციური ("ზედმეტი") ნუკლეოტიდური თანმიმდევრობებით, რომლებიც არ შეიცავს ინფორმაციას ცილებისა და ნუკლეინის მჟავების შესახებ. ამრიგად, ნებისმიერი ორგანიზმის გენომის ძირითადი ნაწილია მისი ქრომოსომების ჰაპლოიდური ნაკრების მთელი დნმ.
გენები არის დნმ-ის მოლეკულების სეგმენტები, რომლებიც კოდირებენ პოლიპეპტიდებს და რნმ-ის მოლეკულებს.
გასული საუკუნის განმავლობაში, გენების ჩვენი გაგება მნიშვნელოვნად შეიცვალა. ადრე, გენომი იყო ქრომოსომის რეგიონი, რომელიც კოდირებს ან განსაზღვრავს ერთ მახასიათებელს ან ფენოტიპური(ხილული) თვისება, როგორიცაა თვალის ფერი.
1940 წელს ჯორჯ ბიდლმა და ედვარდ ტატამმა შემოგვთავაზეს გენის მოლეკულური განმარტება. მეცნიერებმა დაამუშავეს სოკოს სპორები ნეიროსპორა კრასარენტგენი და სხვა აგენტები, რომლებიც იწვევენ ცვლილებებს დნმ-ის თანმიმდევრობაში ( მუტაციები), და აღმოაჩინა სოკოს მუტანტური შტამები, რომლებმაც დაკარგეს გარკვეული სპეციფიკური ფერმენტები, რამაც ზოგიერთ შემთხვევაში გამოიწვია მთელი მეტაბოლური გზის დარღვევა. Beadle და Tatham მივიდნენ დასკვნამდე, რომ გენი არის გენეტიკური მასალის ნაწილი, რომელიც განსაზღვრავს ან კოდირებს ერთ ფერმენტს. ასეა ჰიპოთეზა "ერთი გენი, ერთი ფერმენტი". ეს კონცეფცია მოგვიანებით გაფართოვდა განსაზღვრებამდე "ერთი გენი - ერთი პოლიპეპტიდი"რადგან ბევრი გენი აკოდირებს ცილებს, რომლებიც არ არიან ფერმენტები და პოლიპეპტიდი შეიძლება იყოს რთული ცილის კომპლექსის ქვედანაყოფი.
ნახ. 14 გვიჩვენებს დიაგრამას, თუ როგორ განსაზღვრავს დნმ-ის სამეულები პოლიპეპტიდს, ცილის ამინომჟავის თანმიმდევრობას, შუამავლობით mRNA. დნმ-ის ერთ-ერთი ჯაჭვი ასრულებს mRNA-ს სინთეზის შაბლონის როლს, რომლის ნუკლეოტიდური ტრიპლეტები (კოდონები) ავსებენ დნმ-ის სამეულს. ზოგიერთ ბაქტერიაში და ბევრ ევკარიოტში, კოდირების თანმიმდევრობა წყდება არაკოდირების რეგიონებით (ე.წ. ინტრონები).
გენის თანამედროვე ბიოქიმიური განმარტება კიდევ უფრო კონკრეტულად. გენები არის დნმ-ის ყველა განყოფილება, რომელიც კოდირებს საბოლოო პროდუქტების პირველადი თანმიმდევრობას, რომელიც მოიცავს პოლიპეპტიდებს ან რნმ-ს, რომლებსაც აქვთ სტრუქტურული ან კატალიზური ფუნქცია.
გენებთან ერთად დნმ ასევე შეიცავს სხვა თანმიმდევრობებს, რომლებიც ასრულებენ ექსკლუზიურად მარეგულირებელ ფუნქციას. მარეგულირებელი თანმიმდევრობებიშეიძლება აღინიშნოს გენების დასაწყისი ან დასასრული, გავლენა მოახდინოს ტრანსკრიფციაზე ან მიუთითოს რეპლიკაციის ან რეკომბინაციის დაწყების ადგილი. ზოგიერთი გენი შეიძლება გამოიხატოს სხვადასხვა გზით, დნმ-ის ერთი და იგივე ნაწილი სხვადასხვა პროდუქტის ფორმირების შაბლონად ემსახურება.
ჩვენ შეგვიძლია უხეშად გამოვთვალოთ გენის მინიმალური ზომაშუალედური ცილის კოდირება. თითოეული ამინომჟავა პოლიპეპტიდურ ჯაჭვში კოდირებულია სამი ნუკლეოტიდის თანმიმდევრობით; ამ სამეულების (კოდონების) თანმიმდევრობა შეესაბამება მოცემული გენის მიერ კოდირებულ პოლიპეპტიდში ამინომჟავების ჯაჭვს. 350 ამინომჟავის ნარჩენებისგან შემდგარი პოლიპეპტიდური ჯაჭვი (საშუალო სიგრძის ჯაჭვი) შეესაბამება 1050 bp თანმიმდევრობას. ( ბპ). თუმცა, ბევრი ევკარიოტული გენი და ზოგიერთი პროკარიოტული გენი წყდება დნმ-ის სეგმენტებით, რომლებიც არ ინფორმაციის მატარებლებიცილის შესახებ და, შესაბამისად, გაცილებით გრძელი აღმოჩნდება, ვიდრე მარტივი გაანგარიშება აჩვენებს.
რამდენი გენია ერთ ქრომოსომაზე?
 ბრინჯი. 15. ქრომოსომების ხედი პროკარიოტულ (მარცხნივ) და ევკარიოტულ უჯრედებში. ჰისტონები არის ბირთვული ცილების ფართო კლასი, რომლებიც ასრულებენ ორ ძირითად ფუნქციას: ისინი მონაწილეობენ ბირთვში დნმ-ის ჯაჭვების შეფუთვაში და ბირთვული პროცესების ეპიგენეტიკურ რეგულირებაში, როგორიცაა ტრანსკრიფცია, რეპლიკაცია და შეკეთება.
ბრინჯი. 15. ქრომოსომების ხედი პროკარიოტულ (მარცხნივ) და ევკარიოტულ უჯრედებში. ჰისტონები არის ბირთვული ცილების ფართო კლასი, რომლებიც ასრულებენ ორ ძირითად ფუნქციას: ისინი მონაწილეობენ ბირთვში დნმ-ის ჯაჭვების შეფუთვაში და ბირთვული პროცესების ეპიგენეტიკურ რეგულირებაში, როგორიცაა ტრანსკრიფცია, რეპლიკაცია და შეკეთება.
პროკარიოტების დნმ უფრო მარტივია: მათ უჯრედებს არ აქვთ ბირთვი, ამიტომ დნმ მდებარეობს უშუალოდ ციტოპლაზმაში ნუკლეოიდის სახით.
 მოგეხსენებათ, ბაქტერიულ უჯრედებს აქვთ ქრომოსომა დნმ-ის ჯაჭვის სახით, რომელიც შეფუთულია კომპაქტურ სტრუქტურაში - ნუკლეოიდში. პროკარიოტული ქრომოსომა ეშერიხია კოლი, რომლის გენომი მთლიანად გაშიფრულია, არის წრიული დნმ-ის მოლეკულა (ფაქტობრივად, ეს არ არის მარჯვენა წრე, არამედ მარყუჟი დასაწყისისა და დასასრულის გარეშე), რომელიც შედგება 4,639,675 b.p. ეს თანმიმდევრობა შეიცავს დაახლოებით 4300 ცილის გენს და კიდევ 157 გენს სტაბილური რნმ-ის მოლეკულებისთვის. IN ადამიანის გენომიდაახლოებით 3,1 მილიარდი ბაზის წყვილი, რომელიც შეესაბამება თითქმის 29000 გენს, რომლებიც განლაგებულია 24 სხვადასხვა ქრომოსომაზე.
მოგეხსენებათ, ბაქტერიულ უჯრედებს აქვთ ქრომოსომა დნმ-ის ჯაჭვის სახით, რომელიც შეფუთულია კომპაქტურ სტრუქტურაში - ნუკლეოიდში. პროკარიოტული ქრომოსომა ეშერიხია კოლი, რომლის გენომი მთლიანად გაშიფრულია, არის წრიული დნმ-ის მოლეკულა (ფაქტობრივად, ეს არ არის მარჯვენა წრე, არამედ მარყუჟი დასაწყისისა და დასასრულის გარეშე), რომელიც შედგება 4,639,675 b.p. ეს თანმიმდევრობა შეიცავს დაახლოებით 4300 ცილის გენს და კიდევ 157 გენს სტაბილური რნმ-ის მოლეკულებისთვის. IN ადამიანის გენომიდაახლოებით 3,1 მილიარდი ბაზის წყვილი, რომელიც შეესაბამება თითქმის 29000 გენს, რომლებიც განლაგებულია 24 სხვადასხვა ქრომოსომაზე.
პროკარიოტები (ბაქტერიები).
 ბაქტერია E. coliაქვს ერთი ორჯაჭვიანი წრიული დნმ-ის მოლეკულა. იგი შედგება 4,639,675 ბ.წ. და აღწევს სიგრძე დაახლოებით 1,7 მმ, რაც აღემატება თავად უჯრედის სიგრძეს E. coliდაახლოებით 850 ჯერ. გარდა დიდი წრიული ქრომოსომისა, როგორც ნუკლეოიდის ნაწილი, ბევრი ბაქტერია შეიცავს ერთ ან მეტ წრიულ დნმ-ის მოლეკულას, რომლებიც თავისუფლად მდებარეობს ციტოზოლში. ამ ექსტრაქრომოსომულ ელემენტებს ე.წ პლაზმიდები(სურ. 16).
ბაქტერია E. coliაქვს ერთი ორჯაჭვიანი წრიული დნმ-ის მოლეკულა. იგი შედგება 4,639,675 ბ.წ. და აღწევს სიგრძე დაახლოებით 1,7 მმ, რაც აღემატება თავად უჯრედის სიგრძეს E. coliდაახლოებით 850 ჯერ. გარდა დიდი წრიული ქრომოსომისა, როგორც ნუკლეოიდის ნაწილი, ბევრი ბაქტერია შეიცავს ერთ ან მეტ წრიულ დნმ-ის მოლეკულას, რომლებიც თავისუფლად მდებარეობს ციტოზოლში. ამ ექსტრაქრომოსომულ ელემენტებს ე.წ პლაზმიდები(სურ. 16).
პლაზმიდების უმეტესობა შედგება მხოლოდ რამდენიმე ათასი ბაზის წყვილისგან, ზოგი შეიცავს 10000 bp-ზე მეტს. ისინი ატარებენ გენეტიკურ ინფორმაციას და მრავლდებიან, რათა წარმოქმნან ქალიშვილური პლაზმიდები, რომლებიც შედიან ქალიშვილურ უჯრედებში მშობელი უჯრედის გაყოფის დროს. პლაზმიდები გვხვდება არა მხოლოდ ბაქტერიებში, არამედ საფუარში და სხვა სოკოებშიც. ხშირ შემთხვევაში, პლაზმიდები არ ანიჭებენ უპირატესობას მასპინძელ უჯრედებს და მათი ერთადერთი ამოცანაა დამოუკიდებლად გამრავლება. თუმცა, ზოგიერთი პლაზმიდი ატარებს მასპინძლისთვის სასარგებლო გენებს. მაგალითად, პლაზმიდებში შემავალ გენებს შეუძლიათ ბაქტერიული უჯრედების ანტიბაქტერიული აგენტებისადმი წინააღმდეგობის გაწევა. β-ლაქტამაზას გენის მატარებელი პლაზმიდები რეზისტენტობას ანიჭებენ β-ლაქტამურ ანტიბიოტიკებს, როგორიცაა პენიცილინი და ამოქსიცილინი. პლაზმიდებს შეუძლიათ ანტიბიოტიკებისადმი რეზისტენტული უჯრედებიდან გადავიდნენ იმავე ან სხვადასხვა ბაქტერიული სახეობის სხვა უჯრედებში, რაც იწვევს ამ უჯრედებს ასევე რეზისტენტულს. ანტიბიოტიკების ინტენსიური გამოყენება არის ძლიერი შერჩევითი ფაქტორი, რომელიც ხელს უწყობს ანტიბიოტიკების რეზისტენტობის მაკოდირებელი პლაზმიდების გავრცელებას (ისევე, როგორც ტრანსპოზონებს, რომლებიც კოდირებენ მსგავს გენებს) პათოგენურ ბაქტერიებს შორის და იწვევს ბაქტერიული შტამების წარმოქმნას რამდენიმე ანტიბიოტიკის მიმართ რეზისტენტობით. ექიმები იწყებენ ანტიბიოტიკების ფართო გამოყენების საშიშროების გააზრებას და დანიშნავენ მათ მხოლოდ მაშინ, როცა აბსოლუტურად აუცილებელია. მსგავსი მიზეზების გამო, ფერმის ცხოველების სამკურნალოდ ანტიბიოტიკების ფართო გამოყენება შეზღუდულია.
Იხილეთ ასევე: რავინ ნ.ვ., შესტაკოვი ს.ვ. პროკარიოტების გენომი // Vavilov Journal of Genetics and Breeding, 2013. V. 17. No 4/2. გვ 972-984.
ევკარიოტები.
ცხრილი 2. ზოგიერთი ორგანიზმის დნმ, გენები და ქრომოსომა
|
საერთო დნმ, ბ.წ. |
ქრომოსომების რაოდენობა* |
გენების სავარაუდო რაოდენობა |
|
|
ეშერიხია კოლი(ბაქტერია) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(საფუარი) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(ნემატოდი) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(მცენარე) |
119 186 200 |
33 000 |
|
|
დროზოფილა მელანოგასტერი(ხილის ბუზი) |
120 367 260 |
20 000 |
|
|
ორიზა სატივა(ბრინჯი) |
480 000 000 |
57 000 |
|
|
მუს კუნთი(მაუსი) |
2 634 266 500 |
27 000 |
|
|
ჰომო საპიენსი(ადამიანი) |
3 070 128 600 |
29 000 |
Შენიშვნა.ინფორმაცია მუდმივად განახლდება; დამატებითი უახლესი ინფორმაციისთვის ეწვიეთ ცალკეული გენომიური პროექტის ვებსაიტებს.
* ყველა ევკარიოტისთვის, საფუარის გარდა, მოცემულია ქრომოსომების დიპლოიდური ნაკრები. დიპლოიდურინაკრები ქრომოსომა (ბერძნული დიპლოოსიდან - ორმაგი და ეიდოსიდან - ხედი) - ქრომოსომების ორმაგი ნაკრები(2n), რომელთაგან თითოეულს აქვს თავისი ჰომოლოგია.
**ჰაპლოიდური ნაკრები. საფუარის ველურ შტამებს, როგორც წესი, აქვთ ამ ქრომოსომების რვა (ოქტაპლოიდური) ან მეტი ნაკრები.
*** ორი X ქრომოსომის მქონე ქალებისთვის. მამაკაცებს აქვთ X ქრომოსომა, მაგრამ არა Y, ანუ მხოლოდ 11 ქრომოსომა.
საფუარის უჯრედს, ერთ-ერთ ყველაზე პატარა ევკარიოტს, აქვს 2,6-ჯერ მეტი დნმ, ვიდრე უჯრედს. E. coli(ცხრილი 2). ხილის ბუზის უჯრედები დროზოფილაგენეტიკური კვლევის კლასიკური ობიექტი, შეიცავს 35-ჯერ მეტ დნმ-ს, ხოლო ადამიანის უჯრედები შეიცავს უჯრედებზე 700-ჯერ მეტ დნმ-ს. E. coli.ბევრი მცენარე და ამფიბია კიდევ უფრო მეტ დნმ-ს შეიცავს. ევკარიოტული უჯრედების გენეტიკური მასალა ორგანიზებულია ქრომოსომების სახით. ქრომოსომების დიპლოიდური ნაკრები (2 ნ) დამოკიდებულია ორგანიზმის ტიპზე (ცხრილი 2).
მაგალითად, ადამიანის სომატურ უჯრედში არის 46 ქრომოსომა ( ბრინჯი. 17). თითოეული ქრომოსომა ევკარიოტულ უჯრედში, როგორც ნაჩვენებია ნახ. 17, მაგრამ, შეიცავს ერთ ძალიან დიდ ორჯაჭვიან დნმ-ის მოლეკულას. ადამიანის ოცდაოთხი ქრომოსომა (22 დაწყვილებული ქრომოსომა და ორი სქესის ქრომოსომა X და Y) განსხვავდება სიგრძით 25-ჯერ მეტით. თითოეული ევკარიოტული ქრომოსომა შეიცავს გენების სპეციფიკურ კომპლექტს.

ბრინჯი. 17. ევკარიოტული ქრომოსომა.მაგრამ- წყვილი დაკავშირებული და შედედებული დის ქრომატიდიდან ადამიანის ქრომოსომიდან. ამ ფორმით, ევკარიოტული ქრომოსომა რჩება რეპლიკაციის შემდეგ და მეტაფაზაში მიტოზის დროს. ბ- ქრომოსომების სრული ნაკრები წიგნის ერთ-ერთი ავტორის ლეიკოციტიდან. თითოეული ნორმალური ადამიანის სომატური უჯრედი შეიცავს 46 ქრომოსომას.

დნმ-ის ზომა და ფუნქცია, როგორც მემკვიდრეობითი მასალის შესანახად და გადაცემის მატრიცა, ხსნის ამ მოლეკულის ორგანიზაციაში სპეციალური სტრუქტურული ელემენტების არსებობას. მაღალ ორგანიზმებში დნმ ნაწილდება ქრომოსომებს შორის.
ორგანიზმის დნმ-ის (ქრომოსომების) ერთობლიობას გენომი ეწოდება. ქრომოსომა განლაგებულია უჯრედის ბირთვში და ქმნის სტრუქტურას, რომელსაც ქრომატინი ეწოდება. ქრომატინი არის დნმ-ისა და ძირითადი ცილების (ჰისტონის) კომპლექსი 1:1 თანაფარდობით. დნმ-ის სიგრძე ჩვეულებრივ იზომება დამატებითი ნუკლეოტიდების წყვილის რაოდენობით (bp). მაგალითად, მე-3 ადამიანის ქრომოსომასაუკუნე არის დნმ-ის მოლეკულა, რომლის ზომაა 160 მილიონი bp. აქვს სიგრძე დაახლოებით 1 მმ, შესაბამისად, ადამიანის მე-3 ქრომოსომის ხაზოვანი მოლეკულა იქნება 5 მმ სიგრძის, ხოლო 23-ვე ქრომოსომის დნმ (~ 3 * 10 9 bp, MR = 1.8 * 10 12) ჰაპლოიდის. უჯრედი - კვერცხუჯრედი ან სპერმის უჯრედი - ხაზოვანი სახით იქნება 1 მ. ჩანასახის უჯრედების გარდა, ადამიანის სხეულის ყველა უჯრედი (დაახლოებით 1013 მათგანია) შეიცავს ქრომოსომების ორმაგ კომპლექტს. უჯრედის გაყოფის დროს დნმ-ის 46-ვე მოლეკულა რეპლიკაცია ხდება და რეორგანიზაცია ხდება 46 ქრომოსომად.
თუ თქვენ დააკავშირებთ ადამიანის გენომის დნმ-ის მოლეკულებს (22 ქრომოსომა და ქრომოსომა X და Y ან X და X) ერთმანეთთან, მიიღებთ დაახლოებით ერთი მეტრის სიგრძის თანმიმდევრობას. შენიშვნა: ყველა ძუძუმწოვარსა და სხვა ჰეტეროგამეტურ მამრობითი ორგანიზმში მდედრს აქვს ორი X ქრომოსომა (XX), ხოლო მამრს აქვს ერთი X ქრომოსომა და ერთი Y ქრომოსომა (XY).
ადამიანის უჯრედების უმეტესობა, ამიტომ ასეთი უჯრედების დნმ-ის მთლიანი სიგრძე დაახლოებით 2 მ-ია. ზრდასრულ ადამიანს აქვს დაახლოებით 10 14 უჯრედი, ამიტომ დნმ-ის ყველა მოლეკულის მთლიანი სიგრძეა 2-10 11 კმ. შედარებისთვის, დედამიწის გარშემოწერილობა არის 4~104 კმ, ხოლო დედამიწიდან მზემდე მანძილი 1,5~108 კმ. აი, საოცრად კომპაქტურად შეფუთული დნმ არის ჩვენს უჯრედებში!
ევკარიოტულ უჯრედებში არის დნმ-ის შემცველი სხვა ორგანელები - ეს არის მიტოქონდრია და ქლოროპლასტები. მრავალი ჰიპოთეზა წამოაყენეს მიტოქონდრიული და ქლოროპლასტის დნმ-ის წარმოშობასთან დაკავშირებით. დღეს საყოველთაოდ მიღებული თვალსაზრისი არის ის, რომ ისინი უძველესი ბაქტერიების ქრომოსომების საფუძვლებია, რომლებიც შეაღწიეს მასპინძელი უჯრედების ციტოპლაზმაში და გახდნენ ამ ორგანელების წინამორბედები. მიტოქონდრიული დნმ კოდირებს მიტოქონდრიულ tRNA-ს და rRNA-ს, ასევე რამდენიმე მიტოქონდრიულ ცილას. მიტოქონდრიული ცილების 95%-ზე მეტი კოდირებულია ბირთვული დნმ-ით.
გენების სტრუქტურა
განვიხილოთ გენის სტრუქტურა პროკარიოტებში და ევკარიოტებში, მათი მსგავსება და განსხვავებები. იმისდა მიუხედავად, რომ გენი არის დნმ-ის განყოფილება, რომელიც აკოდირებს მხოლოდ ერთ პროტეინს ან რნმ-ს, გარდა პირდაპირი კოდირების ნაწილისა, იგი ასევე შეიცავს მარეგულირებელ და სხვა სტრუქტურულ ელემენტებს, რომლებსაც აქვთ განსხვავებული სტრუქტურა პროკარიოტებში და ევკარიოტებში.
კოდირების თანმიმდევრობა- გენის მთავარი სტრუქტურული და ფუნქციური ერთეული, მასშია ნუკლეოტიდების სამეული, რომელიც კოდირებსამინომჟავების თანმიმდევრობა. ის იწყება საწყისი კოდონით და მთავრდება გაჩერების კოდონით.
კოდირების თანმიმდევრობამდე და მის შემდეგ არის გადაუთარგმნელი 5' და 3' თანმიმდევრობები. ისინი ასრულებენ მარეგულირებელ და დამხმარე ფუნქციებს, მაგალითად, უზრუნველყოფენ რიბოსომის დაშვებას mRNA-ზე.
გადაუთარგმნელი და კოდირებული თანმიმდევრობები ქმნიან ტრანსკრიფციის ერთეულს - ტრანსკრიბირებული დნმ-ის რეგიონს, ანუ დნმ-ის რეგიონს, საიდანაც სინთეზირებულია mRNA.
ტერმინატორიდნმ-ის არატრანსკრიბირებული რეგიონი გენის ბოლოს, სადაც რნმ-ის სინთეზი ჩერდება.
გენის დასაწყისში არის მარეგულირებელი ტერიტორია, რომელიც შეიცავს პრომოუტერიდა ოპერატორი.
პრომოუტერი- თანმიმდევრობა, რომლითაც პოლიმერაზა უკავშირდება ტრანსკრიფციის დაწყებისას. ოპერატორი- ეს ის უბანია, რომელსაც სპეციალური პროტეინები შეიძლება დაუკავშირდეს - რეპრესორები, რომელსაც შეუძლია შეამციროს რნმ-ის სინთეზის აქტივობა ამ გენიდან - სხვა სიტყვებით რომ ვთქვათ, შეამციროს იგი გამოხატულება.
გენების სტრუქტურა პროკარიოტებში
პროკარიოტებში და ევკარიოტებში გენების სტრუქტურის ზოგადი გეგმა არ განსხვავდება - ორივე მათგანი შეიცავს მარეგულირებელ რეგიონს პრომოტორთან და ოპერატორთან, ტრანსკრიპციის ერთეულს კოდირების და არათარგმნილი თანმიმდევრობით და ტერმინატორთან. თუმცა, გენების ორგანიზაცია პროკარიოტებსა და ევკარიოტებში განსხვავებულია.

ბრინჯი. 18. გენის სტრუქტურის სქემა პროკარიოტებში (ბაქტერიებში) -სურათი გადიდებულია
ოპერონის დასაწყისში და ბოლოს არის საერთო მარეგულირებელი რეგიონები რამდენიმე სტრუქტურული გენისთვის. ოპერონის ტრანსკრიბირებული რეგიონიდან იკითხება ერთი mRNA მოლეკულა, რომელიც შეიცავს რამდენიმე კოდირების თანმიმდევრობას, რომელთაგან თითოეულს აქვს საკუთარი საწყისი და გაჩერების კოდონი. თითოეული ამ სფეროდანსინთეზირდება ერთი ცილა. Ამგვარად, რამდენიმე ცილის მოლეკულა სინთეზირდება ერთი i-RNA მოლეკულიდან.
პროკარიოტებს ახასიათებთ რამდენიმე გენის გაერთიანება ერთ ფუნქციურ ერთეულში - ოპერონი. ოპერონის მუშაობა შეიძლება დარეგულირდეს სხვა გენებით, რომლებიც შესამჩნევად შეიძლება მოიხსნას თავად ოპერონიდან - რეგულატორები. ამ გენიდან ნათარგმნ ცილას ე.წ რეპრესორი. ის აკავშირებს ოპერონის ოპერატორს, არეგულირებს მასში შემავალი ყველა გენის ექსპრესიას ერთდროულად.
ფენომენი ახასიათებს პროკარიოტებსაც ტრანსკრიფცია და თარგმანის კონიუგაციები.
![]()
ბრინჯი. 19 პროკარიოტებში ტრანსკრიფციისა და თარგმანის კონიუგაციის ფენომენი - სურათი გადიდებულია
ეს დაწყვილება არ ხდება ევკარიოტებში ბირთვული კონვერტის არსებობის გამო, რომელიც გამოყოფს ციტოპლაზმას, სადაც ხდება ტრანსლაცია, გენეტიკური მასალისგან, რომელზედაც ხდება ტრანსკრიფცია. პროკარიოტებში, დნმ-ის შაბლონზე რნმ-ის სინთეზის დროს, რიბოსომა შეიძლება დაუყოვნებლივ დაუკავშირდეს სინთეზირებულ რნმ-ის მოლეკულას. ამრიგად, თარგმანი იწყება ტრანსკრიფციის დასრულებამდეც კი. უფრო მეტიც, რამდენიმე რიბოსომას შეუძლია ერთდროულად დაუკავშირდეს რნმ-ის ერთ მოლეკულას, სინთეზირდეს ერთი ცილის რამდენიმე მოლეკულას ერთდროულად.
გენების სტრუქტურა ევკარიოტებში
ევკარიოტების გენები და ქრომოსომა ძალიან კომპლექსურად არის ორგანიზებული.
მრავალი სახეობის ბაქტერიას აქვს მხოლოდ ერთი ქრომოსომა და თითქმის ყველა შემთხვევაში თითოეულ ქრომოსომაზე თითოეული გენის თითო ასლია. მხოლოდ რამდენიმე გენი, როგორიცაა rRNA გენი, შეიცავს მრავალ ასლს. გენები და მარეგულირებელი თანმიმდევრობები ქმნიან პროკარიოტების თითქმის მთელ გენომს. უფრო მეტიც, თითქმის ყველა გენი მკაცრად შეესაბამება ამინომჟავების თანმიმდევრობას (ან რნმ-ის თანმიმდევრობას), რომელსაც ის კოდირებს (ნახ. 14).
ევკარიოტული გენების სტრუქტურული და ფუნქციური ორგანიზაცია გაცილებით რთულია. ევკარიოტული ქრომოსომების შესწავლამ და შემდგომში ევკარიოტული გენომის სრული თანმიმდევრობის თანმიმდევრობამ ბევრი სიურპრიზი გამოიწვია. ბევრ, თუ არა უმეტეს ევკარიოტულ გენს აქვს საინტერესო თვისება: მათი ნუკლეოტიდური თანმიმდევრობები შეიცავს ერთ ან მეტ დნმ რეგიონს, რომელიც არ აკოდირებს პოლიპეპტიდური პროდუქტის ამინომჟავების თანმიმდევრობას. ასეთი არათარგმნილი ჩანართები არღვევს უშუალო შესაბამისობას გენის ნუკლეოტიდურ თანმიმდევრობასა და კოდირებული პოლიპეპტიდის ამინომჟავების თანმიმდევრობას შორის. გენებში ამ გადაუთარგმნელ სეგმენტებს ე.წ ინტრონები, ან ჩაშენებული თანმიმდევრობებიდა კოდირების სეგმენტებია ეგზონები. პროკარიოტებში მხოლოდ რამდენიმე გენი შეიცავს ინტრონებს.
ასე რომ, ევკარიოტებში პრაქტიკულად არ არსებობს გენების კომბინაცია ოპერონებში და ევკარიოტული გენის კოდირების თანმიმდევრობა ყველაზე ხშირად იყოფა ნათარგმნ რეგიონებად. - ეგზონებიდა გადაუთარგმნელი სექციები - ინტრონები.
უმეტეს შემთხვევაში, ინტრონების ფუნქცია დადგენილი არ არის. ზოგადად, ადამიანის დნმ-ის მხოლოდ 1,5% არის „კოდირებელი“, ანუ ის ატარებს ინფორმაციას ცილების ან რნმ-ის შესახებ. თუმცა, დიდი ინტრონების გათვალისწინებით, გამოდის, რომ ადამიანის დნმ-ის 30% გენებისგან შედგება. ვინაიდან გენები შეადგენენ ადამიანის გენომის შედარებით მცირე ნაწილს, დნმ-ის მნიშვნელოვანი რაოდენობა გამოუკვლეველია.

ბრინჯი. 16. გენის სტრუქტურის სქემა ევკარიოტებში - სურათი გადიდებულია
თითოეული გენიდან პირველად სინთეზირდება გაუაზრებელი, ანუ პრე-რნმ, რომელიც შეიცავს ინტრონებსაც და ეგზონებსაც.
ამის შემდეგ ხდება სპლაისინგის პროცესი, რის შედეგადაც ხდება ინტრონის უბნების ამოკვეთა და წარმოიქმნება მომწიფებული mRNA, საიდანაც შესაძლებელია ცილის სინთეზირება.

ბრინჯი. 20. ალტერნატიული შერწყმის პროცესი - სურათი გადიდებულია
გენების ასეთი ორგანიზაცია საშუალებას იძლევა, მაგალითად, როდესაც შესაძლებელია ცილის სხვადასხვა ფორმის სინთეზირება ერთი გენიდან, იმის გამო, რომ ეგზონები შეიძლება შერწყმული იყოს სხვადასხვა თანმიმდევრობით შერწყმის დროს.

ბრინჯი. 21. განსხვავებები პროკარიოტებისა და ევკარიოტების გენების აგებულებაში - სურათი გადიდებულია
მუტაციები და მუტაგენეზი
მუტაციაეწოდება გენოტიპის მუდმივი ცვლილება, ანუ ნუკლეოტიდური თანმიმდევრობის ცვლილება.
პროცესს, რომელიც იწვევს მუტაციას ე.წ მუტაგენეზიდა ორგანიზმი ყველარომლის უჯრედები ატარებენ ერთსა და იმავე მუტაციას მუტანტი.
მუტაციის თეორიაპირველად ჩამოაყალიბა ჰიუ დე ვრისმა 1903 წელს. მისი თანამედროვე ვერსია მოიცავს შემდეგ დებულებებს:
1. მუტაციები ხდება მოულოდნელად, მოულოდნელად.
2. მუტაციები თაობიდან თაობას გადაეცემა.
3. მუტაციები შეიძლება იყოს სასარგებლო, მავნე ან ნეიტრალური, დომინანტური ან რეცესიული.
4. მუტაციების გამოვლენის ალბათობა დამოკიდებულია შესწავლილი ინდივიდების რაოდენობაზე.
5. მსგავსი მუტაციები შეიძლება განმეორებით მოხდეს.
6. მუტაციები არ არის მიმართული.
მუტაციები შეიძლება მოხდეს სხვადასხვა ფაქტორების გავლენის ქვეშ. განასხვავებენ გამოწვეულ მუტაციებს მუტაგენური ზემოქმედებები: ფიზიკური (მაგ. ულტრაიისფერი ან გამოსხივება), ქიმიური (მაგ. კოლხიცინი ან რეაქტიული ჟანგბადის სახეობები) და ბიოლოგიური (მაგ. ვირუსები). მუტაციებიც შეიძლება გამოიწვიოს რეპლიკაციის შეცდომები.
მუტაციების გაჩენის პირობებიდან გამომდინარე იყოფა სპონტანური- ანუ ნორმალურ პირობებში წარმოქმნილი მუტაციები და გამოწვეული- ანუ მუტაციები, რომლებიც წარმოიშვა განსაკუთრებულ პირობებში.
მუტაციები შეიძლება მოხდეს არა მხოლოდ ბირთვულ დნმ-ში, არამედ, მაგალითად, მიტოქონდრიის ან პლასტიდების დნმ-ში. შესაბამისად შეგვიძლია გამოვყოთ ბირთვულიდა ციტოპლაზმურიმუტაციები.
მუტაციების გაჩენის შედეგად ხშირად შეიძლება გამოჩნდეს ახალი ალელები. თუ მუტანტის ალელი აჭარბებს ნორმალურ ალელს, მუტაცია ეწოდება დომინანტური. თუ ნორმალური ალელი თრგუნავს მუტაციურს, მუტაცია ეწოდება რეცესიული. მუტაციების უმეტესობა, რომლებიც წარმოქმნიან ახალ ალელებს, რეცესიულია.
მუტაციები გამოირჩევიან ეფექტით ადაპტაციურირაც იწვევს ორგანიზმის გარემოსთან ადაპტაციის მატებას, ნეიტრალურირაც გავლენას არ ახდენს გადარჩენაზე მავნერომლებიც ამცირებენ ორგანიზმების ადაპტირებას გარემო პირობებთან და სასიკვდილოიწვევს ორგანიზმის სიკვდილს განვითარების ადრეულ ეტაპზე.
შედეგების მიხედვით განასხვავებენ მუტაციებს, რაც იწვევს ცილის ფუნქციის დაკარგვა, მუტაციებს იწვევს გაჩენა პროტეინს აქვს ახალი ფუნქცია, ისევე როგორც მუტაციები, რომლებიც გენის დოზის შეცვლადა, შესაბამისად, მისგან სინთეზირებული ცილის დოზა.
მუტაცია შეიძლება მოხდეს სხეულის ნებისმიერ უჯრედში. თუ მუტაცია ხდება ჩანასახოვან უჯრედში, მას ე.წ ჩანასახოვანი(გენერაციული, ან გენერაციული). ასეთი მუტაციები არ ჩნდება იმ ორგანიზმში, რომელშიც ისინი გამოჩნდნენ, მაგრამ იწვევს მუტანტების გაჩენას შთამომავლობაში და მემკვიდრეობით მიიღება, ამიტომ ისინი მნიშვნელოვანია გენეტიკასა და ევოლუციისთვის. თუ მუტაცია ხდება რომელიმე სხვა უჯრედში, მას ე.წ სომატური. ასეთი მუტაცია შეიძლება გარკვეულწილად გამოვლინდეს იმ ორგანიზმში, რომელშიც ის წარმოიშვა, მაგალითად, გამოიწვიოს კიბოს სიმსივნეების წარმოქმნა. თუმცა, ასეთი მუტაცია არ არის მემკვიდრეობითი და არ მოქმედებს შთამომავლობაზე.
მუტაციებმა შეიძლება გავლენა მოახდინოს გენომის სხვადასხვა ზომის ნაწილებზე. გამოყოფა გენეტიკური, ქრომოსომულიდა გენომურიმუტაციები.
გენური მუტაციები
მუტაციებს, რომლებიც ხდება ერთ გენზე მცირე მასშტაბით, ეწოდება გენეტიკური, ან წერტილოვანი (წერტილი). ასეთი მუტაციები იწვევს ერთი ან რამდენიმე ნუკლეოტიდის ცვლილებას თანმიმდევრობაში. გენური მუტაციები მოიცავსჩანაცვლებებირაც იწვევს ერთი ნუკლეოტიდის მეორით ჩანაცვლებას,წაშლაიწვევს ერთ-ერთი ნუკლეოტიდის დაკარგვას,ჩანართები, რაც იწვევს დამატებით ნუკლეოტიდის დამატებით მიმდევრობას.

ბრინჯი. 23. გენის (წერტილოვანი) მუტაციები
ცილაზე მოქმედების მექანიზმის მიხედვით გენის მუტაციები იყოფა:სინონიმირაც (გენეტიკური კოდის გადაგვარების შედეგად) არ იწვევს ცილოვანი პროდუქტის ამინომჟავის შემადგენლობის ცვლილებას,უაზრო მუტაციებირაც იწვევს ერთი ამინომჟავის მეორით ჩანაცვლებას და შეუძლია გავლენა მოახდინოს სინთეზირებული ცილის სტრუქტურაზე, თუმცა ხშირად ისინი უმნიშვნელოა,უაზრო მუტაციები, რაც იწვევს კოდირების კოდონის ჩანაცვლებას გაჩერების კოდონით,მუტაციებისკენ მიმავალი შერწყმის დარღვევა:

ბრინჯი. 24. მუტაციის სქემები
ასევე, ცილაზე მოქმედების მექანიზმის მიხედვით, იზოლირებულია მუტაციები, რაც იწვევს ჩარჩოს ცვლა საკითხავიროგორიცაა ჩასმა და წაშლა. ასეთი მუტაციები, ისევე როგორც უაზრო მუტაციები, მიუხედავად იმისა, რომ ისინი წარმოიქმნება გენის ერთ წერტილში, ხშირად მოქმედებს ცილის მთელ სტრუქტურაზე, რამაც შეიძლება გამოიწვიოს მისი სტრუქტურის სრული ცვლილება. როდესაც ქრომოსომის სეგმენტი ბრუნავს 180 გრადუსით ბრინჯი. 28. ტრანსლოკაცია
როდესაც ქრომოსომის სეგმენტი ბრუნავს 180 გრადუსით ბრინჯი. 28. ტრანსლოკაცია
ბრინჯი. 29. ქრომოსომა დუბლირებამდე და მის შემდეგ
გენომური მუტაციები
ბოლოს და ბოლოს, გენომური მუტაციებიგავლენას ახდენს მთელ გენომზე, ანუ იცვლება ქრომოსომების რაოდენობა. განასხვავებენ პოლიპლოიდიას - უჯრედის პლოიდიის მატებას და ანევპლოიდიას, ანუ ქრომოსომების რაოდენობის ცვლილებას, მაგალითად, ტრიზომია (დამატებითი ჰომოლოგის არსებობა ერთ-ერთ ქრომოსომაში) და მონოსომია (არარსებობა. ჰომოლოგია ქრომოსომაში).
დნმ-თან დაკავშირებული ვიდეო
დნმ-ის რეპლიკაცია, რნმ-ის კოდირება, ცილების სინთეზი
(თუ ვიდეო არ არის ნაჩვენები, ის ხელმისაწვდომია
დნმ-ის ქიმიური სტრუქტურის მიხედვით ( Დეზოქსირიბონუკლეინის მჟავა) არის ბიოპოლიმერი, რომლის მონომერებია ნუკლეოტიდები. ანუ დნმ არის პოლინუკლეოტიდი. უფრო მეტიც, დნმ-ის მოლეკულა ჩვეულებრივ შედგება ორი ჯაჭვისგან, რომლებიც ერთმანეთთან შედარებით ხვეულია ხვეული ხაზის გასწვრივ (ხშირად უწოდებენ "სპირალურ გრეხილს") და ერთმანეთთან დაკავშირებულია წყალბადის ბმებით.
ჯაჭვები შეიძლება გადატრიალდეს როგორც მარცხნივ, ასევე მარჯვნივ (ყველაზე ხშირად).
ზოგიერთ ვირუსს აქვს ერთჯაჭვიანი დნმ.
თითოეული დნმ ნუკლეოტიდი შედგება 1) აზოტოვანი ბაზისგან, 2) დეზოქსირიბოზისგან, 3) ფოსფორის მჟავას ნარჩენებისგან.
ორმაგი მარჯვენა დნმ-ის სპირალი
დნმ შეიცავს შემდეგს: ადენინი, გუანინი, თიმინიდა ციტოზინი. ადენინი და გუანინი არიან პურინები, ხოლო თიმინი და ციტოზინი - მდე პირიმიდინები. ზოგჯერ დნმ შეიცავს ურაცილს, რომელიც ჩვეულებრივ დამახასიათებელია რნმ-სთვის, სადაც ის ცვლის თიმინს.
დნმ-ის მოლეკულის ერთი ჯაჭვის აზოტოვანი ფუძეები დაკავშირებულია მეორის აზოტოვან ფუძეებთან მკაცრად კომპლემენტარობის პრინციპის მიხედვით: ადენინი მხოლოდ თიმინთან (ისინი ქმნიან ორ წყალბადურ კავშირს ერთმანეთთან), ხოლო გუანინი მხოლოდ ციტოზინთან (სამი ბმული).
თავად ნუკლეოტიდში აზოტოვანი ბაზა დაკავშირებულია ციკლური ფორმის პირველ ნახშირბადის ატომთან. დეზოქსირიბოზა, რომელიც არის პენტოზა (ნახშირწყალი ხუთი ნახშირბადის ატომით). ბმა არის კოვალენტური, გლიკოზიდური (C-N). რიბოზასგან განსხვავებით, დეზოქსირიბოზას აკლია მისი ერთ-ერთი ჰიდროქსილის ჯგუფი. დეზოქსირიბოზის რგოლი იქმნება ოთხი ნახშირბადის ატომით და ერთი ჟანგბადის ატომით. ნახშირბადის მეხუთე ატომი რგოლს გარეთაა და ჟანგბადის ატომის მეშვეობით უკავშირდება ფოსფორის მჟავას ნარჩენს. ასევე, მესამე ნახშირბადის ატომის ჟანგბადის ატომის მეშვეობით, მიმაგრებულია მეზობელი ნუკლეოტიდის ფოსფორმჟავას ნარჩენი.
ამრიგად, დნმ-ის ერთ ჯაჭვში მიმდებარე ნუკლეოტიდები ერთმანეთთან არის დაკავშირებული კოვალენტური ბმებიდეზოქსირიბოზასა და ფოსფორის მჟავას (ფოსფოდიესტერული ბმა) შორის. იქმნება ფოსფატ-დეოქსირიბოზის ხერხემალი. მასზე პერპენდიკულურად, დნმ-ის სხვა ჯაჭვისკენ მიმართულია აზოტოვანი ფუძეები, რომლებიც წყალბადის ბმებით უერთდებიან მეორე ჯაჭვის ფუძეებს.
დნმ-ის სტრუქტურა ისეთია, რომ წყალბადის ობლიგაციებით დაკავშირებული ჯაჭვების ხერხემალი მიმართულია სხვადასხვა მიმართულებით (ისინი ამბობენ „მრავალმიმართულები“, „ანტიპარალელური“). იმ მხარეს, სადაც ერთი მთავრდება ფოსფორის მჟავით, რომელიც დაკავშირებულია დეზოქსირიბოზის მეხუთე ნახშირბადის ატომთან, მეორე მთავრდება "თავისუფალი" მესამე ნახშირბადის ატომით. ანუ ერთი ჯაჭვის ჩონჩხი თავდაყირა დგას, თითქოსდა მეორესთან შედარებით. ამრიგად, დნმ-ის ჯაჭვების სტრუქტურაში გამოიყოფა 5 „ბოლო და 3“ ბოლო.
დნმ-ის რეპლიკაციისას (გაორმაგება), ახალი ჯაჭვების სინთეზი ყოველთვის მიმდინარეობს მათი მე-5 ბოლოდან მესამემდე, ვინაიდან ახალი ნუკლეოტიდების მიმაგრება შესაძლებელია მხოლოდ თავისუფალ მესამე ბოლოზე.
საბოლოო ჯამში (ირიბად რნმ-ის საშუალებით), ყოველი სამი თანმიმდევრული ნუკლეოტიდი დნმ-ის ჯაჭვში კოდირებს ცილის ერთ ამინომჟავას.
დნმ-ის მოლეკულის სტრუქტურის აღმოჩენა მოხდა 1953 წელს ფ. კრიკისა და დ. უოტსონის მუშაობის წყალობით (რასაც ასევე შეუწყო ხელი სხვა მეცნიერების ადრეულმა მუშაობამ). თუმცა როგორ ქიმიური ნივთიერებადნმ ცნობილია მე-19 საუკუნიდან. 1940-იან წლებში გაირკვა, რომ დნმ არის გენეტიკური ინფორმაციის მატარებელი.
ორმაგი სპირალი განიხილება დნმ-ის მოლეკულის მეორად სტრუქტურად. ევკარიოტულ უჯრედებში დნმ-ის დიდი რაოდენობა განლაგებულია ქრომოსომებში, სადაც ის ასოცირდება ცილებთან და სხვა ნივთიერებებთან და ასევე გადის უფრო მჭიდრო შეფუთვას.
Დეზოქსირიბონუკლეინის მჟავა ან დნმ არის გენეტიკური ინფორმაციის მატარებელი. უჯრედებში დნმ-ის უმეტესი ნაწილი მდებარეობს ბირთვში. ეს არის ქრომოსომების მთავარი კომპონენტი. ევკარიოტებში დნმ ასევე გვხვდება მიტოქონდრიებში და პლასტიდებში. დნმ შედგება ერთმანეთზე კოვალენტურად დაკავშირებული მონონუკლეოტიდებისგან, რომლებიც წარმოადგენს გრძელ განშტოებულ პოლიმერს. მონონუკლეოტიდები, რომლებიც ქმნიან დნმ-ს, შედგება დეზოქსირიბოზისგან, 4 აზოტოვანი ბაზიდან ერთ-ერთი (ადენინი, გუანინი, ციტოზინი და თიმინი) და ფოსფორმჟავას ნარჩენი. ამ მონონუკლეოტიდების რაოდენობა ძალიან დიდია. მაგალითად, პროკარიოტულ უჯრედებში, რომლებიც შეიცავს ერთ ქრომოსომას, დნმ არის ერთი მაკრომოლეკულა, რომლის მოლეკულური წონა 2 x 10 9-ზე მეტია.
დნმ-ის ერთი ჯაჭვის მონონუკლეოტიდები ფორმირების გამო სერიულად არის დაკავშირებული ერთმანეთთან კოვალენტური ფოსფოდიესტერის ბმები ერთი მონონუკლეოტიდის დეზოქსირიბოზა OH ჯგუფსა და მეორის ფოსფორმჟავას ნარჩენებს შორის. დნმ-ის ერთი ჯაჭვის ჩამოყალიბებული ხერხემლის ერთ მხარეს არის აზოტოვანი ფუძეები. ისინი შეიძლება შევადაროთ ერთ ძაფზე დადებულ ოთხ სხვადასხვა მძივს, რადგან. ისინი, როგორც იქნა, შაქრის ფოსფატის ჯაჭვზეა მიბმული.
ჩნდება კითხვა, როგორ შეუძლია ამ გრძელ პოლინუკლეოტიდურ ჯაჭვს უჯრედის ან თუნდაც მთელი ორგანიზმის განვითარების პროგრამის კოდირება? ამ კითხვაზე პასუხის მიღება შესაძლებელია იმის გაგებით, თუ როგორ იქმნება დნმ-ის სივრცითი სტრუქტურა. ამ მოლეკულის სტრუქტურა გაშიფრეს და აღწერეს ჯ. უოტსონმა და ფ. კრიკმა 1953 წელს.
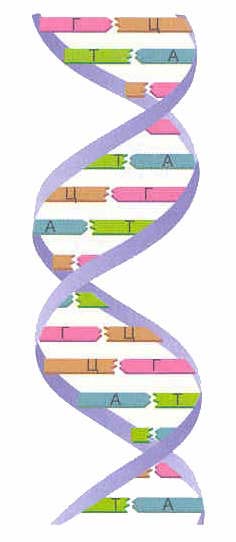
დნმ-ის მოლეკულები არის ორი ჯაჭვი, რომლებიც ერთმანეთის პარალელურია და წარმოიქმნება მარჯვენა სპირალი
. ამ სპირალის სიგანე დაახლოებით 2 ნმ-ია, მაგრამ მისი სიგრძე ასობით ათასი ნანომეტრს აღწევს. უოტსონმა და კრიკმა შემოგვთავაზეს დნმ-ის მოდელი, რომლის მიხედვითაც დნმ-ის ყველა ბაზა განლაგებულია სპირალის შიგნით, შაქარ-ფოსფატის ხერხემალი გარეთაა. ამრიგად, ერთი ჯაჭვის ფუძეები რაც შეიძლება ახლოსაა მეორის ფუძეებთან,
ამიტომ წყალბადის ბმები იქმნება მათ შორის. დნმ-ის სპირალის სტრუქტურა ისეთია, რომ პოლინუკლეოტიდური ჯაჭვები, რომლებიც მას ქმნიან, შეიძლება განცალკევდეს მხოლოდ მისი გახსნის შემდეგ.
დნმ-ის ორი ჯაჭვის მაქსიმალური სიახლოვის გამო, მისი შემადგენლობა შეიცავს ერთი ტიპის აზოტოვან ფუძეებს (ადენინი და გუანინი) და სხვა ტიპის აზოტოვან ფუძეებს (თიმინი და ციტოზინი), ანუ მოქმედებს ფორმულა: A+G=T+C. ეს გამოწვეულია აზოტოვანი ფუძეების ზომით, კერძოდ, სტრუქტურების სიგრძე, რომლებიც წარმოიქმნება წყალბადის კავშირის გამო, ადენინ-თიმინსა და გუანინ-ციტოზინის წყვილებს შორის არის დაახლოებით 1,1 ნმ. ამ წყვილების საერთო ზომები შეესაბამება დნმ-ის სპირალის შიდა ნაწილის ზომებს. სპირალის ჩამოყალიბება წყვილი C-Tძალიან პატარა იქნებოდა წყვილი A-Gპირიქით, ძალიან დიდია. ანუ დნმ-ის პირველი ჯაჭვის აზოტოვანი ფუძე განსაზღვრავს ფუძეს, რომელიც მდებარეობს დნმ-ის მეორე ჯაჭვის იმავე ადგილას. ნუკლეოტიდების მკაცრი კორესპონდენცია, რომელიც მდებარეობს დნმ-ის მოლეკულაში ერთმანეთის პარალელურად დაწყვილებულ ჯაჭვებში ე.წ. კომპლემენტარულობა (სურვილისამებრ). ზუსტი გამრავლება ან რეპლიკაცია გენეტიკური ინფორმაცია შესაძლებელია სწორედ დნმ-ის მოლეკულის ამ მახასიათებლის გამო.
დნმ-ში ბიოლოგიური ინფორმაცია ისეა ჩაწერილი, რომ მისი ზუსტად კოპირება და შთამომავლობის უჯრედებში გადაცემა შესაძლებელია. უჯრედის გაყოფამდე, რეპლიკაცია (თვითგაორმაგება ) დნმ. ვინაიდან თითოეული ჯაჭვი შეიცავს პარტნიორი ჯაჭვის დამატებით ნუკლეოტიდურ თანმიმდევრობას, ისინი რეალურად ატარებენ იგივე გენეტიკურ ინფორმაციას. თუ თქვენ გამოყოფთ ძაფებს და გამოიყენებთ თითოეულ მათგანს, როგორც შაბლონს (მატრიცას) მეორე ჯაჭვის ასაგებად, თქვენ მიიღებთ დნმ-ის ორ ახალ იდენტურ ჯაჭვს. ასე ხდება დნმ-ის დუბლირება უჯრედში.


