ДНҚ неден тұрады. Нуклеотид - бұл не? ДНҚ тізбегіндегі нуклеотидтердің құрамы, құрылымы, саны және реттілігі ДНҚ қос спиралі иондармен тұрақтандырылады.
Нуклеин қышқылдары - мононуклеотидтерден тұратын, бір-бірімен 3",5" - фосфодиэфирлік байланыстар арқылы полимер тізбегінде байланысқан және белгілі бір жолмен жасушаларға оралған макромолекулярлық заттар.
Нуклеин қышқылдары екі түрлі биополимерлер болып табылады: рибонуклеин қышқылы (РНҚ) және дезоксирибонуклеин қышқылы (ДНҚ). Әрбір биополимер көмірсулар қалдығы (рибоза, дезоксирибоза) және азотты негіздердің бірінен (урацил, тимин) ерекшеленетін нуклеотидтерден тұрады. Осыған сәйкес нуклеин қышқылдары өз атауын алды.
Дезоксирибонуклеин қышқылының құрылымы
Нуклеин қышқылдарының біріншілік, екіншілік және үшінші реттік құрылымдары болады.
ДНҚ-ның біріншілік құрылымы
ДНҚ-ның бастапқы құрылымы мононуклеотидтер 3", 5" фосфодиэфирлік байланыстармен байланысқан сызықты полинуклеотидтік тізбек болып табылады. Жасушадағы нуклеин қышқылы тізбегін жинақтаудың бастапқы материалы нуклеозид 5'-трифосфат болып табылады, ол фосфор қышқылының β және γ қалдықтарын жою нәтижесінде басқа нуклеозидтің 3'-көміртек атомын бекітуге қабілетті. . Осылайша, бір дезоксирибозаның 3" көміртегі атомы басқа дезоксирибозаның 5" көміртегі атомымен бір фосфор қышқылы қалдығы арқылы ковалентті байланысады және нуклеин қышқылының сызықты полинуклеотидтік тізбегін құрайды. Осыдан аталды: 3", 5"-фосфодиэфирлік байланыстар. Азотты негіздер бір тізбектің нуклеотидтерінің қосылуына қатыспайды (1. сурет).
Бір нуклеотидтің фосфор қышқылының қалдығы мен екіншісінің көмірсуы арасындағы мұндай байланыс полинуклеотид молекуласының пентозофосфатты діңгегінің түзілуіне әкеледі, оған бүйірден бірінен соң бірі азотты негіздер қосылады. Нуклеин қышқылы молекулаларының тізбегіндегі олардың реттілігі әртүрлі организмдердің жасушалары үшін қатаң түрде тән, яғни. белгілі бір сипатқа ие (Чаргафф ережесі).
Ұзындығы тізбекке кіретін нуклеотидтердің санына байланысты болатын сызықты ДНҚ тізбегінің екі ұшы бар: біреуі 3 «соңы» деп аталады және бос гидроксилден тұрады, ал екіншісінде, 5» ұшында фосфор қышқылы бар. қалдық. Схема полярлы және 5"->3" және 3"->5" болуы мүмкін. Ерекшелік - дөңгелек ДНҚ.
ДНҚ-ның генетикалық «мәтіні» кодтық «сөздерден» – кодондар деп аталатын нуклеотидтердің үштіктерінен тұрады. РНҚ-ның барлық түрлерінің бастапқы құрылымы туралы ақпаратты қамтитын ДНҚ сегменттері құрылымдық гендер деп аталады.
Полинуклеодиттік ДНҚ тізбегі үлкен мөлшерге жетеді, сондықтан олар жасушада белгілі бір түрде оралады.
ДНҚ құрамын зерттей отырып, Чаргафф (1949) жеке ДНҚ негіздерінің мазмұнына қатысты маңызды заңдылықтарды белгіледі. Олар ДНҚ-ның қайталама құрылымын ашуға көмектесті. Бұл үлгілер Чаргафф ережелері деп аталады. Чаргафф ережелері
Бұл ережелер ДНҚ құру кезінде пурин мен пиримидиндік негіздер үшін емес, әсіресе аденинмен тимин және гуанинмен цитозин үшін өте қатаң сәйкестікті (жұптастыру) сақтау керектігін айтады. Осы ережелерге сүйене отырып, басқа нәрселермен қатар, 1953 жылы Уотсон мен Крик қос спираль деп аталатын ДНҚ-ның қайталама құрылымының моделін ұсынды (сурет). |

ДНҚ-ның екіншілік құрылымы
ДНҚ-ның екінші реттік құрылымы – қос спираль, оның моделін 1953 жылы Д.Уотсон мен Ф.Крик ұсынған.
ДНҚ моделін құрудың алғы шарттары
Бастапқы талдаулардың нәтижесінде кез келген шыққан ДНҚ-да барлық төрт нуклеотид бірдей молярлық мөлшерде болады деген идея болды. Алайда 1940 жылдары Э.Чаргафф және оның әріптестері әртүрлі организмдерден бөлініп алынған ДНҚ-ны талдау нәтижесінде олардың құрамында азотты негіздердің әртүрлі сандық қатынаста болатынын анық көрсетті. Чаргафф бір түрдегі организмдердің барлық жасушаларының ДНҚ-сы үшін бұл арақатынастар бірдей болғанымен, әртүрлі түрдегі ДНҚ белгілі бір нуклеотидтердің құрамында айтарлықтай ерекшеленетінін анықтады. Бұл азотты негіздердің арақатынасындағы айырмашылықтар кейбір биологиялық кодпен байланысты болуы мүмкін деген болжам жасады. Әртүрлі ДНҚ үлгілеріндегі жеке пурин және пиримидин негіздерінің арақатынасы бірдей болмаса да, талдау нәтижелерін салыстыру кезінде белгілі бір заңдылық анықталды: барлық үлгілерде пуриндердің жалпы мөлшері пиримидиндердің жалпы санына тең болды ( A + G = T + C), аденин мөлшері тимин мөлшеріне (А = Т), ал гуаниннің мөлшері - цитозиннің мөлшеріне (G = C) тең болды. Сүтқоректілердің жасушаларынан бөлінген ДНҚ әдетте аденин мен тиминге бай және гуанин мен цитозинге салыстырмалы түрде кедей болды, ал бактериялардың ДНҚ гуанин мен цитозинге бай және аденин мен тиминге салыстырмалы түрде кедей болды. Бұл деректер фактілік материалдың маңызды бөлігін құрады, оның негізінде кейінірек Уотсон-Крик ДНҚ құрылымының моделі құрастырылды.
ДНҚ-ның мүмкін болатын құрылымының тағы бір маңызды жанама көрсеткіші Л.Полингтің белок молекулаларының құрылымы туралы мәліметтері болды. Полинг ақуыз молекуласында аминқышқылдары тізбегінің бірнеше түрлі тұрақты конфигурацияларының болуы мүмкін екенін көрсетті. Пептидтік тізбектің жалпы конфигурацияларының бірі – α-спираль – қалыпты спиральдық құрылым. Мұндай құрылыммен тізбектің іргелес бұрылыстарында орналасқан аминқышқылдары арасында сутектік байланыстардың пайда болуы мүмкін. Полинг 1950 жылы полипептидтік тізбектің α-спиральдық конфигурациясын сипаттады және ДНҚ молекулаларының сутегі байланыстарымен бекітілген спиральдық құрылымы болуы мүмкін деген болжам жасады.
Дегенмен, ДНҚ молекуласының құрылымы туралы ең құнды ақпаратты рентгендік дифракциялық талдау нәтижелері берді. Рентген сәулелері ДНҚ кристалы арқылы өтіп, дифракцияға ұшырайды, яғни белгілі бір бағытта ауытқиды. Сәулелердің ауытқу дәрежесі мен сипаты молекулалардың өздерінің құрылымына байланысты. Рентген сәулелерінің дифракция үлгісі (3-сурет) тәжірибелі көзге зерттелетін заттың молекулаларының құрылымына қатысты бірқатар жанама көрсеткіштерді береді. ДНҚ-ның рентгендік дифракция заңдылықтарын талдау азотты негіздер (жалпақ пішіні бар) пластиналар дестесі тәрізді қабаттасады деген қорытындыға әкелді. Рентгендік үлгілер кристалдық ДНҚ құрылымындағы үш негізгі кезеңді анықтауға мүмкіндік берді: 0,34, 2 және 3,4 нм.

Уотсон-Крик ДНҚ моделі
Чаргаффтың аналитикалық деректерінен, Уилкинстің рентгендік сәулелерінен және молекуладағы атомдар арасындағы нақты қашықтық, берілген атомның байланыстары арасындағы бұрыштар және атомдардың өлшемдері туралы ақпарат беретін химиктер Уотсон мен Крик белгілі бір масштабта ДНҚ молекуласының жеке құрамдас бөліктерінің физикалық модельдерін құру және алынған жүйе әртүрлі эксперименттік деректерге сәйкес келетіндей етіп оларды бір-біріне «реттеу». [көрсету] .
Бұдан бұрын да ДНҚ тізбегіндегі көрші нуклеотидтер бір нуклеотидтің дезоксирибозасының 5'-көміртегі атомын келесі нуклеотидтің дезоксирибозасының 3'-көміртекті атомымен байланыстыратын фосфодиэфирлі көпірлер арқылы байланысатыны белгілі болды. Уотсон мен Крик 0,34 нм период ДНҚ тізбегіндегі кезекті нуклеотидтер арасындағы қашықтыққа сәйкес келетініне еш күмәнданбады. Әрі қарай, 2 нм периоды тізбектің қалыңдығына сәйкес келеді деп болжауға болады. Ал 3,4 нм периоды қандай нақты құрылымға сәйкес келетінін түсіндіру үшін Уотсон мен Крик, сондай-ақ бұрын Полинг тізбектің спираль түрінде бұралғанын (немесе дәлірек айтқанда, спираль құрайды, өйткені спираль бұл сөздің қатаң мағынасында бұрылыстар кеңістікте цилиндрлік бетті емес, конусты құраған кезде алынады). Сонда 3,4 нм периоды осы спиральдың кезекті бұрылыстары арасындағы қашықтыққа сәйкес келеді. Мұндай спираль өте тығыз немесе біршама созылған болуы мүмкін, яғни оның бұрылыстары тегіс немесе тік болуы мүмкін. 3,4 нм периоды дәйекті нуклеотидтер арасындағы қашықтықты (0,34 нм) дәл 10 есе көп болғандықтан, спиральдың әрбір толық айналымы 10 нуклеотидтен тұратыны анық. Осы деректерден Уотсон мен Крик диаметрі 2 нм, бұрылыстар арасындағы қашықтық 3,4 нм болатын спиральға бұралған полинуклеотидтік тізбектің тығыздығын есептей алды. Мұндай жіптің тығыздығы бұрыннан белгілі болған ДНҚ-ның нақты тығыздығының жартысы болатыны белгілі болды. Мен ДНҚ молекуласы екі тізбектен тұрады деп болжауға тура келді - бұл нуклеотидтердің қос спиралі.
Келесі міндет, әрине, қос спиралді құрайтын екі жіптің арасындағы кеңістіктік қатынасты түсіндіру болды. Уотсон мен Крик өздерінің физикалық моделінде бірнеше тізбекті орналастыруды сынап көргенде, барлық қол жетімді деректер үшін ең жақсы сәйкестік екі полинуклеотидтік спиралдың қарама-қарсы бағытта жүретінін анықтады; бұл жағдайда қант пен фосфат қалдықтарынан тұратын тізбектер қос спиралдың бетін құрайды, ал оның ішінде пуриндер мен пиримидиндер орналасады. Бір-біріне қарама-қарсы орналасқан, екі тізбекке жататын негіздер сутектік байланыс арқылы жұппен байланысқан; дәл осы сутектік байланыстар тізбектерді біріктіреді, осылайша молекуланың жалпы конфигурациясын бекітеді.
ДНҚ қос спиралын баспалдақтар көлденең қалатын бұрандалы арқан баспалдақтары ретінде қарастыруға болады. Содан кейін екі бойлық арқан қант пен фосфат қалдықтарының тізбегіне сәйкес келеді, ал көлденең жолақтар сутегі байланыстарымен қосылған азотты негіздердің жұптарына сәйкес болады.
Ықтимал модельдерді одан әрі зерттеу нәтижесінде Уотсон мен Крик әрбір «крест» бір пуриннен және бір пиримидиннен тұруы керек деген қорытындыға келді; 2 нм периодта (қос спиральдың диаметріне сәйкес) екі пурин үшін орын жеткіліксіз болады және екі пиримидин дұрыс сутектік байланыстарды құру үшін бір-біріне жақын бола алмайды. Егжей-тегжейлі модельді тереңдетіп зерттеу дұрыс өлшемдегі комбинацияны құрайтын аденин мен цитозинді әлі де олардың арасында сутегі байланыстары пайда болатындай етіп орналастыра алмайтынын көрсетті. Осыған ұқсас есептер гуанин-тимин комбинациясын алып тастауға мәжбүр етті, ал аденин-тимин және гуанин-цитозин комбинациялары әбден қолайлы деп табылды. Сутектік байланыстардың табиғаты аденин тиминмен, гуанин цитозинмен жұптасады. Нақты негіздердің жұптасуының бұл тұжырымдамасы «Шаргафф ережесін» түсіндіруге мүмкіндік берді, оған сәйкес кез келген ДНҚ молекуласында аденин мөлшері әрқашан тиминнің құрамына, ал гуанин мөлшері әрқашан цитозиннің мөлшеріне тең болады. . Аденин мен тимин арасында екі, гуанин мен цитозин арасында үш сутектік байланыс түзіледі. Бір тізбектегі әрбір аденинге қарсы сутектік байланыстың түзілуіндегі осы ерекшелікке байланысты, екінші тізбекте тимин болады; дәл осылай әрбір гуанинге қарсы тек цитозинді қоюға болады. Осылайша, тізбектер бірін-бірі толықтырады, яғни бір тізбектегі нуклеотидтердің тізбегі олардың екінші тізбектегі реттілігін бірегей түрде анықтайды. Екі тізбек қарама-қарсы бағытта жүреді және олардың фосфаттық соңғы топтары қос спиралдың қарама-қарсы ұштарында болады.
Өздерінің зерттеулерінің нәтижесінде 1953 жылы Уотсон мен Крик ДНҚ молекуласының құрылымының моделін ұсынды (3-сурет), ол қазіргі уақытқа дейін өзекті болып қала береді. Модельге сәйкес ДНҚ молекуласы екі комплементарлы полинуклеотидтік тізбектен тұрады. Әрбір ДНҚ тізбегі бірнеше ондаған мың нуклеотидтерден тұратын полинуклеотид болып табылады. Онда көрші нуклеотидтер күшті коваленттік байланыс арқылы фосфор қышқылының қалдығы мен дезоксирибозаның қосылуы есебінен қалыпты пентозофосфатты діңгек түзеді. Бір полинуклеотидтік тізбектің азотты негіздері екіншісінің азотты негіздеріне қарсы қатаң белгіленген тәртіпте орналасады. Полинуклеотидтік тізбектегі азотты негіздердің кезектесуі дұрыс емес.
ДНҚ тізбегіндегі азотты негіздердің орналасуы комплементарлы (грекше «комплемент» - қосу), яғни. аденинге (А) қарсы әрқашан тимин (Т), ал гуанинге (G) қарсы - тек цитозин (С). Бұл A және T, сондай-ақ G және C бір-біріне қатаң сәйкес келетіндігімен түсіндіріледі, яғни. бірін-бірі толықтырады. Бұл сәйкестік пурин мен пиримидин жұбында сутектік байланыстардың түзілуіне мүмкіндік беретін негіздердің химиялық құрылымымен берілген. А мен Т арасында екі байланыс, G мен С арасында үш байланыс бар. Бұл байланыстар ДНҚ молекуласының кеңістікте ішінара тұрақтануын қамтамасыз етеді. Қос спиральдың тұрақтылығы A=T байланыстарына қарағанда тұрақтырақ G≡C байланыстарының санына тура пропорционал.
ДНҚ-ның бір тізбегіндегі нуклеотидтердің белгілі тізбегі комплементарлық принцип бойынша басқа тізбектің нуклеотидтерін орнатуға мүмкіндік береді.
Сонымен қатар, хош иісті құрылымға ие азотты негіздер су ерітіндісінде бірінің үстіне бірі орналасып, тиындар дестесін құрайтыны анықталды. Бұл стектерді қалыптастыру процесі органикалық молекулаларжинақтау деп аталады. Қарастырылып отырған Уотсон-Крик моделінің ДНҚ молекуласының полинуклеотидтік тізбектері ұқсас физика-химиялық күйге ие, олардың азотты негіздері тиындар дестелері түрінде орналасады, олардың жазықтықтары арасында ван-дер-Ваальс әрекеттесулері (стектік әрекеттесулер) жүреді.
Ван-дер-Ваальс күштерінің әсерінен (тігінен) полинуклеотидтік тізбектегі комплементарлы негіздер арасындағы сутектік байланыстар және базалық жазықтықтар арасындағы стектік өзара әрекеттесу ДНҚ молекуласын кеңістікте қосымша тұрақтандыруды қамтамасыз етеді.
Екі тізбектің қант-фосфатты омыртқалары сыртқа, ал негіздері ішке, бір-біріне қарай бұрылған. ДНҚ-дағы жіптердің бағыты антипараллельді (бірінің бағыты 5"->3", екіншісі - 3"->5", яғни бір тізбектің 3"-ұшы 5"-ұшына қарама-қарсы орналасқан. екіншісінен.). Тізбектер ортақ осі бар оң жақ спиральдарды құрайды. Спиральдың бір айналымы 10 нуклеотид, бұрылыс өлшемі 3,4 нм, әрбір нуклеотидтің биіктігі 0,34 нм, спираль диаметрі 2,0 нм. Бір жіптің екіншісінің айналасында айналуы нәтижесінде ДНҚ қос спиралінде үлкен ойық (диаметрі шамамен 20 Å) және кіші ойық (шамамен 12 Å) пайда болады. Уотсон-Крик қос спиралының бұл түрі кейінірек В-формасы деп аталды. Жасушаларда ДНҚ әдетте В түрінде болады, ол ең тұрақты.
ДНҚ функциялары
Ұсынылған модель дезоксирибонуклеин қышқылының көптеген биологиялық қасиеттерін, соның ішінде генетикалық ақпаратты сақтауды және гендердің әртүрлілігін, 4 нуклеотидтердің бірізді қосылыстарының алуан түрлілігімен және генетикалық кодтың болуы фактісімен, гендік кодтың болуын түсіндірді. репликация процесімен қамтамасыз етілген генетикалық ақпаратты өздігінен көбейту және беру және белоктар түрінде генетикалық ақпаратты жүзеге асыру, сонымен қатар ферменттік ақуыздардың көмегімен түзілетін кез келген басқа қосылыстар.
ДНҚ-ның негізгі қызметтері.
- ДНҚ – генетикалық ақпараттың тасымалдаушысы, ол генетикалық кодтың болуымен қамтамасыз етіледі.
- Жасушалар мен ағзалардың ұрпақтарында көбею және берілетін генетикалық ақпарат. Бұл функция репликация процесі арқылы қамтамасыз етіледі.
- Генетикалық ақпаратты белоктар түрінде жүзеге асыру, сонымен қатар ферменттік ақуыздардың көмегімен түзілетін кез келген басқа қосылыстар. Бұл функция транскрипция және аударма процестерімен қамтамасыз етіледі.

Қос тізбекті ДНҚ-ның ұйымдасу формалары
ДНҚ қос спиральдың бірнеше түрін құра алады (4-сурет). Қазіргі уақытта алты пішін белгілі (А-дан Е және Z-формаға дейін).
Розалинд Франклин белгілеген ДНҚ-ның құрылымдық формалары нуклеин қышқылы молекуласының сумен қанығуына байланысты. Рентгендік дифракциялық талдауды қолдану арқылы ДНҚ талшықтарын зерттеуде рентгендік дифракцияның заңдылығы қандай салыстырмалы ылғалдылықта, осы талшықтың сумен қанығу дәрежесі қандай дәрежеде тәжірибе жүргізілетініне түбегейлі тәуелді болатыны көрсетілді. Егер талшық сумен жеткілікті қаныққан болса, онда бір рентгенограмма алынды. Кептіру кезінде ылғалдылығы жоғары талшықтың рентгендік үлгісінен мүлде басқа рентгендік сурет пайда болды.
Ылғалдылығы жоғары ДНҚ молекуласы В-тәрізді деп аталады. Физиологиялық жағдайларда (тұздың төмен концентрациясы, гидратацияның жоғары дәрежесі) ДНҚ-ның құрылымдық түрі В-формасы (екі тізбекті ДНҚ-ның негізгі формасы Уотсон-Крик моделі) болып табылады. Мұндай молекуланың спираль қадамы 3,4 нм. Бір айналымда «тиындардың» бұралған дестелері – азотты негіздер түріндегі 10 толықтырушы жұп бар. Стектерді бір-біріне қарама-қарсы екі «тиын» арасындағы сутектік байланыстар арқылы ұстайды және оң жақ бұрандаға бұралған фосфодиэфирлік магистральдың екі лентасы бар «орамалды». Азотты негіздердің жазықтықтары спираль осіне перпендикуляр. Көршілес толықтауыш жұптар бір-біріне қатысты 36°-қа бұрылады. Спираль диаметрі 20Å, пурин нуклеотиді 12Å және пиримидин нуклеотиді 8Å алады.
Ылғалдылығы төмен ДНҚ молекуласы А-формасы деп аталады. А-формасы аз жоғары гидратация жағдайында және Na+ немесе K+ иондарының көп мөлшерімен түзіледі. Бұл кеңірек оң жақ конформацияда бір айналымда 11 негізгі жұп бар. Азотты негіздердің жазықтықтары спираль осіне күштірек еңіске ие, олар спиралдың осіне нормальдан 20° ауытқиды. Бұл диаметрі 5 Å болатын ішкі қуыстың болуын білдіреді. Көршілес нуклеотидтер арасындағы қашықтық 0,23 нм, катушканың ұзындығы 2,5 нм, спираль диаметрі 2,3 нм.
Бастапқыда ДНҚ-ның А-пішіні онша маңызды емес деп есептелді. Алайда кейінірек ДНҚ-ның А-формасы сияқты В-формасының биологиялық маңызы зор екені белгілі болды. Шаблон-тұқым кешеніндегі РНҚ-ДНҚ спиральында А-формасы бар, сонымен қатар РНҚ-РНҚ спираль және РНҚ шаш қыстырғыш құрылымдары бар (рибозаның 2'-гидроксил тобы РНҚ молекулаларының В-формасын құруына мүмкіндік бермейді) . ДНҚ-ның А-формасы спораларда кездеседі. ДНҚ-ның А-формасы В-формасына қарағанда УК сәулелеріне төзімділігі 10 есе жоғары екені анықталды.
А-формасы және В-формасы деп аталады канондық формаларДНҚ.
C-E пішіндерісонымен қатар оң қолдар, олардың түзілуін тек арнайы эксперименттерде байқауға болады, және, шамасы, олар in vivo жоқ. ДНҚ-ның С-формасы В-ДНҚ-ға ұқсас құрылымға ие. Бір айналымдағы негізгі жұптардың саны 9,33, ал спираль ұзындығы 3,1 нм. Негізгі жұптар оське перпендикулярлық жағдайға қатысты 8 градус бұрышта көлбеу. Ойықтар көлемі бойынша В-ДНҚ ойықтарына жақын. Бұл жағдайда негізгі ойық біршама кішірек, ал кіші ойық тереңірек болады. Табиғи және синтетикалық ДНҚ полинуклеотидтері С пішініне өте алады.
| Кесте 1. ДНҚ құрылымдарының кейбір түрлерінің сипаттамасы | |||
| Спираль түрі | А | Б | З |
| Спиральды қадам | 0,32 нм | 3,38 нм | 4,46 нм |
| Спиральды бұралу | Дұрыс | Дұрыс | Сол |
| Бір айналымдағы негізгі жұптардың саны | 11 | 10 | 12 |
| Негізгі жазықтықтар арасындағы қашықтық | 0,256 нм | 0,338 нм | 0,371 нм |
| Гликозидтік байланыстың конформациясы | қарсы | қарсы | қарсы С син-Г |
| Фураноза сақинасының конформациясы | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-C |
| Ойықтың ені, кіші/үлкен | 1,11/0,22 нм | 0,57/1,17 нм | 0,2/0,88 нм |
| Ойық тереңдігі, шағын/үлкен | 0,26/1,30 нм | 0,82/0,85 нм | 1,38/0,37 нм |
| Спираль диаметрі | 2,3 нм | 2,0 нм | 1,8 нм |
ДНҚ-ның құрылымдық элементтері
(канондық емес ДНҚ құрылымдары)
ДНҚ-ның құрылымдық элементтеріне кейбір арнайы тізбектермен шектелген әдеттен тыс құрылымдар жатады:
|

ДНҚ-ның Z-формасы 1979 жылы d(CG)3 - гексануклеотидін зерттеу барысында ашылды. Оны MIT профессоры Александр Рич және оның қызметкерлері ашты. Z-формасы ДНҚ-ның ең маңызды құрылымдық элементтерінің біріне айналды, себебі оның түзілуі пуриндер пиримидиндермен (мысалы, 5'-HCHCHC-3') кезектесетін ДНҚ аймақтарында немесе 5' қайталануларында байқалды. -CHCHCH-3' құрамында метилденген цитозин бар. Z-ДНҚ түзілуі мен тұрақтануының маңызды шарты ондағы пурин нуклеотидтерінің син-конформацияда, антиконформацияда пиримидиндік негіздермен алмасып тұруы болды.
Табиғи ДНҚ молекулалары, егер оларда (CG)n сияқты тізбектер болмаса, көбінесе дұрыс В түрінде болады. Алайда, егер мұндай тізбектер ДНҚ-ның бөлігі болса, онда бұл аймақтар ерітіндінің иондық күші немесе фосфодиэфирлік магистральдағы теріс зарядты бейтараптандыратын катиондар Z-формасына ауыса алады, ал тізбектегі басқа ДНҚ аймақтары қалады. классикалық В-пішінде. Мұндай ауысу мүмкіндігі ДНҚ қос спиралындағы екі жіптің динамикалық күйде болатынын және бір-біріне қатысты босап, оң жақтан солға және керісінше өтетінін көрсетеді. ДНҚ құрылымының конформациялық түрленуіне мүмкіндік беретін бұл лабильдіктің биологиялық салдары әлі толық зерттелмеген. Z-ДНҚ аймақтары белгілі бір гендердің экспрессиясын реттеуде рөл атқарады және генетикалық рекомбинацияға қатысады деп саналады.
ДНҚ-ның Z-пішіні сол жақ қос спирал болып табылады, оның фосфодиэфирлік арқауы молекула осінің бойымен иректелген. Молекуланың (зигзаг)-ДНҚ атауы осыдан. Z-ДНҚ табиғатта ең аз бұралған (бір айналымда 12 негіз жұбы) және ең жұқа. Көршілес нуклеотидтер арасындағы қашықтық 0,38 нм, катушка ұзындығы 4,56 нм, Z-ДНҚ диаметрі 1,8 нм. Сонымен қатар, сыртқы түріБұл ДНҚ молекуласы бір ойықтың болуымен ерекшеленеді.
ДНҚ-ның Z-формасы прокариоттық және эукариоттық жасушаларда табылған. Бүгінгі күні ДНҚ-ның Z-формасы мен В-формасын ажырата алатын антиденелер алынды. Бұл антиденелер Drosophila (Dr. melanogaster) сілекей безі жасушаларының алып хромосомаларының белгілі бір аймақтарымен байланысады. Бұл хромосомалардың әдеттен тыс құрылымына байланысты байланыстыру реакциясын орындау оңай, оларда тығыз аймақтар (дискілер) азырақ тығыз аймақтармен (дискілер) қарама-қайшы келеді. Z-ДНҚ аймақтары дискілерде орналасқан. Бұдан шығатыны, Z пішінінің жеке бөлімдерінің өлшемдері әлі белгісіз болса да, табиғи жағдайда Z-формасы шын мәнінде бар.

(жылжытқыштар) - ДНҚ-дағы ең танымал және жиі кездесетін базалық тізбектер. Палиндрома - солдан оңға қарай және керісінше бірдей оқылатын сөз немесе сөз тіркесі. Мұндай сөздердің немесе сөз тіркестерінің мысалдары: ХУТ, КАЗАК, СУ ТАСКЫН, АЗОРНЫҢ ТАБЫСЫНА ТҮСІП ТҰРҒАН РАҚШ. ДНҚ бөлімдеріне қолданылғанда бұл термин (палиндром) тізбек бойынша оңнан солға және солдан оңға қарай нуклеотидтердің бірдей кезектесуін білдіреді («сашық» сөзіндегі әріптер сияқты, т.б.).
Палиндрома екі ДНҚ тізбегіне қатысты екінші ретті симметрияға ие негізгі тізбектердің инверттелген қайталануларының болуымен сипатталады. Мұндай тізбектер, белгілі себептермен, өзін-өзі толықтырады және шаш қыстырғыштарын немесе крест тәрізді құрылымдарды қалыптастыруға бейім (Cурет). Шаш қыстырғыштары реттеуші ақуыздарға хромосоманың ДНҚ-ның генетикалық мәтіні көшірілетін орынды тануға көмектеседі.
Бір ДНҚ тізбегінде инверттелген қайталану болған жағдайда мұндай реттілік айна қайталануы деп аталады. Айна қайталануы өзін-өзі толықтыратын қасиеттерге ие емес, сондықтан шаш қыстырғыштарын немесе крест тәрізді құрылымдарды қалыптастыруға қабілетті емес. Бұл түрдегі тізбектер барлық дерлік үлкен ДНҚ молекулаларында кездеседі және бірнеше негізгі жұптан бірнеше мың негіз жұбына дейін болуы мүмкін.
Эукариоттық жасушаларда крест тәрізді құрылымдар түріндегі палиндромдардың болуы дәлелденбеген, дегенмен бірқатар крест тәрізді құрылымдар in vivo E. coli жасушаларында табылған. РНҚ немесе бір тізбекті ДНҚ-да өзін-өзі толықтыратын тізбектердің болуы ерітінділердегі нуклеин тізбегінің көптеген «шаш түйреуіштердің» пайда болуымен сипатталатын белгілі бір кеңістіктік құрылымға қатпарлануының негізгі себебі болып табылады.

ДНҚ-ның Н-формасы- бұл ДНҚ-ның үш тізбегінен түзілетін спираль - ДНҚ-ның үштік спиралі. Бұл Гугстин жұбы деп аталатын пайда болуымен оның үлкен ойығына сәйкес келетін үшінші бір тізбекті ДНҚ тізбегі бар Уотсон-Крик қос спиралының кешені.
Мұндай триплекстің түзілуі ДНҚ қос спиралының қосылуы нәтижесінде оның қимасының жартысы қос спираль түрінде қалатындай, ал екінші жартысы ажыратылатындай болады. Бұл жағдайда ажыратылған спиральдардың бірі қос спиралдың бірінші жартысы - үштік спиральмен жаңа құрылымды құрайды, ал екіншісі құрылымсыз, бір талшықты кесінді түрінде шығады. Бұл құрылымдық ауысудың ерекшелігі - протондары жаңа құрылымды тұрақтандыратын ортаның рН-ға күрт тәуелділігі. Осы ерекшелігіне байланысты жаңа құрылым ДНҚ-ның Н-формасы деп аталды, оның түзілуі айна қайталануы болып табылатын гомопурин-гомопиримидин аймақтары бар супер ширатылған плазмидаларда табылды.
Кейінгі зерттеулерде кейбір гомопуринді-гомопиримидинді қос тізбекті полинуклеотидтердің құрылымдық ауысу мүмкіндігі үш тізбекті құрылымның қалыптасуымен анықталды:
- бір гомопурин және екі гомопиримидин тізбегі ( Py-Pu-Py триплексі) [Гугстиннің өзара әрекеттесуі].
Py-Pu-Py триплексінің құрамдас блоктары канондық изоморфты CGC+ және TAT триадалары болып табылады. Триплексті тұрақтандыру үшін CGC+ триадасының протондануы қажет, сондықтан бұл триплекстер ерітіндінің рН-ына тәуелді.
- бір гомопиримидин және екі гомопурин тізбегі ( Py-Pu-Pu триплексі) [Гугстиннің кері әсерлесуі].
Py-Pu-Pu триплексінің құрамдас блоктары канондық изоморфты CGG және TAA триадалары болып табылады. Py-Pu-Pu триплекстерінің маңызды қасиеті олардың тұрақтылығының қосарланған зарядталған иондардың болуына тәуелділігі болып табылады және әртүрлі тізбектегі триплекстерді тұрақтандыру үшін әртүрлі иондар қажет. Py-Pu-Pu триплекстерінің түзілуі олардың құрамдас нуклеотидтерінің протондалуын қажет етпейтіндіктен, мұндай триплекстер бейтарап рН кезінде болуы мүмкін.
Ескерту: Гугстиннің тура және кері әрекеттесуі 1-метилтиминнің симметриясымен түсіндіріледі: 180 ° айналу O4 атомының орнын O2 атомы алып жатқандығына әкеледі, бұл ретте сутегі байланыстары жүйесі сақталады.
Үштік спиральдың екі түрі бар:
- Үшінші жіптің полярлығы Уотсон-Крик дуплексінің гомопурин тізбегінің полярлығымен бірдей параллель үштік спиральдар
- антипараллельді үштік спиральдар, оларда үшінші және гомопуриндік тізбектердің полярлықтары қарама-қарсы.

G-төрт жақты- 4 тізбекті ДНҚ. Мұндай құрылым, егер төрт гуанин болса, түзіледі, олар G-квадруплекс деп аталатын - төрт гуаниннің дөңгелек биін құрайды.
Мұндай құрылымдардың пайда болу мүмкіндігі туралы алғашқы кеңестер Уотсон мен Криктің серпінді жұмысынан көп бұрын - 1910 жылы алынған. Содан кейін неміс химигі Ивар Бэнг ДНҚ құрамдастарының бірі – гуаноз қышқылы жоғары концентрацияда гель түзетінін, ал ДНҚ-ның басқа компоненттерінде мұндай қасиет жоқ екенін анықтады.
1962 жылы рентгендік дифракция әдісін қолдана отырып, бұл гельдің жасушалық құрылымын орнату мүмкін болды. Ол бір-бірін шеңбер бойымен біріктіріп, өзіне тән шаршы құрайтын төрт гуанин қалдықтарынан тұратын болып шықты. Орталықта байланыс металл ионымен (Na, K, Mg) тіреледі. ДНҚ-да гуанин көп болса, дәл осындай құрылымдар түзілуі мүмкін. Бұл жалпақ шаршылар (G-квартеттері) жеткілікті тұрақты, тығыз құрылымдарды (G-төртбұрыштар) қалыптастыру үшін қабаттастырылған.
ДНҚ-ның төрт бөлек тізбегі төрт тізбекті кешендерге тоқылуы мүмкін, бірақ бұл ерекшелік. Көбінесе нуклеин қышқылының бір тізбегі жай ғана түйінге байланып, өзіне тән қалыңдауды (мысалы, хромосомалардың ұштарында) құрайды немесе қос тізбекті ДНҚ гуанинге бай кейбір жерлерде жергілікті төртбұрышты құрайды.
Ең көп зерттелгені хромосомалардың ұштарында – теломерлерде және онкопромоторларда төртбұрыштардың болуы. Дегенмен, адам хромосомаларында мұндай ДНҚ-ның локализациясы туралы толық түсінік әлі белгісіз.
Сызықтық түрдегі ДНҚ-ның барлық осы ерекше құрылымдары ДНҚ-ның В-формасымен салыстырғанда тұрақсыз. Дегенмен, ДНҚ көбінесе топологиялық шиеленістің сақиналы түрінде болады, егер ол супер орам деп аталады. Бұл жағдайларда канондық емес ДНҚ құрылымдары оңай қалыптасады: Z-формалары, «кресттер» және «шаш түйреуіштер», Н-формалары, гуаниндік төрт плекстер және i-мотив.
- Асқын ширатылған пішін – пентоз-фосфатты омыртқаға зақым келтірмей, жасуша ядросынан босатылған кезде белгіленеді. Ол өте бұралған жабық сақиналар түрінде болады. Өте бұралған күйде ДНҚ қос спиралі кем дегенде бір рет «өзіне бұрылады», яғни оның құрамында кем дегенде бір суперкоир бар (сегіздік фигура пішінін алады).
- ДНҚ-ның босаңсыған күйі – бір үзіліспен байқалады (бір тізбектің үзілуі). Бұл жағдайда супер катушкалар жойылып, ДНҚ тұйық сақина пішінін алады.
- ДНҚ-ның сызықтық түрі қос спиралдың екі тізбегі үзілгенде байқалады.
ДНҚ-ның үшінші реттік құрылымы
ДНҚ-ның үшінші реттік құрылымықос жіпшелі молекуланың кеңістікте қосымша бұралуы – оның асқын ширатылуы нәтижесінде түзіледі. Эукариот жасушаларында ДНҚ молекуласының супер ширатылуы прокариоттардан айырмашылығы белоктармен комплекстер түрінде жүзеге асады.
Эукариоттық ДНҚ-ның барлығы дерлік ядролардың хромосомаларында орналасады, оның аз ғана бөлігі митохондрияларда, өсімдіктер мен пластидтерде кездеседі. Эукариот жасушаларының хромосомаларының негізгі заты (оның ішінде адам хромосомалары) екі тізбекті ДНҚ, гистон және гистон емес ақуыздардан тұратын хроматин.
Хроматиннің гистондық белоктары
Гистондар хроматиннің 50%-ын құрайтын қарапайым белоктар. Жануарлар мен өсімдіктердің барлық зерттелген жасушаларында гистондардың бес негізгі класы табылды: H1, H2A, H2B, H3, H4, олардың мөлшері, аминқышқылдарының құрамы және зарядтары бойынша ерекшеленеді (әрдайым оң).
Сүтқоректілердің H1 гистоны құрамында шамамен 215 аминқышқылдары бар бір полипептидтік тізбектен тұрады; басқа гистондардың өлшемдері 100-ден 135 аминқышқылдарына дейін өзгереді. Олардың барлығы спиральданған және диаметрі шамамен 2,5 нм глобулға айналдырылған, лизин мен аргинин оң зарядталған аминқышқылдарының әдеттен тыс көп мөлшерін қамтиды. Гистондар ацетилденуі, метилденуі, фосфорлануы, поли(АДФ)-рибозилденуі мүмкін, ал Н2А және Н2В гистондары убиквитинмен ковалентті түрде байланысуы мүмкін. Гистондардың құрылымы мен қызметтерін орындауында мұндай модификациялардың рөлі қандай екендігі әлі толық анықталмаған. Бұл олардың ДНҚ-мен әрекеттесу және гендердің әрекетін реттеу механизмдерінің бірін қамтамасыз ету қабілеті деп болжанады.
Гистондар ДНҚ-мен негізінен ДНҚ-ның теріс зарядты фосфат топтары мен гистондардың оң зарядталған лизин және аргинин қалдықтары арасында түзілген иондық байланыстар (тұзды көпірлер) арқылы әрекеттеседі.
Хроматиннің гистонды емес ақуыздары
Гистон емес белоктар гистондардан айырмашылығы өте алуан түрлі. ДНҚ-байланыстырушы гистонды емес ақуыздардың 590-ға дейін әртүрлі фракциялары бөлініп алынды. Оларды қышқылдық белоктар деп те атайды, өйткені олардың құрылымында қышқыл аминқышқылдары басым болады (олар полианиондар). Хроматин белсенділігінің спецификалық реттелуі әртүрлі гистондық емес ақуыздармен байланысты. Мысалы, ДНҚ репликациясы мен экспрессиясы үшін маңызды ферменттер хроматинмен уақытша байланыса алады. Басқа белоктар, әртүрлі реттеу процестеріне қатысатындар, ДНҚ-мен белгілі бір тіндерде немесе дифференциацияның белгілі бір кезеңдерінде ғана байланысады. Әрбір ақуыз ДНҚ нуклеотидтерінің белгілі бір тізбегіне (ДНҚ учаскесі) комплементар болып табылады. Бұл топқа мыналар кіреді:
- сайтқа тән мырыш саусақ ақуыздарының отбасы. Әрбір «мырыш саусақ» 5 нуклеотидтік жұптан тұратын белгілі бір учаскені таниды.
- сайтқа тән белоктар тұқымдасы – гомодимерлер. Мұндай белоктың ДНҚ-мен жанасатын фрагменті «спираль-бұрылу-спираль» құрылымына ие.
- жоғары қозғалғыш протеиндер (HMG протеиндері - ағылшын тілінен, жоғары қозғалғыш гель ақуыздары) хроматинмен үнемі байланыста болатын құрылымдық және реттеуші белоктар тобы болып табылады. Олардың молекулалық салмағы 30 кД-ден аз және зарядталған аминқышқылдарының жоғары мөлшерімен сипатталады. Төмен молекулалық салмағына байланысты HMG протеиндері полиакриламидті гельді электрофорез кезінде жоғары қозғалғыш.
- репликация, транскрипция және жөндеу ферменттері.
ДНҚ мен РНҚ синтезіне қатысатын құрылымдық, реттеуші белоктар мен ферменттердің қатысуымен нуклеосома жіпі белоктар мен ДНҚ-ның жоғары конденсацияланған кешеніне айналады. Алынған құрылым бастапқы ДНҚ молекуласынан 10 000 есе қысқа.

Хроматин
Хроматин – ядролық ДНҚ бар белоктар кешені және бейорганикалық заттар. Хроматиннің көп бөлігі белсенді емес. Оның құрамында тығыз оралған, конденсацияланған ДНҚ бар. Бұл гетерохроматин. Экспрессияланбаған аймақтардан тұратын конститутивтік, генетикалық белсенді емес хроматин (серіктік ДНҚ) және факультативті - бірқатар ұрпақтарда белсенді емес, бірақ белгілі бір жағдайларда экспрессиялауға қабілетті.
Белсенді хроматин (эухроматин) конденсацияланбаған, яғни. тығыз емес оралған. Әртүрлі жасушаларда оның мөлшері 2-ден 11% -ға дейін болады. Ми жасушаларында ол ең көп - 10-11%, бауыр жасушаларында - 3-4 және бүйректер - 2-3%. Эухроматиннің белсенді транскрипциясы бар. Сонымен бірге оның құрылымдық ұйымдастырылуы белгілі бір ағза түріне тән бірдей ДНҚ генетикалық ақпаратты мамандандырылған жасушаларда әртүрлі тәсілдермен пайдалануға мүмкіндік береді.
Электрондық микроскопта хроматиннің кескіні моншақтарға ұқсайды: жіп тәрізді көпірлермен бөлінген өлшемі шамамен 10 нм сфералық қалыңдаулар. Бұл шар тәрізді қалыңдауды нуклеосомалар деп атайды. Нуклеосома хроматиннің құрылымдық бірлігі болып табылады. Әрбір нуклеосома нуклеосома өзегінде 1,75 солға айналу түзетін 146 бит ұзындықтағы аса ширатылған ДНҚ сегментін қамтиды. Нуклеосомалық өзек – H2A, H2B, H3 және H4 гистондарынан, әр типті екі молекуладан (9-сурет) тұратын гистонды октамер, диаметрі 11 нм, қалыңдығы 5,7 нм дискіге ұқсайды. Бесінші гистон, Н1, нуклеосомалық ядроның бөлігі емес және гистон октамерінің айналасында ДНҚ-ның айналу процесіне қатыспайды. Ол қос спиралдың нуклеосомалық өзекке енетін және шығатын нүктелерінде ДНҚ-мен байланысады. Бұл ДНҚ-ның интеркоралық (байланыстырушы) бөлімдері, олардың ұзындығы жасуша түріне байланысты 40-тан 50 жұп нуклеотидке дейін өзгереді. Нәтижесінде нуклеосомалардың құрамына кіретін ДНҚ фрагментінің ұзындығы да өзгереді (186-дан 196 жұп нуклеотидке дейін).
Нуклеосоманың құрамында ДНҚ-ның 90%-ға жуығы болады, қалған бөлігі байланыстырушы болып табылады. Нуклеосомалар «тыныш» хроматиннің фрагменттері болып табылады, ал байланыстырушы белсенді. Алайда нуклеосомалар жайылып, сызықты болуы мүмкін. Бүктелген нуклеосомалар қазірдің өзінде белсенді хроматин болып табылады. Бұл функцияның құрылымға тәуелділігін анық көрсетеді. Глобулярлы нуклеосомалардың құрамында хроматин неғұрлым көп болса, соғұрлым оның белсенділігі аз болады деп болжауға болады. Әлбетте, әртүрлі жасушаларда тыныштық хроматинінің тең емес үлесі осындай нуклеосомалардың санымен байланысты.
Электрондық микроскопиялық фотосуреттерде оқшаулану жағдайлары мен созылу дәрежесіне байланысты хроматин тек қалыңдауы бар ұзын жіп - нуклеосомалардың «моншақтары» ретінде ғана емес, сонымен қатар диаметрі қысқарақ және тығызырақ фибрил (талшық) ретінде де көрінуі мүмкін. 30 нм, оның түзілуі ДНҚ-ның линкер аймағымен және Н3 гистонымен байланысты Н1 гистонының өзара әрекеттесуі кезінде байқалады, бұл диаметрі 30 нм соленоидтың пайда болуымен бір айналымда алты нуклеосома спиралының қосымша бұралуына әкеледі. . Бұл жағдайда гистон протеині бірқатар гендердің транскрипциясына кедергі жасап, осылайша олардың белсенділігін реттей алады.
ДНҚ-ның жоғарыда сипатталған гистондармен әрекеттесуі нәтижесінде орташа диаметрі 2 нм және ұзындығы 57 нм болатын 186 негіздік жұп ДНҚ қос спиралының сегменті диаметрі 10 нм және ұзындығы бар спиральға айналады. 5 нм. Бұл спиралдың кейіннен диаметрі 30 нм талшыққа қысылуымен конденсация дәрежесі тағы алты есе артады.
Сайып келгенде, ДНҚ дуплексін бес гистонмен қаптау 50 еселік ДНҚ конденсациясына әкеледі. Дегенмен, конденсацияның мұндай жоғары дәрежесінің өзінде метафаза хромосомасындағы ДНҚ-ның 50 000-100 000 есе дерлік тығыздалуын түсіндіре алмайды. Өкінішке орай, хроматиннің метафаза хромосомасына дейін одан әрі оралуының егжей-тегжейлері әлі белгісіз, сондықтан бұл процестің жалпы ерекшеліктерін ғана қарастыруға болады.
Хромосомалардағы ДНҚ тығыздалу деңгейлері
Әрбір ДНҚ молекуласы бөлек хромосомаға оралған. Адамның диплоидты жасушаларында 46 хромосома бар, олар жасуша ядросында орналасқан. Жасушаның барлық хромосомаларының ДНҚ-ның жалпы ұзындығы 1,74 м, бірақ хромосомалары жинақталған ядроның диаметрі миллиондаған есе аз. ДНҚ-ның хромосомалардағы және жасуша ядросындағы хромосомалардың мұндай ықшам қаптамасы ДНҚ-мен белгілі бір реттілікпен әрекеттесетін әртүрлі гистондық және гистондық емес ақуыздар арқылы қамтамасыз етіледі (жоғарыдан қараңыз). Хромосомалардағы ДНҚ-ның тығыздалуы оның сызықтық өлшемдерін шамамен 10 000 есе - шартты түрде 5 см-ден 5 микронға дейін азайтуға мүмкіндік береді. Тығыздаудың бірнеше деңгейі бар (Cурет 10).


- ДНҚ қос спиралі диаметрі 2 нм және ұзындығы бірнеше см болатын теріс зарядты молекула.
- нуклеосомалық деңгей- хроматин электронды микроскопта «моншақтардың» - нуклеосомалардың - «жіптегі» тізбегі ретінде көрінеді. Нуклеосома эухроматинде де, гетерохроматинде де, фазааралық ядрода және метафаза хромосомаларында болатын әмбебап құрылымдық бірлік.
Тығыздаудың нуклеосомалық деңгейі арнайы белоктармен - гистондармен қамтамасыз етіледі. Сегіз оң зарядталған гистон домендері теріс зарядталған ДНҚ молекуласы айналатын нуклеосоманың өзегін (өзегін) құрайды. Бұл 7 есе қысқаруды береді, ал диаметрі 2-ден 11 нм-ге дейін артады.
- электромагниттік деңгей
Хромосома ұйымының соленоидтық деңгейі нуклеосомалық жіпшенің бұралуымен және одан диаметрі 20-35 нм қалың фибрилдердің – соленоидтардың немесе супербидтердің түзілуімен сипатталады. Соленоидтық қадам 11 нм және бір айналымда шамамен 6-10 нуклеосома болады. Соленоидты қаптама супербидті қаптамаға қарағанда ықтималдырақ болып саналады, оған сәйкес диаметрі 20–35 нм хроматиндік фибрил әрқайсысы сегіз нуклеосомадан тұратын түйіршіктер тізбегі немесе супербидтер болып табылады. Соленоидтық деңгейде ДНҚ-ның сызықтық өлшемі 6-10 есе азаяды, диаметрі 30 нм-ге дейін артады.
- цикл деңгейі
Цикл деңгейі шамамен 30-300 кб ілмектер құра отырып, белгілі бір ДНҚ тізбегін танитын және байланыстыратын гистондық емес аймаққа тән ДНҚ-байланыстырушы белоктармен қамтамасыз етіледі. Цикл геннің экспрессиясын қамтамасыз етеді, яғни. ілмек құрылымдық қана емес, сонымен қатар функционалдық формация. Бұл деңгейде қысқару 20-30 есе болады. Диаметрі 300 нм-ге дейін артады. Цитологиялық препараттарда амфибиялардың ооциттеріндегі ілмек тәрізді «шам қылшық» құрылымдарын көруге болады. Бұл ілмектер аса ширатылған болып көрінеді және хроматин транскрипциясы мен репликациясының бірліктеріне сәйкес келетін ДНҚ домендерін білдіреді. Арнайы белоктар ілмектердің негіздерін және, мүмкін, олардың кейбір ішкі аймақтарын бекітеді. Цикл тәрізді домендік ұйым метафазалық хромосомалардағы хроматиннің жоғары ретті спиральдық құрылымдарға қатпарлануын жеңілдетеді.
- домен деңгейі
Хромосомаларды ұйымдастырудың домендік деңгейі жеткілікті түрде зерттелмеген. Бұл деңгейде ілмектік домендердің түзілуі атап өтіледі - қалыңдығы 25-30 нм жіпшелердің (фибрилдердің) құрылымдары, құрамында 60% ақуыз, 35% ДНҚ және 5% РНҚ бар, жасушалық циклдің барлық фазаларында іс жүзінде көрінбейді. митозды қоспағанда және жасуша ядросына біршама кездейсоқ бөлінеді. Цитологиялық препараттарда амфибиялардың ооциттеріндегі ілмек тәрізді «шам қылшық» құрылымдарын көруге болады.
Циклдік домендер өздерінің негізімен бірге MAR/SAR реттілігі деп аталатын кірістірілген тіркеме тораптарындағы ядроішілік ақуыз матрицасына бекітіледі (MAR, ағылшын матрицасымен байланысты аймақтан; SAR, ағылшын тіліндегі тіреу аймақтарынан) - ДНҚ бірнеше жүздеген ұзын негіз жұптарын бөледі, олар A/T негіз жұптарының жоғары құрамымен (>65%) сипатталады. Әрбір домен репликацияның бір бастауына ие болып көрінеді және автономды аса ширатылған бірлік ретінде қызмет етеді. Кез келген циклдік домен жұмысы үйлестірілген болуы мүмкін көптеген транскрипция бірліктерін қамтиды - бүкіл домен белсенді немесе белсенді күйде.
Домен деңгейінде хроматиннің дәйекті орау нәтижесінде ДНҚ-ның сызықтық өлшемдері шамамен 200 есе (700 нм) азаяды.
- хромосома деңгейі
Хромосомалық деңгейде профазалық хромосома гистонды емес ақуыздардың осьтік шеңберінің айналасында ілмектік домендердің тығыздалуы арқылы метафазаға конденсацияланады. Бұл супер орамдық жасушадағы барлық H1 молекулаларының фосфорлануымен бірге жүреді. Нәтижесінде метафаза хромосомасын тығыз спиральға оралған тығыз оралған соленоидты ілмектер ретінде бейнелеуге болады. Адамның әдеттегі хромосомасында 2600-ге дейін ілмек болуы мүмкін. Мұндай құрылымның қалыңдығы 1400 нм (екі хроматид) жетеді, ал ДНҚ молекуласы 104 есе қысқарады, яғни. 5 см-ден 5 мкм-ге дейін созылған ДНҚ.
Хромосомалардың қызметтері
Экстрахромосомалық механизмдермен әрекеттесу кезінде хромосомалар қамтамасыз етеді
- тұқым қуалайтын ақпаратты сақтау
- осы ақпаратты ұялы ұйымды құру және қолдау үшін пайдалану
- тұқым қуалайтын ақпаратты оқуды реттеу
- генетикалық материалдың өздігінен қайталануы
- генетикалық материалдың аналық жасушадан аналық жасушаға ауысуы.
Хроматин аймағының активтенуі кезінде, яғни. транскрипция кезінде алдымен одан H1 гистоны, содан кейін гистон октеті қайтымды түрде жойылады. Бұл хроматиннің деконденсациялануын, 30 нм хроматин фибрилінің 10 нм жіпке бірізді ауысуын және оның әрі қарай бос ДНҚ аймақтарына таралуын тудырады, яғни. нуклеосомалық құрылымның жоғалуы.
Адамның сыртқы келбеті, кейбір әдеттері, тіпті аурулары да тұқым қуалайтыны бәрімізге белгілі. Тірі тіршілік иесі туралы осы мәліметтердің барлығы гендерде кодталған. Сонымен, бұл атышулы гендер қалай көрінеді, олар қалай жұмыс істейді және олар қайда орналасқан?
Сонымен, кез келген адамның немесе жануардың барлық гендерінің тасымалдаушысы ДНҚ болып табылады. Бұл қосылысты 1869 жылы Иоганн Фридрих Мишер ашқан.Химиялық жағынан ДНҚ дезоксирибонуклеин қышқылы болып табылады. Бұл нені білдіреді? Бұл қышқыл планетамыздағы барлық тіршіліктің генетикалық кодын қалай тасымалдайды?
ДНҚ қай жерде орналасқанын қарастырудан бастайық. Адамның жасушасында әртүрлі қызметтерді атқаратын көптеген органеллалар бар. ДНҚ ядрода орналасқан. Ядро - бұл барлық генетикалық материалды - ДНҚ-ны сақтайтын арнайы мембранамен қоршалған шағын органоид.
ДНҚ молекуласының құрылымы қандай?
Алдымен ДНҚ дегеніміз не екенін қарастырайық. ДНҚ – құрылымдық элементтерден – нуклеотидтерден тұратын өте ұзын молекула. Нуклеотидтердің 4 түрі бар - аденин (А), тимин (Т), гуанин (G) және цитозин (С). Нуклеотидтер тізбегі схемалық түрде келесідей: GGAATTSTAAG.... Бұл нуклеотидтер тізбегі ДНҚ тізбегі болып табылады.ДНҚ құрылымын алғаш рет 1953 жылы Джеймс Уотсон мен Фрэнсис Крик ашты.
Бір ДНҚ молекуласында бір-бірінің айналасында бұралған екі нуклеотид тізбегі бар. Бұл нуклеотидтік тізбектер қалай бір-біріне жабысып, спиральға айналады? Бұл құбылыс толықтауыш қасиетіне байланысты. Комплементарлылық дегеніміз тек белгілі бір нуклеотидтер (комплементарлы) екі тізбекте бір-біріне қарама-қарсы болуы мүмкін. Сонымен, қарама-қарсы аденин әрқашан тимин, ал гуанинге қарама-қарсы әрқашан тек цитозин болады. Сонымен гуанин цитозинмен, ал аденин тиминмен комплементарлы болады.Әртүрлі тізбекте бір-біріне қарама-қарсы орналасқан мұндай жұп нуклеотидтерді комплементарлы деп те атайды.
 Оны схемалық түрде келесідей көрсетуге болады:
Оны схемалық түрде келесідей көрсетуге болады:
G - C
Т - А
Т - А
С - Г
Бұл толықтауыш A - T және G - C жұптары химиялық байланысжұптың нуклеотидтері арасында, ал G мен С арасындағы байланыс А мен Т арасындағыдан күштірек. Байланыс қатаң түрде комплементарлы негіздер арасында түзіледі, яғни комплементарлы емес G мен А арасында байланыстың түзілуі мүмкін емес.
ДНҚ-ның «орауы», ДНҚ тізбегі қалай хромосомаға айналады?
Неліктен ДНҚ-ның бұл нуклеотидтік тізбектері де бір-біріне айналады? Бұл не үшін қажет? Өйткені, нуклеотидтердің саны өте үлкен және мұндай ұзын тізбектерді орналастыру үшін сізге көп орын қажет. Осы себепті ДНҚ-ның екі тізбегінің екіншісінің айналасында спираль тәрізді бұралуы бар. Бұл құбылысспиральдану деп аталады. Спиральдану нәтижесінде ДНҚ тізбектері 5-6 есе қысқарады.Кейбір ДНҚ молекулалары организммен белсенді түрде пайдаланылады, ал басқалары сирек қолданылады. Мұндай сирек қолданылатын ДНҚ молекулалары спиральданумен қатар, одан да ықшам «қаптамадан» өтеді. Мұндай ықшам қаптама суперкоилинг деп аталады және ДНҚ тізбегін 25-30 есе қысқартады!
ДНҚ спиралі қалай оралған?
 Асқын ширату үшін таяқша немесе жіп катушкасының сыртқы түрі мен құрылымы бар гистон протеиндері қолданылады. ДНҚ-ның спиральданған жіптері осы «орамдарға» - гистон протеиндеріне оралады. Осылайша, ұзын жіп өте жинақы оралып, өте аз орын алады.
Асқын ширату үшін таяқша немесе жіп катушкасының сыртқы түрі мен құрылымы бар гистон протеиндері қолданылады. ДНҚ-ның спиральданған жіптері осы «орамдарға» - гистон протеиндеріне оралады. Осылайша, ұзын жіп өте жинақы оралып, өте аз орын алады. Егер сол немесе басқа ДНҚ молекуласын қолдану қажет болса, «таратылу» процесі жүреді, яғни ДНҚ жіпі «орамадан» - гистон протеинінен (егер ол оған оралған болса) «ажыратылады» және одан шығады. спираль екі параллель тізбекке бөлінеді. Ал ДНҚ молекуласы мұндай бұрмаланбаған күйде болғанда, одан қажетті генетикалық ақпаратты оқуға болады. Оның үстіне, генетикалық ақпаратты оқу тек бұрылмаған ДНҚ жіптерінен болады!
Асқын ширатылған хромосомалардың жиынтығы деп аталады гетерохроматин, және ақпаратты оқу үшін қол жетімді хромосомалар - эвхроматин.
Гендер дегеніміз не, олардың ДНҚ-мен байланысы қандай?
Енді гендер деген не екенін қарастырайық. Қан тобын, көздің түсін, шаштың, терінің және біздің денеміздің басқа да көптеген қасиеттерін анықтайтын гендер бар екені белгілі. Ген – қатаң анықталған комбинацияда орналасқан нуклеотидтердің белгілі бір санынан тұратын ДНҚ-ның қатаң анықталған бөлімі. ДНҚ-ның қатаң анықталған бөлімінде орналасуы белгілі бір геннің өз орны бар екенін білдіреді және бұл орынды өзгерту мүмкін емес. Мұндай салыстыруды жасаған орынды: адам белгілі бір көшеде, белгілі бір үйде және пәтерде тұрады, ал адам басқа үйге, пәтерге немесе басқа көшеге өз еркімен көше алмайды. Гендегі нуклеотидтердің белгілі бір саны әрбір генде нуклеотидтердің белгілі бір саны бар екенін және олардың көп немесе аз бола алмайтынын білдіреді. Мысалы, инсулин өндірісін кодтайтын геннің ұзындығы 60 негізгі жұпты құрайды; окситоцин гормонының өндірісін кодтайтын ген 370 бит. Қатаң нуклеотидтер тізбегі әрбір ген үшін бірегей және қатаң анықталған. Мысалы, AATTAATA тізбегі инсулин өндірісін кодтайтын геннің фрагменті болып табылады. Инсулин алу үшін дәл осындай реттілік қолданылады, мысалы, адреналин алу үшін нуклеотидтердің басқа комбинациясы қолданылады. Нуклеотидтердің белгілі бір комбинациясы ғана белгілі бір «өнімді» (адреналин, инсулин және т.б.) кодтайтынын түсіну маңызды. «Өз орнында» тұрған нуклеотидтердің белгілі бір санының мұндай бірегей комбинациясы - бұл ген.
ДНҚ тізбегінде гендерден басқа «кодталмаған тізбектер» деп аталатындар орналасқан. Мұндай кодталмаған нуклеотидтер тізбегі гендердің жұмысын реттейді, хромосомалардың спирализациясына көмектеседі және геннің бастапқы және соңғы нүктелерін белгілейді. Дегенмен, осы уақытқа дейін кодталмаған тізбектердің көпшілігінің рөлі түсініксіз болып қала береді.
Хромосома дегеніміз не? жыныстық хромосомалар
Жеке адамның гендерінің жиынтығы геном деп аталады. Әрине, бүкіл геномды бір ДНҚ-ға біріктіру мүмкін емес. Геном 46 жұп ДНҚ молекуласына бөлінген. Бір жұп ДНҚ молекуласы хромосома деп аталады. Демек, адамда 46 дана болатын дәл осы хромосомалар. Әрбір хромосома гендердің қатаң анықталған жиынтығын алып жүреді, мысалы, 18-ші хромосомада көздің түсін кодтайтын гендер бар және т.б. Хромосомалар бір-бірінен ұзындығы мен пішіні бойынша ерекшеленеді. Ең көп таралған формалар X немесе Y түрінде, бірақ басқалары да бар. Адамда бір пішінді екі хромосома болады, олар жұп (жұп) деп аталады. Осындай айырмашылықтарға байланысты барлық жұп хромосомалар нөмірленеді - 23 жұп бар. Бұл #1 жұп хромосома, #2, #3 жұп және т.б. Белгілі бір белгіге жауапты әрбір ген бір хромосомада орналасқан. Мамандарға арналған заманауи нұсқаулықтарда геннің локализациясы, мысалы, келесідей көрсетілуі мүмкін: хромосома 22, ұзын қол.Хромосомалардың қандай айырмашылықтары бар?
 Хромосомалардың бір-бірінен тағы қандай айырмашылығы бар? Ұзын қол термині нені білдіреді? Х-тәрізді хромосомаларды алайық.ДНҚ жіпшелерінің қиылысуы қатаң түрде ортасында (Х) болуы мүмкін немесе орталықта емес болуы мүмкін. ДНҚ жіптерінің мұндай қиылысуы орталықта болмаған кезде, қиылысу нүктесіне қатысты кейбір ұштары ұзағырақ, басқалары, сәйкесінше қысқа болады. Мұндай ұзын ұштарды әдетте хромосоманың ұзын қолы, ал қысқа ұштарын сәйкесінше қысқа иық деп атайды. Y-тәрізді хромосомаларды негізінен ұзын қолдар алады, ал қысқалары өте кішкентай (олар тіпті схемалық суретте көрсетілмеген).
Хромосомалардың бір-бірінен тағы қандай айырмашылығы бар? Ұзын қол термині нені білдіреді? Х-тәрізді хромосомаларды алайық.ДНҚ жіпшелерінің қиылысуы қатаң түрде ортасында (Х) болуы мүмкін немесе орталықта емес болуы мүмкін. ДНҚ жіптерінің мұндай қиылысуы орталықта болмаған кезде, қиылысу нүктесіне қатысты кейбір ұштары ұзағырақ, басқалары, сәйкесінше қысқа болады. Мұндай ұзын ұштарды әдетте хромосоманың ұзын қолы, ал қысқа ұштарын сәйкесінше қысқа иық деп атайды. Y-тәрізді хромосомаларды негізінен ұзын қолдар алады, ал қысқалары өте кішкентай (олар тіпті схемалық суретте көрсетілмеген). Хромосомалардың мөлшері ауытқиды: ең үлкендері No1 және No3 жұптардың хромосомалары, ең кішілері No17, No19 жұптардың хромосомалары.
Пішіндері мен өлшемдерінен басқа хромосомалар атқаратын қызметтерінде де ерекшеленеді. 23 жұптың 22 жұбы соматикалық, 1 жұбы жыныстық. Бұл нені білдіреді? Соматикалық хромосомалар жеке адамның барлық сыртқы белгілерін, оның ерекшеліктерін анықтайды мінез-құлық жауаптары, тұқым қуалайтын психотип, яғни әрбір жеке адамның барлық ерекшеліктері мен ерекшеліктері. Жұп жыныстық хромосома адамның жынысын анықтайды: еркек немесе әйел. Адамның жыныстық хромосомаларының екі түрі бар - X (X) және Y (Y). Егер олар ХХ (х - х) ретінде біріктірілсе - бұл әйел, ал XY (х - у) болса - біздің алдымызда ер адам бар.
Тұқым қуалайтын аурулар және хромосомалардың зақымдануы
Дегенмен, геномның «бұзылуы» бар, содан кейін адамдарда генетикалық аурулар анықталады. Мысалы, 21 жұп хромосомада екі емес, үш хромосома болса, адам Даун синдромымен туылады.Аурудың басталуына әкелмейтін, керісінше, жақсы қасиеттер беретін генетикалық материалдың көптеген кішігірім «бұзылулары» бар. Генетикалық материалдың барлық «бұзылуы» мутациялар деп аталады. Ауруға немесе ағзаның қасиеттерінің нашарлауына әкелетін мутациялар теріс, ал жаңа пайдалы қасиеттердің пайда болуына әкелетін мутациялар оң деп саналады.
Дегенмен, бүгінгі күні адамдар зардап шегетін аурулардың көпшілігіне қатысты, бұл тұқым қуалайтын ауру емес, тек бейімділік. Мысалы, баланың әкесінде қант баяу сіңеді. Бұл бала қант диабетімен туылады дегенді білдірмейді, бірақ баланың бейімділігі болады. Бұл бала тәттілер мен ұн өнімдерін теріс пайдаланатын болса, онда ол қант диабетімен ауырады дегенді білдіреді.
Бүгінгі күні деп аталатын предикативтідәрі. Осы медициналық тәжірибенің бөлігі ретінде адамда бейімділіктер анықталады (тиісті гендерді анықтау негізінде), содан кейін оған ұсыныстар беріледі - қандай диетаны ұстану керек, жұмыс пен демалыс режимдерін алмас үшін қалай дұрыс ауыстыру керек. ауру.
ДНҚ-да кодталған ақпаратты қалай оқуға болады?
Бірақ ДНҚ-дағы ақпаратты қалай оқуға болады? Оны өз денесі қалай пайдаланады? ДНҚ өзі матрицаның бір түрі, бірақ қарапайым емес, бірақ кодталған. ДНҚ матрицасынан ақпаратты оқу үшін ол алдымен арнайы тасымалдаушы – РНҚ-ға беріледі. РНҚ – химиялық рибонуклеин қышқылы. Оның ДНҚ-дан айырмашылығы – ядролық мембрана арқылы жасушаға өте алады, ал ДНҚ-да мұндай қабілет жоқ (ол тек ядрода ғана болады). Кодталған ақпарат ұяшықтың өзінде пайдаланылады. Сонымен, РНҚ ядродан жасушаға кодталған ақпаратты тасымалдаушы болып табылады.РНҚ синтезі қалай жүреді, РНҚ көмегімен белок қалай синтезделеді?
 Ақпаратты «оқылуы» керек ДНҚ жіптері ашылады, арнайы фермент «құрылысшы» оларға жақындап, ДНҚ тізбегімен параллельді комплементарлы РНҚ тізбегін синтездейді. РНҚ молекуласы сонымен қатар нуклеотидтердің 4 түрінен тұрады - аденин (А), урацил (U), гуанин (G) және цитозин (С). Бұл жағдайда келесі жұптар комплементарлы болады: аденин – урацил, гуанин – цитозин. Көріп отырғаныңыздай, ДНҚ-дан айырмашылығы, РНҚ тиминнің орнына урацилді пайдаланады. Яғни, «құрылысшы» фермент келесідей жұмыс істейді: егер ол ДНҚ тізбегінде А-ны көрсе, онда ол РНҚ тізбегіне Y-ді қосады, егер G болса, онда ол С және т.б. Осылайша, транскрипция кезінде әрбір белсенді геннен шаблон түзіледі - ядролық мембрана арқылы өте алатын РНҚ көшірмесі.
Ақпаратты «оқылуы» керек ДНҚ жіптері ашылады, арнайы фермент «құрылысшы» оларға жақындап, ДНҚ тізбегімен параллельді комплементарлы РНҚ тізбегін синтездейді. РНҚ молекуласы сонымен қатар нуклеотидтердің 4 түрінен тұрады - аденин (А), урацил (U), гуанин (G) және цитозин (С). Бұл жағдайда келесі жұптар комплементарлы болады: аденин – урацил, гуанин – цитозин. Көріп отырғаныңыздай, ДНҚ-дан айырмашылығы, РНҚ тиминнің орнына урацилді пайдаланады. Яғни, «құрылысшы» фермент келесідей жұмыс істейді: егер ол ДНҚ тізбегінде А-ны көрсе, онда ол РНҚ тізбегіне Y-ді қосады, егер G болса, онда ол С және т.б. Осылайша, транскрипция кезінде әрбір белсенді геннен шаблон түзіледі - ядролық мембрана арқылы өте алатын РНҚ көшірмесі. Белгілі бір генмен кодталған ақуыздың синтезі қалай жүреді?
Ядродан шыққаннан кейін РНҚ цитоплазмаға енеді. Қазірдің өзінде цитоплазмада РНҚ матрица ретінде арнайы ферменттік жүйелерге (рибосомаларға) салынған болуы мүмкін, олар РНҚ ақпаратын басшылыққа ала отырып, ақуыздың сәйкес аминқышқылдық тізбегін синтездей алады. Белгілі болғандай, ақуыз молекуласы аминқышқылдарынан тұрады. Рибосома өсіп келе жатқан ақуыз тізбегіне қандай амин қышқылын қосу керектігін қалай біледі? Бұл үштік код негізінде жасалады. Триплет код РНҚ тізбегінің үш нуклеотидтерінің реттілігін білдіреді ( үштік,мысалы, GGU) бір амин қышқылын кодтайды (бұл жағдайда глицин). Әрбір амин қышқылы белгілі бір триплет арқылы кодталады. Осылайша, рибосома триплетті «оқады», РНҚ-ға ақпарат оқылған кезде келесі амин қышқылын қосу керектігін анықтайды. Аминқышқылдар тізбегі пайда болған кезде ол белгілі бір кеңістіктік пішінді алады және оған жүктелген ферментативті, құрылыстық, гормондық және басқа да функцияларды орындауға қабілетті ақуызға айналады.Кез келген тірі ағзаға арналған ақуыз гендік өнім болып табылады. Бұл гендердің барлық әртүрлі қасиеттерін, қасиеттерін және сыртқы көріністерін анықтайтын белоктар.
Оң жақта 2016 жылы 23 сәуірде Гиннестің рекордтар кітабына енгізілген Варна (Болгария) жағажайындағы адамдардан жасалған ең үлкен адам ДНҚ спиралы.
Дезоксирибонуклеин қышқылы. Негізгі ақпарат
ДНҚ (дезоксирибонуклеин қышқылы) - бұл тұқым қуалайтын ақпарат туралы деректерді қамтитын күрделі код, тіршіліктің бір түрі. Бұл күрделі макромолекула тұқым қуалайтын генетикалық ақпаратты ұрпақтан ұрпаққа сақтауға және беруге қабілетті. ДНҚ кез келген тірі ағзаның тұқымқуалаушылық және өзгергіштік сияқты қасиеттерін анықтайды. Ондағы кодталған ақпарат кез келген тірі ағзаның бүкіл даму бағдарламасын анықтайды. Генетикалық кіріктірілген факторлар адамның және кез келген басқа ағзаның бүкіл өмір сүру жолын алдын ала анықтайды. Сыртқы ортаның жасанды немесе табиғи әсері жеке генетикалық белгілердің жалпы ауырлығына аз ғана әсер етуі немесе бағдарламаланған процестердің дамуына әсер етуі мүмкін.
Дезоксирибонуклеин қышқылы(ДНҚ) – макромолекула (негізгі үшінің бірі, қалған екеуі РНҚ және белоктар), сақтауды, ұрпақтан ұрпаққа беруді және тірі ағзалардың дамуы мен қызмет етуінің генетикалық бағдарламасын жүзеге асыруды қамтамасыз етеді. ДНҚ-да әртүрлі РНҚ түрлері мен белоктардың құрылымы туралы ақпарат бар.
Эукариоттық жасушаларда (жануарлар, өсімдіктер және саңырауқұлақтар) ДНҚ хромосомалардың бөлігі ретінде жасуша ядросында, сондай-ақ кейбір жасуша органеллаларында (митохондриялар мен пластидтер) кездеседі. Прокариоттық организмдердің жасушаларында (бактериялар мен археялар) жасуша мембранасына ішкі жағынан нуклеоид деп аталатын дөңгелек немесе сызықты ДНҚ молекуласы бекітіледі. Оларда және төменгі эукариоттарда (мысалы, ашытқыларда) плазмидалар деп аталатын шағын автономды, негізінен дөңгелек ДНҚ молекулалары болады.
Химиялық тұрғыдан алғанда ДНҚ қайталанатын блоктардан – нуклеотидтерден тұратын ұзын полимерлі молекула. Әрбір нуклеотид азотты негізден, қанттан (дезоксирибоза) және фосфат тобынан тұрады. Тізбектегі нуклеотидтер арасындағы байланыстарды дезоксирибоза түзеді. бірге) және фосфат ( Ф) топтары (фосфодиэфирлік байланыстар).

Күріш. 2. Нуклертид азотты негізден, қанттан (дезоксирибоза) және фосфат тобынан тұрады.
Жағдайлардың басым көпшілігінде (құрамында бір тізбекті ДНҚ бар кейбір вирустардан басқа) ДНҚ макромолекуласы бір-біріне азотты негіздермен бағытталған екі тізбектен тұрады. Бұл қос тізбекті молекула спираль түрінде бұралған.
ДНҚ-да азотты негіздердің төрт түрі бар (аденин, гуанин, тимин және цитозин). Тізбектердің бірінің азотты негіздері екінші тізбектің азотты негіздерімен комплементарлылық принципі бойынша сутектік байланыстар арқылы қосылады: аденин тек тиминмен ( А-Т), гуанин - тек цитозинмен ( G-C). Дәл осы жұптар ДНҚ спиральдық «баспалдақтарының» «басқыштарын» құрайды (қараңыз: 2, 3 және 4-сурет).

Күріш. 2. Азотты негіздер
Нуклеотидтердің реттілігі туралы ақпаратты «кодтауға» мүмкіндік береді әртүрлі түрлеріРНҚ, олардың ең маңыздылары ақпарат немесе шаблон (мРНҚ), рибосомалық (рРНҚ) және тасымалдау (тРНҚ). РНҚ-ның осы түрлерінің барлығы ДНҚ тізбегін транскрипция кезінде синтезделген РНҚ тізбегіне көшіру арқылы ДНҚ шаблонында синтезделеді және ақуыз биосинтезіне (трансляция процесіне) қатысады. Жасуша ДНҚ кодтау ретімен қатар реттеуші және құрылымдық функцияларды орындайтын тізбектерді қамтиды.

Күріш. 3. ДНҚ репликациясы
ДНҚ химиялық қосылыстарының негізгі комбинацияларының орналасуы және осы комбинациялар арасындағы сандық арақатынастар тұқым қуалайтын ақпаратты кодтауды қамтамасыз етеді.
Білім жаңа ДНҚ (репликация)
- Репликация процесі: ДНҚ қос спиралының ағытылуы – ДНҚ-полимераза арқылы комплементарлы тізбектердің синтезі – бірінен екі ДНҚ молекуласының түзілуі.
- Ферменттер химиялық қосылыстардың негізгі жұптары арасындағы байланысты үзген кезде қос спираль екі тармаққа «ашылады».
- Әрбір тармақ жаңа ДНҚ элементі болып табылады. Жаңа негізгі жұптар тектік тармақтағыдай реттілікпен қосылады.
Көшіру аяқталғаннан кейін ата-аналық ДНҚ-ның химиялық қосылыстарынан жасалған және онымен бірдей генетикалық кодқа ие екі тәуелсіз спираль пайда болады. Осылайша, ДНҚ ақпаратты жасушадан жасушаға тарата алады.
Толығырақ ақпарат:
НУКЛЕИН ҚЫШҚЫЛДАРЫНЫҢ ҚҰРЫЛЫМЫ

Күріш. 4 . Азотты негіздер: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеин қышқылы(ДНҚ) нуклеин қышқылдарына жатады. Нуклеин қышқылдарымономерлері нуклеотидтер болып табылатын тұрақты емес биополимерлер класы.
нуклеотидтертұрады азотты негіз, бес көміртекті көмірсуларға (пентозаға) байланысты - дезоксирибоза(ДНҚ жағдайында) немесе рибоза(РНҚ жағдайында), ол фосфор қышқылының қалдығымен (H 2 PO 3 -) біріктіріледі.
Азотты негіздерЕкі түрі бар: пиримидиндік негіздер - урацил (тек РНҚ-да), цитозин және тимин, пуриндік негіздер - аденин және гуанин.

Күріш. 5-сурет. Нуклеотидтердің құрылымы (сол жақта), ДНҚ-дағы нуклеотидтің орналасуы (төменгі жағында) және азотты негіздердің түрлері (оң жақта): пиримидин және пурин.

Пентозаның молекуласындағы көміртек атомдары 1-ден 5-ке дейін нөмірленген. Фосфат үшінші және бесінші көміртек атомдарымен біріктіріледі. Осылайша нуклеин қышқылдары бір-бірімен байланысып, нуклеин қышқылдарының тізбегін құрайды. Осылайша, біз ДНҚ тізбегінің 3' және 5' ұштарын оқшаулай аламыз:

Күріш. 6. ДНҚ тізбегінің 3' және 5' ұштарын оқшаулау
ДНҚ-ның екі тізбегі түзіледі қос спираль. Спиральдағы бұл тізбектер қарама-қарсы бағытта бағытталған. ДНҚ-ның әртүрлі тізбектерінде азотты негіздер бір-бірімен байланысады сутектік байланыстар. Аденин әрқашан тиминмен, ал цитозин әрқашан гуанинмен біріктіріледі. деп аталады толықтыру ережесі(см. толықтыру принципі).
Толықтау ережесі:
| A-T G-C |
Мысалы, бізге реттілігі бар ДНҚ тізбегі берілсе
3'-ATGTCCTAGCTGCTCG - 5',
содан кейін екінші тізбек оған қосымша болады және қарама-қарсы бағытта - 5'-ұшынан 3'-ұшына қарай бағытталады:
5'- TACAGGATCGACGAGC- 3'.

Күріш. 7. ДНҚ молекуласы тізбектерінің бағыты және азотты негіздердің сутектік байланыс арқылы қосылуы.
ДНҚ РЕПЛИКАЦИЯСЫ
ДНҚ репликациясышаблондық синтез арқылы ДНҚ молекуласын екі еселеу процесі болып табылады. Табиғи ДНҚ репликациясының көп жағдайдапраймерДНҚ синтезі үшін қысқа үзінді (қайтадан құрылды). Мұндай рибонуклеотидті праймерді примаза ферменті жасайды (прокариоттарда ДНҚ-приаза, эукариоттарда ДНҚ-полимераза) және кейіннен дезоксирибонуклеотидті полимераза ауыстырылады, ол әдетте қалпына келтіру функцияларын орындайды (ДНҚ молекуласындағы химиялық зақымданулар мен үзілістерді түзетеді).
Репликация жартылай консервативті түрде жүреді. Бұл ДНҚ-ның қос спиралының босап, оның әрбір тізбегінде комплементарлық принцип бойынша жаңа тізбек аяқталатынын білдіреді. Осылайша, еншілес ДНҚ молекуласында ата-аналық молекуладан бір жіп және жаңадан синтезделген бір тізбек болады. Репликация ата-аналық жіптің 3' - 5' бағытында жүреді.

Күріш. 8. ДНҚ молекуласының репликациясы (екі еселенуі).
ДНҚ синтезі- бұл бірінші көзқараста көрінетіндей күрделі процесс емес. Егер сіз бұл туралы ойласаңыз, алдымен синтездің не екенін анықтауыңыз керек. Бұл бір нәрсені біріктіру процесі. Жаңа ДНҚ молекуласының түзілуі бірнеше сатыда жүреді:
1) ДНҚ топоизомеразасы, репликациялық айырдың алдында орналасқан, ДНҚ-ны оның орауы мен шешілуін жеңілдету үшін кеседі.
2) ДНҚ геликазасы топоизомеразадан кейін ДНҚ спиралының «ашу» процесіне әсер етеді.
3) ДНҚ-байланыстырушы белоктар ДНҚ жіпшелерінің байланысуын жүзеге асырады, сонымен қатар олардың бір-біріне жабысуын болдырмай, тұрақтануын жүзеге асырады.
4) ДНҚ полимераза δ(дельта) , репликация шанышқысының қозғалыс жылдамдығымен үйлестірілген, синтезді орындайдыжетекшітізбектереншілес ұйым Матрицада 5" → 3" бағытында ДНҚаналық ДНҚ жіптері оның 3" ұшынан 5" ұшына дейін (жылдамдығы секундына 100 негізгі жұпқа дейін) бағытта. Бұл оқиғалар аналықДНҚ тізбектері шектеулі.

Күріш. 9. ДНҚ репликация процесінің сызбалық көрінісі: (1) артта қалған жіп (кешті тізбек), (2) жетекші тізбек (жетекші тізбек), (3) ДНҚ полимераза α (Полα), (4) ДНҚ лигаза, (5) РНҚ -праймер, (6) Примаза, (7) Оказаки фрагменті, (8) ДНҚ полимераза δ (Полδ ), (9) Геликаза, (10) Бір тізбекті ДНҚ-байланыстырушы ақуыздар, (11) Топоизомераза.
Артта қалған еншілес ДНҚ тізбегінің синтезі төменде сипатталған (төменде қараңыз). схемарепликация шанышқысы және репликация ферменттерінің қызметі)
ДНҚ репликациясы туралы қосымша ақпаратты қараңыз
5) Бастапқы молекуланың басқа тізбегі шешіліп, тұрақтанғаннан кейін бірден ол қосыладыДНҚ полимераза α(альфа)ал 5 «→3» бағытында праймер (РНҚ праймер) – ұзындығы 10-нан 200 нуклеотидке дейінгі ДНҚ шаблонындағы РНҚ тізбегі синтезделеді. Осыдан кейін ферментДНҚ тізбегінен жойылады.
Орнына ДНҚ полимеразаα
праймердің 3 дюймдік ұшына бекітілгенДНҚ полимеразаε
.
6)
ДНҚ полимеразаε
(эпсилон) праймерді ұзартуды жалғастыратын сияқты, бірақ субстрат ретінде ендірілгендезоксирибонуклеотидтер(150-200 нуклеотид мөлшерінде). Нәтижесінде екі бөліктен қатты жіп пайда болады -РНҚ(яғни праймер) және ДНҚ.
ДНҚ полимераза εалдыңғысының праймеріне тап болғанша жұмыс істейдіОказаки фрагменті(біраз бұрын синтезделген). Содан кейін бұл фермент тізбектен жойылады.
7) ДНҚ полимераза β(бета) орнында тұрДНҚ полимеразалары ε,бір бағытта қозғалады (5" → 3") және олардың орнына дезоксирибонуклеотидтерді енгізе отырып, праймер рибонуклеотидтерін жояды. Фермент праймер толығымен жойылғанша жұмыс істейді, яғни. дезоксирибонуклеотидке дейін (одан да бұрын синтезделген).ДНҚ полимераза ε). Фермент өз жұмысының нәтижесі мен алдындағы ДНҚ-ны байланыстыра алмайды, сондықтан ол тізбектен шығады.
Нәтижесінде аналық жіптің матрицасында қыз ДНҚ-ның фрагменті «жатады». деп аталадыоказаки фрагменті.
8) ДНҚ лигазасы екі іргелес лигаза фрагменттері Оказаки , яғни. 5 «-сегменттің соңы, синтезделгенДНҚ полимераза ε,және 3 дюймдік тізбектің ұшы кіріктірілгенДНҚ полимеразаβ .
РНҚ ҚҰРЫЛЫМЫ
Рибонуклеин қышқылы(РНҚ) барлық тірі организмдердің жасушаларында кездесетін үш негізгі макромолекуланың бірі (қалған екеуі ДНҚ және белоктар).
ДНҚ сияқты, РНҚ әрбір буын деп аталатын ұзын тізбектен тұрады нуклеотид. Әрбір нуклеотид азотты негізден, рибоза қантынан және фосфат тобынан тұрады. Дегенмен, ДНҚ-дан айырмашылығы, РНҚ әдетте екі емес, бір тізбектен тұрады. РНҚ-дағы пентозаны дезоксирибоза емес, рибоза көрсетеді (рибозаның екінші көмірсу атомында қосымша гидроксил тобы бар). Ақырында, ДНҚ РНҚ-дан азотты негіздердің құрамы бойынша ерекшеленеді: тиминнің орнына ( Т) урацил РНҚ-да болады ( У) , ол да аденинге қосымша болып табылады.
Нуклеотидтер тізбегі РНҚ-ға генетикалық ақпаратты кодтауға мүмкіндік береді. Барлық жасушалық организмдер ақуыз синтезін бағдарламалау үшін РНҚ (мРНҚ) пайдаланады.
Жасушалық РНҚ деп аталатын процесте қалыптасады транскрипция , яғни арнайы ферменттер арқылы жүзеге асырылатын ДНҚ шаблонында РНҚ синтезі - РНҚ полимеразалары.
Хабаршы РНҚ (мРНҚ) содан кейін деп аталатын процеске қатысады хабар тарату, анау. рибосомалардың қатысуымен мРНҚ шаблонында ақуыз синтезі. Басқа РНҚ-лар транскрипциядан кейін химиялық модификацияға ұшырайды, екіншілік және үшінші реттік құрылымдар пайда болғаннан кейін РНҚ түріне байланысты қызметтерді орындайды.

Күріш. 10. ДНҚ мен РНҚ-ның азотты негіз бойынша айырмашылығы: РНҚ-да тиминнің (Т) орнына урацил (U) болады, ол да аденинге комплементарлы.
ТРАНСКРИПЦИЯ
Бұл ДНҚ шаблонындағы РНҚ синтезінің процесі. ДНҚ сайттардың бірінде ашылады. Тізбектердің бірінде РНҚ молекуласына көшіруді қажет ететін ақпарат бар - бұл тізбек кодтау деп аталады. Кодтау тізбегіне қосымша болып табылатын ДНҚ-ның екінші тізбегі шаблондық тізбек деп аталады. Қалыптық тізбекте 3'-5' бағытта (ДНҚ тізбегі бойымен) транскрипция процесінде оған комплементарлы РНҚ тізбегі синтезделеді. Осылайша, кодтау тізбегінің РНҚ көшірмесі жасалады.
![]()
Күріш. 11. Транскрипцияның схемалық көрінісі
Мысалы, егер бізге кодтау тізбегінің реттілігі берілсе
3'-ATGTCCTAGCTGCTCG - 5',
онда комплементарлылық ережесіне сәйкес матрицалық тізбек тізбекті тасымалдайды
5'- TACAGGATCGACGAGC- 3',
ал одан синтезделген РНҚ реттілік болып табылады
ХАБАРЛАМА
Механизмді қарастырыңыз ақуыз синтезіРНҚ матрицасында, сондай-ақ генетикалық код және оның қасиеттері. Сондай-ақ, түсінікті болу үшін төмендегі сілтеме бойынша тірі жасушада болатын транскрипция және трансляция процестері туралы қысқаша бейнені қарауды ұсынамыз:

Күріш. 12. Ақуыз синтезі процесі: РНҚ-ның ДНҚ кодтары, ақуыздың РНҚ кодтары
ГЕНЕТИКАЛЫҚ КОД
Генетикалық код- нуклеотидтер тізбегі арқылы белоктардың аминқышқылдарының тізбегін кодтау әдісі. Әрбір амин қышқылы үш нуклеотидтер тізбегі арқылы кодталған - кодон немесе триплет.
Про- және эукариоттардың көпшілігіне тән генетикалық код. Кестеде барлық 64 кодон және сәйкес аминқышқылдары берілген. Негізгі реті мРНҚ-ның 5"-ден 3" ұшына дейін.
Кесте 1. Стандартты генетикалық код
|
1-ші жоқ |
2-ші негіз |
3-ші жоқ |
|||||||
|
У |
C |
А |
Г |
||||||
|
У |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Тыр/Ж) |
У Г У |
(Cys/C) |
У |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
У У А |
(Лей/л) |
U C A |
У А А |
Кодонды тоқтату** |
У Г А |
Кодонды тоқтату** |
А |
||
|
У У Г |
U C G |
У А Г |
Кодонды тоқтату** |
У Г Г |
(Тр/Вт) |
Г |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(Оның/H) |
C G U |
(Arg/R) |
У |
|
|
C U C |
С С С |
C A C |
C G C |
C |
|||||
|
C U A |
С С А |
C A A |
(Gln/Q) |
CGA |
А |
||||
|
C U G |
C C G |
C A G |
C G G |
Г |
|||||
|
А |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Көк/Қ) |
А Г У |
(Ser/S) |
У |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
ӘУ А |
A C A |
А А А |
(Lys/K) |
А Г А |
А |
||||
|
А У Г |
(Кездесті/М) |
A C G |
А Г |
A G G |
Г |
||||
|
Г |
Г У У |
(Вал/В) |
G C U |
(Ала/А) |
G A U |
(Asp/D) |
G G U |
(Гли/Г) |
У |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
Г У А |
G C A |
Г А А |
(Желім) |
G G A |
А |
||||
|
Г У Г |
G C G |
Г А Г |
G G G |
Г |
|||||
Үштіктердің ішінде «тыныс белгілері» қызметін атқаратын 4 арнайы реттілік бар:
- *Үштік ТАМЫЗ, сонымен қатар метионинді кодтайтын, деп аталады кодонды бастау. Бұл кодон белок молекуласының синтезін бастайды. Осылайша, ақуыз синтезі кезінде тізбектегі бірінші амин қышқылы әрқашан метионин болады.
- **Үшемдер UAA, UAGжәне UGAшақырды кодондарды тоқтатужәне кез келген амин қышқылдарын кодтамаңыз. Бұл тізбектерде ақуыз синтезі тоқтайды.
Генетикалық кодтың қасиеттері
1. Үштік. Әрбір амин қышқылы үш нуклеотидтер тізбегі арқылы кодталған - триплет немесе кодон.
2. Үздіксіздік. Триплеттердің арасында қосымша нуклеотидтер болмайды, ақпарат үздіксіз оқылады.
3. Бірін-бірі қайталамау. Бір нуклеотид бір уақытта екі триплеттің бөлігі бола алмайды.
4. Бірегейлік. Бір кодон тек бір амин қышқылын кодтай алады.

5. Дегенерация. Бір амин қышқылы бірнеше түрлі кодондармен кодталуы мүмкін.

6. Жан-жақтылық. Генетикалық код барлық тірі ағзалар үшін бірдей.
Мысал. Бізге кодтау тізбегінің тізбегі берілген:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Матрицалық тізбек келесі реттілікке ие болады:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Енді біз осы тізбектен ақпараттық РНҚ-ны «синтездейміз»:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Ақуыз синтезі 5' → 3' бағытында жүреді, сондықтан генетикалық кодты «оқу» үшін тізбекті аудару керек:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
Енді AUG бастау кодонын табыңыз:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
Тізімді үштікке бөліңіз:
келесідей естіледі: ДНҚ-дан ақпарат РНҚ-ға (транскрипция), РНҚ-дан ақуызға (трансляция) беріледі. ДНҚ репликация арқылы да қайталануы мүмкін және кері транскрипция процесі де мүмкін, ДНҚ РНҚ шаблонынан синтезделгенде, бірақ мұндай процесс негізінен вирустарға тән.

Күріш. 13. Орталық догма молекулалық биология
ГЕНОМ: ГЕНДЕР ЖӘНЕ ХРОМОСОМАЛАР
(жалпы ұғымдар)
Геном – организмнің барлық гендерінің жиынтығы; оның толық хромосома жиынтығы.
«Геном» терминін 1920 жылы Г.Винклер бір биологиялық түрге жататын организмдердің хромосомаларының гаплоидты жиынтығындағы гендердің жиынтығын сипаттау үшін ұсынған. Бұл терминнің бастапқы мағынасы геном ұғымы генотиптен айырмашылығы жеке адамның емес, жалпы түрдің генетикалық сипаттамасы екенін көрсетті. Молекулалық генетиканың дамуымен бұл терминнің мағынасы өзгерді. Ағзалардың көпшілігінде генетикалық ақпараттың тасымалдаушысы болып табылатын, демек, геномның негізін құрайтын ДНҚ сөздің қазіргі мағынасында тек гендерді ғана қамтымайтыны белгілі. Көп бөлігіЭукариот жасушаларының ДНҚ-сы белоктар мен нуклеин қышқылдары туралы ақпаратты қамтымайтын кодталмаған («артық») нуклеотидтер тізбегі арқылы ұсынылған. Сонымен, кез келген организмнің геномының негізгі бөлігі оның гаплоидты хромосома жиынтығының бүкіл ДНҚ-сы болып табылады.
Гендер – полипептидтер мен РНҚ молекулаларын кодтайтын ДНҚ молекулаларының сегменттері.
Өткен ғасырда гендер туралы түсінігіміз айтарлықтай өзгерді. Бұрын геном бір белгіні кодтайтын немесе анықтайтын хромосоманың аймағы болды. фенотиптіккөздің түсі сияқты (көрінетін) қасиет.
1940 жылы Джордж Бидл мен Эдвард Тэтхэм геннің молекулалық анықтамасын ұсынды. Ғалымдар саңырауқұлақ спораларын өңдеді НейроспораларРентген сәулелері және ДНҚ тізбегінің өзгеруін тудыратын басқа агенттер ( мутациялар) және кейбір спецификалық ферменттерді жоғалтқан саңырауқұлақтың мутантты штаммдарын тапты, бұл кейбір жағдайларда бүкіл метаболикалық жолдың бұзылуына әкелді. Бидл мен Тэтхэм ген бір ферментті анықтайтын немесе кодтайтын генетикалық материалдың бөлімі деген қорытындыға келді. Гипотеза осылай «Бір ген, бір фермент». Бұл ұғым кейінірек анықтамаға дейін кеңейтілді «бір ген – бір полипептид», өйткені көптеген гендер ферменттер болып табылмайтын белоктарды кодтайды және полипептид күрделі ақуыз кешенінің суббірлігі бола алады.
Суретте. 14 ДНҚ триплеттері полипептидті, мРНҚ арқылы жүзеге асырылатын белоктың аминқышқылдарының тізбегін қалай анықтайтынының диаграммасын көрсетеді. ДНҚ тізбегінің бірі мРНҚ синтезі үшін шаблон рөлін атқарады, оның нуклеотидтік триплеттері (кодондары) ДНҚ трилеттеріне комплементарлы болып табылады. Кейбір бактериялар мен көптеген эукариоттарда кодтау реті кодталмаған аймақтармен үзіледі (деп аталады). интрондар).
Геннің қазіргі биохимиялық анықтамасы одан да нақтырақ. Гендер - құрылымдық немесе каталитикалық қызметі бар полипептидтер немесе РНҚ кіретін соңғы өнімдердің бастапқы тізбегін кодтайтын ДНҚ-ның барлық бөлімдері.
ДНҚ-да гендермен қатар тек реттеуші функцияны орындайтын басқа тізбектер де бар. Нормативтік реттіліктергендердің басын немесе аяқталуын белгілеуі, транскрипцияға әсер етуі немесе репликация немесе рекомбинацияның басталу орнын көрсетуі мүмкін. Кейбір гендер әртүрлі тәсілдермен экспрессиялануы мүмкін, ДНҚ-ның бір бөлігі әртүрлі өнімдердің қалыптасуына шаблон ретінде қызмет етеді.
Біз шамамен есептей аламыз ең аз ген мөлшеріаралық ақуызды кодтау. Полипептидтік тізбектегі әрбір амин қышқылы үш нуклеотидтер тізбегі арқылы кодталады; осы триплеттердің (кодондардың) тізбегі берілген генмен кодталған полипептидтегі аминқышқылдарының тізбегіне сәйкес келеді. 350 амин қышқылы қалдықтарынан тұратын полипептидтік тізбегі (орта ұзындықтағы тізбек) 1050 б.б. реттілікке сәйкес келеді. ( bp). Дегенмен, көптеген эукариоттық гендер мен кейбір прокариоттық гендер ДНҚ сегменттерімен үзіледі, олар жоқ. ақпаратты тасымалдаушыларақуыз туралы, сондықтан қарапайым есептеу көрсеткеннен әлдеқайда ұзағырақ болып шықты.
Бір хромосомада қанша ген бар?
 Күріш. 15. Прокариот (сол жақта) және эукариоттық жасушалардағы хромосомалардың көрінісі. Гистондар екі негізгі функцияны орындайтын ядролық белоктардың кең класы болып табылады: олар ядродағы ДНҚ жіпшелерін қаптауға және транскрипция, репликация және репарация сияқты ядролық процестердің эпигенетикалық реттелуіне қатысады.
Күріш. 15. Прокариот (сол жақта) және эукариоттық жасушалардағы хромосомалардың көрінісі. Гистондар екі негізгі функцияны орындайтын ядролық белоктардың кең класы болып табылады: олар ядродағы ДНҚ жіпшелерін қаптауға және транскрипция, репликация және репарация сияқты ядролық процестердің эпигенетикалық реттелуіне қатысады.
Прокариоттардың ДНҚ-сы қарапайым: олардың жасушаларында ядро болмайды, сондықтан ДНҚ нуклеоид түрінде тікелей цитоплазмада орналасады.
 Өздеріңіз білетіндей, бактерия жасушаларында ықшам құрылымға - нуклеоидқа оралған ДНҚ тізбегі түріндегі хромосома болады. прокариоттық хромосома ішек таяқшасыгеномы толығымен декодталған, дөңгелек ДНҚ молекуласы (шын мәнінде ол емес оң жақ шеңбер, бірақ басы немесе соңы жоқ цикл), 4 639 675 б.б. Бұл тізбекте шамамен 4300 белок гендері және тұрақты РНҚ молекулаларына арналған тағы 157 ген бар. AT адам геномы 24 түрлі хромосомада орналасқан шамамен 29 000 генге сәйкес келетін шамамен 3,1 миллиард базалық жұп.
Өздеріңіз білетіндей, бактерия жасушаларында ықшам құрылымға - нуклеоидқа оралған ДНҚ тізбегі түріндегі хромосома болады. прокариоттық хромосома ішек таяқшасыгеномы толығымен декодталған, дөңгелек ДНҚ молекуласы (шын мәнінде ол емес оң жақ шеңбер, бірақ басы немесе соңы жоқ цикл), 4 639 675 б.б. Бұл тізбекте шамамен 4300 белок гендері және тұрақты РНҚ молекулаларына арналған тағы 157 ген бар. AT адам геномы 24 түрлі хромосомада орналасқан шамамен 29 000 генге сәйкес келетін шамамен 3,1 миллиард базалық жұп.
Прокариоттар (бактериялар).
 Бактерия E. coliбір қос тізбекті дөңгелек ДНҚ молекуласы бар. Ол 4 639 675 б.б. және ұзындығы шамамен 1,7 мм жетеді, бұл ұяшықтың ұзындығынан асып түседі E. coliшамамен 850 рет. Нуклеоидтың бөлігі ретінде үлкен дөңгелек хромосомамен қатар, көптеген бактерияларда цитозольде еркін орналасқан бір немесе бірнеше шағын дөңгелек ДНҚ молекулалары болады. Бұл хромосомадан тыс элементтер деп аталады плазмидалар(Cурет 16).
Бактерия E. coliбір қос тізбекті дөңгелек ДНҚ молекуласы бар. Ол 4 639 675 б.б. және ұзындығы шамамен 1,7 мм жетеді, бұл ұяшықтың ұзындығынан асып түседі E. coliшамамен 850 рет. Нуклеоидтың бөлігі ретінде үлкен дөңгелек хромосомамен қатар, көптеген бактерияларда цитозольде еркін орналасқан бір немесе бірнеше шағын дөңгелек ДНҚ молекулалары болады. Бұл хромосомадан тыс элементтер деп аталады плазмидалар(Cурет 16).
Плазмидалардың көпшілігі бірнеше мың негіз жұптарынан тұрады, кейбіреулерінде 10 000 б.б. Олар генетикалық ақпаратты тасымалдайды және аналық жасушаның бөлінуі кезінде еншілес жасушаларға енетін еншілес плазмидаларды қалыптастыру үшін репликацияланады. Плазмидтер тек бактерияларда ғана емес, ашытқыларда және басқа саңырауқұлақтарда да кездеседі. Көптеген жағдайларда плазмидалар хост жасушаларына ешқандай артықшылық бермейді және олардың жалғыз жұмысы - тәуелсіз көбею. Дегенмен, кейбір плазмидалар иесіне пайдалы гендерді тасымалдайды. Мысалы, плазмидалардың құрамындағы гендер бактериялық жасушалардағы бактерияға қарсы агенттерге төзімділікті қамтамасыз ете алады. β-лактамаза генін тасымалдайтын плазмидтер пенициллин және амоксициллин сияқты β-лактамды антибиотиктерге төзімділік береді. Плазмидтер антибиотиктерге төзімді жасушалардан бірдей немесе әртүрлі бактериялардың басқа жасушаларына өтіп, сол жасушалардың да төзімді болуына әкеледі. Антибиотиктерді қарқынды қолдану патогендік бактериялар арасында антибиотиктерге төзімділікті кодтайтын плазмидалардың (сонымен қатар ұқсас гендерді кодтайтын транспозондар) таралуына ықпал ететін және бірнеше антибиотиктерге төзімді бактерия штаммдарының пайда болуына әкелетін күшті селективті фактор болып табылады. Дәрігерлер антибиотиктерді кеңінен қолданудың қауіптілігін түсіне бастады және оларды өте қажет болған жағдайда ғана тағайындайды. Осыған ұқсас себептермен ауылшаруашылық жануарларын емдеу үшін антибиотиктерді кеңінен қолдану шектелген.
Сондай-ақ қараңыз: Равин Н.В., Шестаков С.В. Прокариоттардың геномы // Вавилов атындағы Генетика және селекция журналы, 2013. V. 17. № 4/2. 972-984 беттер.
Эукариоттар.
Кесте 2. Кейбір ағзалардың ДНҚ, гендері мен хромосомалары
|
ортақ ДНҚ, б.с. |
хромосома саны* |
Гендердің шамамен саны |
|
|
ішек таяқшасы(бактерия) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(ашытқы) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(нематод) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(өсімдік) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(жеміс шыбыны) |
120 367 260 |
20 000 |
|
|
Орыза сатива(күріш) |
480 000 000 |
57 000 |
|
|
Бұлшық ет(тышқан) |
2 634 266 500 |
27 000 |
|
|
Гомо сапиенс(Адам) |
3 070 128 600 |
29 000 |
Ескерту.Ақпарат үнемі жаңартылып отырады; Қосымша ақпарат алу үшін жеке геномдық жоба веб-сайттарын қараңыз.
* Ашытқылардан басқа барлық эукариоттар үшін хромосомалардың диплоидты жиынтығы берілген. диплоидтыжинақ хромосомалар (грек тілінен аударғанда diploos – қос және eidos – көрініс) – хромосомалардың қос жиынтығы(2n), олардың әрқайсысының өзіне гомологиясы бар.
**Гаплоидты жиынтық. Ашытқылардың жабайы штаммдарында әдетте осы хромосомалардың сегіз (октаплоидты) немесе одан да көп жиынтығы болады.
***Екі Х хромосомалары бар аналықтар үшін. Еркектерде Х хромосомасы бар, бірақ Y жоқ, яғни тек 11 хромосома.
Ең кішкентай эукариоттардың бірі ашытқы жасушасының ДНҚ-сы жасушаға қарағанда 2,6 есе көп. E. coli(2-кесте). жеміс шыбындарының жасушалары Дрозофила, генетикалық зерттеудің классикалық объектісі, ДНҚ 35 есе көп, ал адам жасушаларында жасушаларға қарағанда шамамен 700 есе көп ДНҚ бар. E. coli.Көптеген өсімдіктер мен қосмекенділерде одан да көп ДНҚ бар. Эукариоттық жасушалардың генетикалық материалы хромосома түрінде ұйымдастырылған. Хромосомалардың диплоидты жиынтығы (2 n) организмнің түріне байланысты (2-кесте).
Мысалы, адамның соматикалық жасушасында 46 хромосома ( күріш. 17). Эукариоттық жасушадағы әрбір хромосома суретте көрсетілгендей. 17, а, бір өте үлкен қос тізбекті ДНҚ молекуласын қамтиды. Адамның жиырма төрт хромосомасы (22 жұп хромосома және екі жыныстық хромосома X және Y) ұзындығы бойынша 25 еседен астам ерекшеленеді. Әрбір эукариоттық хромосома гендердің белгілі бір жиынтығынан тұрады.

Күріш. 17. эукариоттық хромосомалар.а- адам хромосомасынан жұп байланысқан және конденсацияланған қарындас хроматидтер. Бұл формада эукариоттық хромосомалар репликациядан кейін және митоз кезінде метафазада қалады. б- кітап авторларының бірінің лейкоцитінен хромосомалардың толық жиынтығы. Адамның әрбір қалыпты соматикалық жасушасында 46 хромосома болады.

Тұқым қуалайтын материалды сақтауға және тасымалдауға арналған матрица ретіндегі ДНҚ мөлшері мен қызметі осы молекуланың ұйымдасуында арнайы құрылымдық элементтердің болуын түсіндіреді. Жоғары сатыдағы организмдерде ДНҚ хромосомалардың арасында таралады.
Ағзаның ДНҚ (хромосома) жиынтығы геном деп аталады. Хромосомалар жасуша ядросында орналасып, хроматин деп аталатын құрылымды құрайды. Хроматин – 1:1 қатынасындағы ДНҚ мен негізгі белоктардың (гистондар) кешені. ДНҚ ұзындығы әдетте комплементарлы нуклеотидтер жұптарының санымен (bp) өлшенеді. Мысалы, адамның 3-ші хромосомасығасыр – көлемі 160 млн бит болатын ДНҚ молекуласы. ұзындығы шамамен 1 мм, сондықтан адамның 3-ші хромосомасының сызықтық молекуласының ұзындығы 5 мм, ал гаплоидтың барлық 23 хромосомасының ДНҚ-сы (~ 3 * 10 9 bp, MR = 1,8 * 10 12) болады. жасуша - жұмыртқа немесе сперматозоид - сызықты түрде 1 м болады Жыныс жасушаларын қоспағанда, адам денесінің барлық жасушаларында (олардың шамамен 1013-і бар) хромосомалардың қос жиынтығы бар. Жасушаның бөлінуі кезінде барлық 46 ДНҚ молекуласы репликацияланып, 46 хромосомаға айналады.
Адам геномының ДНҚ молекулаларын (22 хромосома мен Х және У немесе Х және Х хромосомалары) бір-бірімен байланыстырсаңыз, ұзындығы бір метрге жуық тізбекті аласыз. Ескерту: Барлық сүтқоректілерде және басқа гетерогаметикалық аталық организмдерде аналықтардың екі Х хромосомасы (XX), ал еркектердің бір Х хромосомасы және бір У хромосомасы (XY) болады.
Адам жасушаларының көпшілігі, сондықтан мұндай жасушалардың жалпы ДНҚ ұзындығы шамамен 2 м. Ересек адамда шамамен 10 14 жасуша болады, сондықтан барлық ДНҚ молекулаларының жалпы ұзындығы 2・1011 км. Салыстыру үшін, Жердің шеңбері 4・10 4 км, ал Жерден Күнге дейінгі қашықтық 1,5・10 8 км. Міне, біздің жасушаларымызда ғажайып жинақы оралған ДНҚ бар!
Эукариоттық жасушаларда ДНҚ бар басқа органеллалар бар - бұл митохондриялар мен хлоропласттар. Митохондриялық және хлоропласт ДНҚ-ның шығу тегі туралы көптеген гипотезалар айтылды. Бүгінгі күні жалпы қабылданған көзқарас - олар қабылдаушы жасушалардың цитоплазмасына еніп, осы органеллалардың прекурсорларына айналған ежелгі бактериялардың хромосомаларының рудименті болып табылады. Митохондриялық ДНҚ митохондриялық тРНҚ мен рРНҚ-ны, сонымен қатар бірнеше митохондриялық ақуыздарды кодтайды. Митохондриялық ақуыздардың 95%-дан астамы ядролық ДНҚ-мен кодталған.
ГЕНДЕРДІҢ ҚҰРЫЛЫМЫ
Прокариоттар мен эукариоттардағы геннің құрылымын, олардың ұқсастықтары мен айырмашылықтарын қарастырайық. Ген тек бір белокты немесе РНҚ-ны кодтайтын ДНҚ бөлімі болғанына қарамастан, оған тікелей кодтау бөлігінен басқа прокариоттар мен эукариоттарда құрылымы басқаша реттеуші және басқа да құрылымдық элементтер кіреді.
кодтау реті- геннің негізгі құрылымдық-функционалдық бірлігі, онда нуклеотидтердің триплеттері кодталады.аминқышқылдарының тізбегі. Ол бастапқы кодоннан басталып, тоқтату кодонымен аяқталады.
Кодтау тізбегіне дейін және кейін аударылмаған 5' және 3' тізбектері. Олар реттеуші және көмекші функцияларды орындайды, мысалы, рибосоманың мРНҚ-ға түсуін қамтамасыз етеді.
Трансляцияланбаған және кодтаушы тізбектер транскрипция бірлігін – транскрипцияланған ДНҚ аймағын, яғни мРНҚ синтезделетін ДНҚ аймағын құрайды.
ТерминаторРНҚ синтезі тоқтайтын геннің соңында ДНҚ-ның транскрипцияланбаған аймағы.
Геннің басында реттеу аймағы, оның ішінде промоутержәне оператор.
промоутер- транскрипция инициациясында полимеразаның байланысу реттілігі. Оператор- бұл арнайы ақуыздар байланыса алатын аймақ - репрессорлар, бұл геннен РНҚ синтезінің белсенділігін төмендетуі мүмкін - басқаша айтқанда, оны азайтады өрнек.
Прокариоттардағы гендердің құрылымы
Прокариоттар мен эукариоттардағы гендердің құрылымының жалпы жоспары бір-бірінен ерекшеленбейді - олардың екеуінде промоторы мен операторы бар реттеуші аймақ, кодтау және трансляцияланбаған тізбектері бар транскрипция бірлігі және терминатор бар. Бірақ прокариоттар мен эукариоттардағы гендердің ұйымдасуы әртүрлі.

Күріш. 18. Прокариоттардағы (бактериялардағы) геннің құрылымының схемасы -сурет үлкейтілді
Оперонның басында және соңында бірнеше құрылымдық гендер үшін ортақ реттеуші аймақтар бар. Оперонның транскрипцияланған аймағынан бір мРНҚ молекуласы оқылады, онда бірнеше кодтау тізбегі бар, олардың әрқайсысының өзіндік бастау және тоқтату кодоны бар. Осы аймақтардың әрқайсысынанбір белок синтезделеді. Осылайша, Бір i-РНҚ молекуласынан бірнеше ақуыз молекулалары синтезделеді.
Прокариоттар бірнеше гендердің бір функционалды бірлікке қосылуымен сипатталады - оперон. Оперонның жұмысын басқа гендер реттеуге болады, олар оперонның өзінен айтарлықтай жойылуы мүмкін - реттеушілер. Осы геннен аударылған ақуыз деп аталады репрессор. Ол оперон операторымен байланысып, оның құрамындағы барлық гендердің экспрессиясын бірден реттейді.
Прокариоттарға да құбылыс тән транскрипция және аударма конъюгациялары.
![]()
Күріш. 19 Прокариоттардағы транскрипция мен трансляцияның конъюгациясы құбылысы – сурет үлкейтілді
Эукариоттарда транскрипция жүретін генетикалық материалдан трансляция жүретін цитоплазманы бөлетін ядролық қабықтың болуына байланысты бұл жұптасу болмайды. Прокариоттарда ДНҚ шаблонында РНҚ синтезі кезінде рибосома синтезделген РНҚ молекуласымен бірден байланыса алады. Осылайша, аударма транскрипция аяқталмай тұрып-ақ басталады. Сонымен қатар, бірнеше рибосомалар бір уақытта бір ақуыздың бірнеше молекуласын синтездей отырып, бір РНҚ молекуласымен байланыса алады.
Эукариоттардағы гендердің құрылымы
Эукариоттардың гендері мен хромосомалары өте күрделі ұйымдастырылған.
Көптеген түрлердің бактерияларында бір ғана хромосома болады және барлық жағдайда дерлік әрбір хромосомада әрбір геннің бір көшірмесі болады. рРНҚ гендері сияқты бірнеше гендер ғана бірнеше көшірмелерде болады. Гендер мен реттеуші тізбектер прокариоттардың барлық дерлік геномын құрайды. Оның үстіне әрбір ген дерлік өзі кодтайтын аминқышқылдарының тізбегіне (немесе РНҚ тізбегіне) қатаң сәйкес келеді (14-сурет).
Эукариоттық гендердің құрылымдық және қызметтік ұйымдастырылуы әлдеқайда күрделі. Эукариоттық хромосомаларды зерттеу, кейінірек эукариоттық геномдардың толық тізбегінің реттілігі көптеген тосын сыйлар әкелді. Эукариоттық гендердің көпшілігі болмаса да, қызықты ерекшелігі бар: олардың нуклеотидтер тізбегі полипептидтік өнімнің аминқышқылдарының тізбегін кодтамайтын бір немесе бірнеше ДНҚ аймақтарын қамтиды. Мұндай трансляцияланбаған кірістірулер геннің нуклеотидтер тізбегі мен кодталған полипептидтің аминқышқылдарының тізбегі арасындағы тікелей сәйкестікті бұзады. Бұл гендердегі трансляцияланбаған сегменттер деп аталады интрондар, немесе кіріктірілген тізбектер, және кодтау сегменттері болып табылады экзондар. Прокариоттарда тек бірнеше гендерде интрондар болады.
Сонымен, эукариоттарда гендердің оперондарға комбинациясы іс жүзінде жоқ, ал эукариоттық геннің кодтау тізбегі көбінесе аударылған аймақтарға бөлінеді. - экзондар, және аударылмаған бөлімдер - интрондар.
Көп жағдайда интрондардың қызметі анықталмаған. Жалпы алғанда, адам ДНҚ-ның шамамен 1,5% ғана «кодтау» болып табылады, яғни ол белоктар немесе РНҚ туралы ақпаратты алып жүреді. Дегенмен, үлкен интрондарды ескере отырып, адам ДНҚ-ның 30% гендерден тұрады екен. Гендер адам геномының салыстырмалы түрде аз бөлігін құрайтындықтан, ДНҚ-ның едәуір бөлігі есепке алынбайды.

Күріш. 16. Эукариоттардағы ген құрылысының схемасы – сурет үлкейтілді
Әрбір геннен алдымен жетілмеген немесе алдын ала РНҚ синтезделеді, оның құрамында интрондар да, экзондар да бар.
Осыдан кейін сплайсинг процесі жүреді, соның нәтижесінде интрон аймақтары кесіліп, жетілген мРНҚ түзіледі, одан белок синтезделеді.

Күріш. 20. Альтернативті сплайсинг процесі - сурет үлкейтілді
Гендердің мұндай ұйымдастырылуы, мысалы, бір геннен белоктың әртүрлі формаларын синтездеуге мүмкіндік береді, өйткені сплайсинг кезінде экзондар әртүрлі реттілікпен біріктірілуі мүмкін.

Күріш. 21. Прокариоттар мен эукариоттар гендерінің құрылысындағы айырмашылықтар – сурет үлкейтілді
МУТАЦИЯЛАР ЖӘНЕ МУТАГЕНез
мутациягенотиптің тұрақты өзгеруі, яғни нуклеотидтер тізбегінің өзгеруі деп аталады.
Мутацияға әкелетін процесс деп аталады мутагенез, және ағза бәріжасушаларында бірдей мутация бар мутант.
мутация теориясы 1903 жылы Хью де Врис алғаш рет тұжырымдаған. Оның заманауи нұсқасы келесі ережелерді қамтиды:
1. Мутациялар кенеттен, кенеттен пайда болады.
2. Мутация ұрпақтан ұрпаққа беріледі.
3. Мутациялар пайдалы, зиянды немесе бейтарап, доминантты немесе рецессивті болуы мүмкін.
4. Мутацияларды анықтау ықтималдығы зерттелетін даралар санына байланысты.
5. Ұқсас мутациялар қайталануы мүмкін.
6. Мутациялар бағытталған емес.
Мутация әртүрлі факторлардың әсерінен пайда болуы мүмкін. тудырған мутацияларды ажыратыңыз мутагендік әсерлері: физикалық (мысалы, ультракүлгін немесе радиация), химиялық (мысалы, колхицин немесе реактивті оттегі түрлері) және биологиялық (мысалы, вирустар). Мутациялар да туындауы мүмкін репликация қателері.
Пайда болу шарттарына байланысты мутациялар бөлінеді өздігінен- яғни қалыпты жағдайда пайда болған мутациялар және индукцияланған- яғни ерекше жағдайларда пайда болған мутациялар.
Мутация тек ядролық ДНҚ-да ғана емес, мысалы, митохондриялардың немесе пластидтердің ДНҚ-сында да болуы мүмкін. Осыған сәйкес, біз ажырата аламыз ядролықжәне цитоплазмалықмутациялар.
Мутациялардың пайда болуының нәтижесінде жиі жаңа аллельдер пайда болуы мүмкін. Егер мутантты аллель қалыпты аллельді жоққа шығарса, мутация деп аталады басым. Егер қалыпты аллель мутацияланғанды басса, мутация деп аталады рецессивті. Жаңа аллельдерді тудыратын мутациялардың көпшілігі рецессивті.
Мутациялар әсері бойынша ажыратылады бейімделгіш, организмнің қоршаған ортаға бейімделуінің жоғарылауына әкелетін, бейтарапбұл өмір сүруге әсер етпейді зияндыорганизмдердің қоршаған орта жағдайларына бейімделуін төмендететін және өлімге әкелетіндамуының бастапқы кезеңдерінде организмнің өлуіне әкеледі.
Салдарына сәйкес мутациялар ажыратылады, соларға әкеледі ақуыз функциясының жоғалуы, соқтыратын мутациялар пайда болуы ақуыздың жаңа қызметі бар, сондай-ақ мутациялар геннің дозасын өзгерту, және, тиісінше, одан синтезделген ақуыздың дозасы.
Мутация дененің кез келген жасушасында болуы мүмкін. Жыныс жасушасында мутация пайда болса, ол деп аталады герминальды(герминальды немесе генеративті). Мұндай мутациялар өзі пайда болған ағзада болмайды, бірақ ұрпақта мутанттардың пайда болуына әкеледі және тұқым қуалайды, сондықтан олардың генетика мен эволюция үшін маңызы зор. Егер мутация кез келген басқа жасушада пайда болса, ол деп аталады соматикалық. Мұндай мутация белгілі бір дәрежеде пайда болған ағзада көрінуі мүмкін, мысалы, қатерлі ісіктердің пайда болуына әкеледі. Бірақ мұндай мутация тұқым қуаламайды және ұрпаққа әсер етпейді.
Мутациялар геномның әртүрлі көлемдегі бөліктеріне әсер етуі мүмкін. Бөлу генетикалық, хромосомалықжәне геномдықмутациялар.
Гендік мутациялар
Бір геннен кіші масштабта болатын мутациялар деп аталады генетикалық, немесе нүктелі (нүкте). Мұндай мутациялар тізбектегі бір немесе бірнеше нуклеотидтердің өзгеруіне әкеледі. Гендік мутацияларға жатадыауыстыруларбір нуклеотидтің екіншісімен алмастырылуына әкеледі,жоюларнуклеотидтердің бірінің жоғалуына әкеледі,кірістірулер, тізбекке қосымша нуклеотидтің қосылуына әкеледі.

Күріш. 23. Гендік (нүктелік) мутациялар
Белокқа әсер ету механизмі бойынша гендік мутациялар бөлінеді:синоним, бұл (генетикалық кодтың деградациясының нәтижесінде) ақуыз өнімінің аминқышқылдық құрамының өзгеруіне әкелмейді,қате мутациялар, бұл бір амин қышқылының екіншісімен ауыстырылуына әкеледі және синтезделген ақуыздың құрылымына әсер етуі мүмкін, бірақ олар көбінесе елеусіз болса да,мағынасыз мутациялар, кодтау кодонын тоқтату кодонымен ауыстыруға әкеледі,әкелетін мутациялар қосылыстардың бұзылуы:

Күріш. 24. Мутация схемалары
Сондай-ақ, ақуызға әсер ету механизміне сәйкес мутациялар оқшауланып, соқтырады кадрдың ауысуы оқуларкірістіру және жою сияқты. Мұндай мутациялар, мағынасыз мутациялар сияқты, геннің бір нүктесінде пайда болғанымен, көбінесе белоктың бүкіл құрылымына әсер етеді, бұл оның құрылымының толық өзгеруіне әкелуі мүмкін. хромосоманың сегменті 180 градусқа айналғанда Күріш. 28. Транслокация
хромосоманың сегменті 180 градусқа айналғанда Күріш. 28. Транслокация
Күріш. 29. Дупликацияға дейінгі және кейінгі хромосома
Геномдық мутациялар
Ақырында, геномдық мутацияларбүкіл геномға әсер етеді, яғни хромосомалардың саны өзгереді. Полиплоидия ажыратылады - жасушаның плоидтылығының жоғарылауы және анеуплоидия, яғни хромосома санының өзгеруі, мысалы, трисомия (хромосомалардың бірінде қосымша гомологтың болуы) және моносомия (жоқ. хромосомадағы гомолог).
ДНҚ-ға қатысты бейне
ДНҚ-НЫҢ РЕПЛИКАЦИЯСЫ, РНҚ-НЫҢ КОДТАЛАУЫ, БЕКІЗ СИНТЕЗІ
(Бейне көрсетілмесе, ол қол жетімді
ДНҚ-ның химиялық құрылымы бойынша ( Дезоксирибонуклеин қышқылы) болып табылады биополимер, олардың мономерлері нуклеотидтер. Яғни, ДНҚ полинуклеотид. Сонымен қатар, ДНҚ молекуласы әдетте бір-біріне қатысты бұралған (көбінесе «спиральды бұралған» деп аталады) және сутегі байланыстарымен байланыстырылған екі тізбектен тұрады.
Тізбектерді солға да, оңға да (көбінесе) бүйірге бұруға болады.
Кейбір вирустарда бір тізбекті ДНҚ болады.
Әрбір ДНҚ нуклеотиді 1) азотты негізден, 2) дезоксирибозадан, 3) фосфор қышқылының қалдығынан тұрады.
Екі оң жақ ДНҚ спиралі
ДНҚ мыналарды қамтиды: аденин, гуанин, тиминжәне цитозин. Аденин және гуанин пуриндер, ал тимин мен цитозин - дейін пиримидиндер. Кейде ДНҚ-ның құрамында тиминді алмастыратын әдетте РНҚ-ға тән урацил болады.
ДНҚ молекуласының бір тізбегінің азотты негіздері басқасының азотты негіздерімен комплементарлылық принципі бойынша қатаң байланысқан: аденин тек тиминмен (олар бір-бірімен екі сутектік байланыс түзеді), ал гуанин тек цитозинмен (үш байланыс) байланысады. .
Нуклеотидтің өзіндегі азотты негіз циклдік түрдегі бірінші көміртек атомымен байланысқан. дезоксирибоза, бұл пентоз (бес көміртегі атомы бар көмірсу). Байланыс ковалентті, гликозидті (C-N). Рибозаға қарағанда дезоксирибозада гидроксил топтарының біреуі жоқ. Дезоксирибоза сақинасын төрт көміртек атомы және бір оттегі атомы құрайды. Бесінші көміртек атомы сақинадан тыс және оттегі атомы арқылы фосфор қышқылының қалдығымен байланысқан. Сондай-ақ үшінші көміртегі атомындағы оттегі атомы арқылы көрші нуклеотидтің фосфор қышқылының қалдығы бекітіледі.
Осылайша, ДНҚ-ның бір тізбегінде көрші нуклеотидтер өзара байланысқан коваленттік байланыстардезоксирибоза мен фосфор қышқылы арасында (фосфодиэфирлік байланыс). Фосфатты-дезоксирибоза арқауы түзіледі. Оған перпендикуляр ДНҚ-ның басқа тізбегіне қарай екінші тізбектің негіздерімен сутектік байланыстар арқылы байланысқан азотты негіздер бағытталған.
ДНҚ-ның құрылымы сутегі байланыстары арқылы қосылған тізбектердің магистральдары әртүрлі бағытта бағытталған («көп бағытты», «антипараллель» дейді). Бір жағы дезоксирибозаның бесінші көміртегі атомымен байланысқан фосфор қышқылымен аяқталса, екіншісі «бос» үшінші көміртек атомымен аяқталады. Яғни, бір тізбектің қаңқасы екіншісіне қатысты төңкерілген сияқты. Сонымен, ДНҚ тізбектерінің құрылымында 5 «ұш және 3» ұштары ажыратылады.
ДНҚ-ны репликациялау (екі еселеу) кезінде жаңа тізбектердің синтезі әрқашан олардың 5-ші ұшынан үшінші ұшына дейін жүреді, өйткені жаңа нуклеотидтер тек бос үшінші ұшына ғана қосылуы мүмкін.
Сайып келгенде (жанама РНҚ арқылы) ДНҚ тізбегіндегі әрбір дәйекті үш нуклеотид белоктың бір амин қышқылын кодтайды.
ДНҚ молекуласының құрылымының ашылуы 1953 жылы Ф.Крик пен Д.Уотсонның (басқа ғалымдардың алғашқы жұмыстары да ықпал етті) жұмыстарының арқасында болды. Қалай болса да Химиялық затДНҚ 19 ғасырдан бері белгілі. 1940 жылдары ДНҚ генетикалық ақпаратты тасымалдаушы екені белгілі болды.
Қос спираль ДНҚ молекуласының қайталама құрылымы болып саналады. Эукариоттық жасушаларда ДНҚ-ның басым бөлігі хромосомаларда орналасады, онда ол белоктармен және басқа заттармен байланысады, сонымен қатар тығызырақ қаптамадан өтеді.
Дезоксирибонуклеин қышқылы немесе ДНҚ генетикалық ақпараттың тасымалдаушысы болып табылады. Жасушалардағы ДНҚ-ның көп бөлігі ядрода орналасқан. Ол хромосомалардың негізгі құрамдас бөлігі болып табылады. Эукариоттарда ДНҚ митохондриялар мен пластидтерде де кездеседі. ДНҚ бір-бірімен ковалентті байланысқан мононуклеотидтерден тұрады, ұзын тармақталмаған полимерді білдіреді. ДНҚ-ны құрайтын мононуклеотидтер 4 азотты негіздің бірі (аденин, гуанин, цитозин және тимин) дезоксирибозадан және фосфор қышқылының қалдығынан тұрады. Бұл мононуклеотидтердің саны өте көп. Мысалы, құрамында бір хромосома бар прокариот жасушаларында ДНҚ молекулалық массасы 2 х 10 9 дан асатын жалғыз макромолекула болып табылады.
ДНҚ-ның бір тізбегінің мононуклеотидтері түзілуіне байланысты бір-бірімен тізбектей жалғасады ковалентті фосфодиэфирлік байланыстар бір мононуклеотидтің дезоксирибоза OH тобы мен екіншісінің фосфор қышқылының қалдығы арасында. ДНҚ-ның бір тізбегінің түзілген омыртқасының бір жағында азотты негіздер орналасады. Оларды бір жіпке салынған төрт түрлі моншақпен салыстыруға болады, өйткені. олар қант-фосфат тізбегіне байланған сияқты.
Сұрақ туындайды, бұл ұзын полинуклеотидтік тізбек жасушаның немесе тіпті бүкіл ағзаның даму бағдарламасын қалай кодтай алады? Бұл сұрақтың жауабын ДНҚ-ның кеңістіктік құрылымы қалай қалыптасатынын түсіну арқылы алуға болады. Бұл молекуланың құрылымын 1953 жылы Дж.Уотсон мен Ф.Крик ашты және сипаттады.
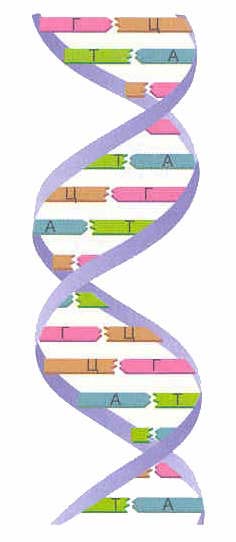
ДНҚ молекулалары бір-біріне параллель және түзілетін екі тізбек оң жақ спираль
. Бұл спиральдың ені шамамен 2 нм, бірақ оның ұзындығы жүздеген мың нанометрге жетуі мүмкін. Уотсон мен Крик ДНҚ моделін ұсынды, оған сәйкес барлық ДНҚ негіздері спираль ішінде, қант-фосфатты магистраль сыртында орналасқан. Осылайша, бір тізбектің табандары екіншісінің негіздеріне мүмкіндігінше жақын,
сондықтан олардың арасында сутектік байланыстар пайда болады. ДНҚ спиральының құрылымы оны құрайтын полинуклеотидтік тізбектерді тек қана ашылғаннан кейін бөлуге болатындай.
ДНҚ-ның екі тізбегінің максималды жақындығына байланысты оның құрамында бір түрдегі азотты негіздердің (аденин және гуанин) және басқа түрдегі (тимин және цитозин) азотты негіздерінің бірдей мөлшері бар, яғни формула жарамды: A+G=T+C. Бұл азотты негіздердің көлеміне байланысты, атап айтқанда, аденин-тимин және гуанин-цитозин жұптары арасында сутектік байланыстың пайда болуына байланысты түзілетін құрылымдардың ұзындығы шамамен 1,1 нм. Бұл жұптардың жалпы өлшемдері ДНҚ спиралының ішкі бөлігінің өлшемдеріне сәйкес келеді. Спираль жасау үшін жұп C-Tтым кішкентай болар еді жұп A-G, керісінше, тым үлкен. Яғни, ДНҚ-ның бірінші тізбегінің азотты негізі, басқа ДНҚ тізбегінің бір орнында орналасқан негізді көрсетеді. ДНҚ молекуласында бір-біріне параллель жұпталған тізбектерде орналасқан нуклеотидтердің қатаң сәйкестігі деп аталады. толықтырушылық (міндетті емес). нақты көбейту немесе репликация генетикалық ақпарат дәл ДНҚ молекуласының осы қасиетінің арқасында мүмкін болады.
ДНҚ-да биологиялық ақпарат дәл көшіріліп, ұрпақ жасушаларына берілетіндей етіп жазылады. Жасушаның бөлінуіне дейін, репликация (өзін-өзі еселеу ) ДНҚ. Әрбір тізбекте серіктестік тізбектің реттілігін толықтыратын нуклеотидтер тізбегі болғандықтан, олар іс жүзінде бірдей генетикалық ақпаратты тасымалдайды. Егер сіз жіптерді бөліп, олардың әрқайсысын екінші тізбекті құру үшін шаблон (матрица) ретінде қолдансаңыз, сіз екі жаңа бірдей ДНҚ тізбегін аласыз. Жасушада ДНҚ осылайша қайталанады.