În ce constă ADN-ul? Ce este o nucleotidă? Compoziția, structura, numărul și secvența nucleotidelor din lanțul ADN Helixul dublu al ADN stabilizează ionii
Acizii nucleici sunt substanțe moleculare ridicate constând din mononucleotide, care sunt conectate între ele într-un lanț polimeric folosind legături fosfodiester de 3 ", 5" și sunt ambalate într-un anumit mod în celule.
Acizii nucleici sunt biopolimeri de două tipuri: acid ribonucleic (ARN) și acid dezoxiribonucleic (ADN). Fiecare biopolimer este format din nucleotide care diferă prin reziduul de carbohidrați (riboză, dezoxiriboză) și una dintre bazele azotate (uracil, timină). Conform acestor diferențe, acizii nucleici și-au primit numele.
Structura acidului dezoxiribonucleic
Acizii nucleici au o structură primară, secundară și terțiară.
Structura ADN-ului primar
Structura primară a ADN-ului se numește lanț polinucleotidic liniar, în care mononucleotidele sunt legate prin legături de 3 ", 5" -fosfodiester. Materia primă pentru asamblarea lanțului de acid nucleic în celulă este nucleozida 5 "-trifosfat, care, ca urmare a îndepărtării reziduurilor de acid fosforic β și γ, este capabilă să atașeze atomul de carbon de 3" al unui alt nucleozid . Astfel, atomul de carbon de 3 "al unei dezoxiriboze se leagă covalent de atomul de carbon de 5" al celeilalte dezoxiriboze printr-un reziduu de acid fosforic și formează un lanț liniar de acid nucleic polinucleotidic. De aici și numele: legături de 3 ", 5" -fosfodiester. Bazele azotate nu iau parte la combinația de nucleotide dintr-un lanț (Fig. 1.).
O astfel de conexiune, între restul moleculei de acid fosforic a unui nucleotid și carbohidratul altuia, duce la formarea unui schelet de pentoză-fosfat al moleculei de polinucleotidă, pe care bazele azotate sunt atașate una câte una pe lateral. Secvența lor de aranjare în lanțurile moleculelor de acid nucleic este strict specifică pentru celulele diferitelor organisme, adică are o natură specifică (regula lui Chargaff).
Un lanț ADN liniar, a cărui lungime depinde de numărul de nucleotide incluse în lanț, are două capete: unul se numește capătul 3 "și conține un hidroxil liber, iar celălalt, capătul 5", conține un acid fosforic reziduu. Lanțul este polarizat și poate avea o direcție de 5 "-> 3" și 3 "-> 5". O excepție este ADN-ul circular.
„Textul” genetic al ADN-ului este compus din „cuvinte” de cod - triplete de nucleotide numite codoni. Regiunile ADN care conțin informații despre structura primară a tuturor tipurilor de ARN se numesc gene structurale.
Lanțurile de ADN polinucleodit ating dimensiuni gigantice, așa că sunt ambalate într-un anumit mod în celulă.
Studiind compoziția ADN-ului, Chargaff (1949) a stabilit legi importante cu privire la conținutul bazelor individuale de ADN. Au ajutat la descoperirea structurii secundare a ADN-ului. Aceste tipare se numesc reguli Chargaff. Regulile Chargaff
Aceste reguli indică faptul că atunci când se construiește ADN, nu trebuie respectată o corespondență destul de strictă (împerechere) între bazele de purină și pirimidină în general, ci în mod specific timina cu adenină și citozina cu guanina. Pe baza acestor reguli, inclusiv, în 1953, Watson și Crick au propus un model al structurii secundare a ADN-ului, numit dubla helix (Fig.). |

Structura secundară a ADN-ului
Structura secundară a ADN-ului este o dublă spirală, al cărei model a fost propus de D. Watson și F. Crick în 1953.
Condiții preliminare pentru crearea unui model ADN
Analizele inițiale au dat impresia că ADN-ul de orice origine conține toate cele patru nucleotide în cantități molare egale. Cu toate acestea, în anii 1940, E. Chargaff și colegii săi, ca urmare a analizei ADN-ului izolat din diferite organisme, au arătat în mod clar că bazele azotate sunt conținute în ele în diferite rapoarte cantitative. Chargaff a constatat că, deși aceste rapoarte sunt aceleași pentru ADN-ul din toate celulele aceluiași tip de organism, ADN-ul din specii diferite poate diferi semnificativ în ceea ce privește conținutul anumitor nucleotide. Acest lucru a sugerat că diferențele în raportul bazelor azotate pot fi asociate cu un fel de cod biologic. Deși raportul bazelor individuale de purină și pirimidină în diferite probe de ADN s-a dovedit a fi diferit, la compararea rezultatelor analizei, a apărut un anumit model: în toate probele, cantitatea totală de purine a fost egală cu cantitatea totală de pirimidine (A + G = T + C), cantitatea de adenină a fost egală cu cantitatea de timină (A = T), iar cantitatea de guanină - cantitatea de citozină (G = C). ADN-ul izolat din celule de mamifere a fost în general mai bogat în adenină și timină și relativ mai sărac în guanină și citozină, în timp ce în bacterii ADN-ul a fost mai bogat în guanină și citozină și relativ mai sărac în adenină și timină. Aceste date au format o parte importantă a materialului de fapt, pe baza căruia a fost construit ulterior un model al structurii ADN-ului Watson-Crick.
O altă indicație indirectă importantă a posibilei structuri a ADN a fost datele lui L. Pauling privind structura moleculelor de proteine. Pauling a arătat că sunt posibile mai multe configurații stabile diferite ale lanțului de aminoacizi dintr-o moleculă proteică. Una dintre configurațiile comune ale lanțului peptidic, α-helix, este o structură elicoidală regulată. Cu o astfel de structură, este posibilă formarea de legături de hidrogen între aminoacizi situați pe virajele adiacente ale lanțului. Pauling a descris configurația α-elicoidală a lanțului polipeptidic în 1950 și a sugerat că și moleculele ADN au probabil o structură elicoidală, fixată prin legături de hidrogen.
Cu toate acestea, cele mai valoroase informații despre structura moleculei de ADN au fost furnizate de rezultatele analizei structurale cu raze X. Razele X, care trec prin cristalul ADN, suferă difracție, adică sunt deviate în anumite direcții. Gradul și natura deformării razelor depind de structura moleculelor în sine. Modelul de difracție cu raze X (Fig. 3) oferă ochiului experimentat o serie de indicații indirecte cu privire la structura moleculelor substanței investigate. Analiza modelelor de difracție cu raze X ale ADN-ului a condus la concluzia că bazele azotate (având o formă plană) sunt stivuite ca un teanc de plăci. Modelele de difracție cu raze X au relevat trei perioade principale în structura ADN-ului cristalin: 0,34, 2 și 3,4 nm.

Modelul ADN-ului Watson-Crick
Pe baza datelor analitice ale lui Chargaff, modelele de difracție cu raze X obținute de Wilkins și cercetările chimiștilor care au furnizat informații despre distanțele exacte dintre atomii dintr-o moleculă, despre unghiurile dintre legăturile unui atom dat și dimensiunea atomi, Watson și Crick au început să construiască modele fizice ale părților constitutive individuale ale moleculei de ADN pe o anumită scară. [spectacol] .
Chiar mai devreme, se știa că nucleotidele adiacente din lanțul ADN sunt conectate prin punți de fosfodiester care leagă atomul de carbon 5 'al dezoxiribozei unui nucleotid cu atomul de carbon 3'al dezoxiribozei nucleotidei următoare. Watson și Crick nu au avut nicio îndoială că perioada de 0,34 nm corespunde distanței dintre nucleotidele consecutive din lanțul ADN. Mai mult, s-ar putea presupune că perioada de 2 nm corespunde grosimii lanțului. Și pentru a explica ce structură reală corespunde perioadei de 3,4 nm, Watson și Crick, la fel ca Pauling mai devreme, au sugerat că lanțul este răsucit sub forma unei spirale (sau, mai exact, formează o linie elicoidală, deoarece spirală în sensul strict al acestui cuvânt este obținut atunci când virajele formează o suprafață conică mai degrabă decât cilindrică în spațiu). Atunci perioada de 3,4 nm va corespunde distanței dintre virajele succesive ale acestei spirale. O astfel de spirală poate fi foarte densă sau oarecum întinsă, adică virajele sale pot fi blânde sau abrupte. Deoarece perioada de 3,4 nm este exact de 10 ori distanța dintre nucleotidele consecutive (0,34 nm), este clar că fiecare rotație completă a helixului conține 10 nucleotide. Din aceste date, Watson și Crick au reușit să calculeze densitatea unui lanț polinucleotidic răsucit într-o helică de 2 nm cu o distanță între viraje egală cu 3,4 nm. S-a dovedit că densitatea unui astfel de lanț ar fi jumătate din densitatea reală a ADN-ului, ceea ce era deja cunoscut. A trebuit să presupun că molecula de ADN constă din două catene - că este o dublă helix de nucleotide.
Următoarea sarcină a fost, desigur, clarificarea relației spațiale dintre cele două circuite care formează o dublă spirală. După testarea mai multor machete de lanț pe modelul lor fizic, Watson și Crick au constatat că toate datele disponibile se potrivesc cel mai bine cu o variantă în care două helice polinucleotidice rulează în direcții opuse; în acest caz, lanțurile formate din reziduuri de zahăr și fosfat formează suprafața unei spirale duble, iar purinele și pirimidinele sunt situate în interior. Bazele situate una față de cealaltă, aparținând a două lanțuri, sunt conectate în perechi prin legături de hidrogen; aceste legături de hidrogen sunt cele care țin lanțurile împreună, stabilind astfel configurația generală a moleculei.
Helixul dublu al ADN-ului poate fi imaginat ca o scară de frânghie în formă de spirală, astfel încât treptele sale să rămână în poziție orizontală. Apoi, două corzi longitudinale vor corespunde lanțurilor de reziduuri de zahăr și fosfat, iar barele transversale vor corespunde perechilor de baze azotate conectate prin legături de hidrogen.
Ca rezultat al studiului suplimentar al posibilelor modele, Watson și Crick au concluzionat că fiecare „bară” ar trebui să fie compusă dintr-o purină și o pirimidină; cu o perioadă de 2 nm (care corespunde diametrului helixului dublu), nu ar fi suficient loc pentru cele două purine, iar cele două pirimidine nu ar fi suficient de apropiate unele de altele pentru a forma legături de hidrogen adecvate. Un studiu aprofundat al unui model detaliat a arătat că adenina și citozina, alcătuind o combinație adecvată ca mărime, încă nu puteau fi localizate în așa fel încât să se formeze legături de hidrogen între ele. Rapoarte similare au forțat excluderea combinației guanină-timină, în timp ce combinațiile adenină-timină și guanină-citozină erau destul de acceptabile. Natura legăturilor de hidrogen este de așa natură încât adenina formează o pereche cu timina, iar guanina cu citozina. Acest concept de împerechere specifică a bazelor a permis explicarea „regulii Chargaff”, conform căreia în orice moleculă de ADN cantitatea de adenină este întotdeauna egală cu conținutul de timină, iar cantitatea de guanină este egală cu cantitatea de citozină. Două legături de hidrogen se formează între adenină și timină, iar trei se formează între guanină și citozină. Datorită acestei specificități în formarea de legături de hidrogen împotriva fiecărei adenine dintr-un lanț, timina se găsește în cealaltă; la fel, împotriva oricărei guanine, se poate găsi doar citozină. Astfel, lanțurile sunt complementare unele cu altele, adică secvența de nucleotide dintr-un lanț determină în mod unic succesiunea lor în celălalt. Cele două lanțuri rulează în direcții opuse, iar grupele lor fosfat terminale se află la capetele opuse ale helixului dublu.
Ca rezultat al cercetărilor lor, în 1953 Watson și Crick au propus un model al structurii moleculei de ADN (Fig. 3), care rămâne relevant până în prezent. Conform modelului, o moleculă de ADN constă din două lanțuri polinucleotidice complementare. Fiecare catenă ADN este o polinucleotidă formată din câteva zeci de mii de nucleotide. În el, nucleotidele adiacente formează o coloană vertebrală regulată pentoză-fosfat datorită conexiunii reziduului de acid fosforic și dezoxiriboză cu o legătură covalentă puternică. În acest caz, bazele azotate ale unui lanț polinucleotidic sunt dispuse într-o ordine strict definită în raport cu bazele azotate ale celeilalte. Alternanța bazelor azotate în lanțul polinucleotidic este neregulată.
Localizarea bazelor azotate în lanțul ADN este complementară (din grecescul "complement" - adiție), adică împotriva adeninei (A) există întotdeauna timina (T) și împotriva guaninei (G) - doar citozina (C). Acest lucru se datorează faptului că A și T, precum și G și C corespund strict unul cu celălalt, adică se completează reciproc. Această corespondență este dată de structura chimică a bazelor, care permite formarea de legături de hidrogen în perechea de purină și pirimidină. Există două conexiuni între A și T, trei între G și C. Aceste legături asigură stabilizarea parțială a moleculei de ADN în spațiu. În acest caz, stabilitatea dublei spirale este direct proporțională cu numărul de legături G≡C, care sunt mai stabile în comparație cu legăturile A = T.
Secvența cunoscută a aranjării nucleotidelor într-o catena de ADN permite, conform principiului complementarității, să stabilească nucleotidele celeilalte catene.
În plus, s-a constatat că bazele azotate cu o structură aromatică într-o soluție apoasă sunt situate una deasupra celeilalte, formând, parcă, un teanc de monede. Acest proces de formare a teancurilor de molecule organice numit miza. Lanțurile polinucleotidice ale moleculei de ADN ale modelului Watson-Crick în cauză au o stare fizico-chimică similară, bazele lor azotate sunt situate sub forma unui teanc de monede, între planurile cărora apar interacțiunile van der Waals (interacțiuni de stivuire).
Legăturile de hidrogen între bazele complementare (orizontal) și interacțiunea de stivuire între planurile bazelor din lanțul polinucleotidic datorită forțelor van der Waals (pe verticală) asigură moleculei de ADN o stabilizare suplimentară în spațiu.
Coloanele zahăr-fosfat ale ambelor lanțuri sunt orientate spre exterior, iar bazele spre interior, una spre cealaltă. Direcția lanțurilor din ADN este antiparalelă (una dintre ele are direcția 5 "-> 3", cealaltă - 3 "-> 5", adică capătul 3 "al unui lanț este opus capătului 5" al celuilalt .). Lanțurile formează spirale dreptaci cu o axă comună. O rotație a helixului este de 10 nucleotide, dimensiunea bobinei este de 3,4 nm, înălțimea fiecărui nucleotid este de 0,34 nm, iar diametrul helixului este de 2,0 nm. Ca rezultat al rotației unui fir în jurul celuilalt, se formează o canelură mare (cu diametrul de aproximativ 20 Å) și o canelură mică (aproximativ 12 Å) a dublei spirale ADN. Această formă a dublei spirale Watson-Crick a fost numită ulterior forma B. În celule, ADN-ul există de obicei în forma B, care este cea mai stabilă.
Funcțiile ADN
Modelul propus a explicat multe proprietăți biologice ale acidului dezoxiribonucleic, inclusiv stocarea informațiilor genetice și diversitatea genelor furnizate de o mare varietate de combinații secvențiale de 4 nucleotide și faptul existenței codului genetic, capacitatea de auto-reproducere și să transfere informații genetice furnizate de procesul de replicare și implementarea informațiilor genetice sub formă de proteine, precum și orice alți compuși formați cu ajutorul proteinelor enzimatice.
Funcțiile de bază ale ADN-ului.
- ADN-ul este purtătorul informațiilor genetice, care este asigurat de faptul existenței codului genetic.
- Reproducerea și transmiterea informațiilor genetice în generații de celule și organisme. Această funcționalitate este furnizată de procesul de replicare.
- Realizarea informațiilor genetice sub formă de proteine, precum și a oricăror alți compuși formați cu ajutorul proteinelor enzimatice. Această funcție este asigurată de procesele de transcriere și traducere.

Forme de organizare a ADN bicatenar
ADN-ul poate forma mai multe tipuri de spirale duble (Fig. 4). În prezent, șase forme sunt deja cunoscute (de la A la E și forma Z).
Formele structurale ale ADN-ului, așa cum a stabilit Rosalind Franklin, depind de saturația moleculei de acid nucleic cu apă. În studiile asupra fibrelor ADN care utilizează analiza difracției cu raze X, s-a arătat că modelul de difracție cu raze X depinde radical de la ce umiditate relativă, la ce grad de saturație de apă a acestei fibre are loc experimentul. Dacă fibra era suficient de saturată cu apă, atunci se obținea o raze X. La uscare, a apărut un model cu raze X complet diferit, care era foarte diferit de modelul cu raze X al fibrei cu umiditate ridicată.
Molecula de ADN cu umiditate ridicată se numește formă B... În condiții fiziologice (concentrație scăzută de sare, grad ridicat de hidratare), tipul structural dominant de ADN este forma B (principala formă de ADN dublu catenar este modelul Watson-Crick). Pasul elicei unei astfel de molecule este de 3,4 nm. Există 10 perechi complementare pe rând sub formă de stive răsucite de „monede” - baze azotate. Stivele sunt ținute de legături de hidrogen între două „monede” opuse ale stivei și sunt „înfășurate” în două panglici de schelet fosfodiester răsucite într-o spirală dreaptă. Planurile bazelor azotate sunt perpendiculare pe axa spiralei. Perechile complementare adiacente sunt rotite una față de cealaltă cu 36 °. Diametrul helixului este de 20 Å, nucleotida purinică ocupând 12 Å, iar nucleotida pirimidinică - 8 Å.
Molecula de ADN cu umiditate mai scăzută se numește formă A... Forma A se formează în condiții de hidratare mai puțin ridicată și cu un conținut mai mare de ioni Na + sau K +. Această conformație dreaptă mai largă are 11 perechi de baze pe tură. Planurile bazelor azotate au o înclinație mai puternică spre axa helixului; sunt deviate de la normal la axa helixului cu 20 °. Prin urmare, urmează prezența unui gol intern cu un diametru de 5 Å. Distanța dintre nucleotidele adiacente este de 0,23 nm, lungimea bobinei este de 2,5 nm, iar diametrul helixului este de 2,3 nm.
Inițial, forma A a ADN-ului a fost considerată a fi mai puțin importantă. Cu toate acestea, mai târziu a devenit clar că forma A a ADN-ului, precum și forma B, au o semnificație biologică enormă. O helică ARN-ADN din complexul șablon-primer are o formă A, precum și o helică ARN-ARN și structuri de ac de păr ARN (gruparea 2'-hidroxil a ribozei nu permite moleculelor de ARN să formeze o formă B) . Forma A a ADN-ului a fost găsită în controversă. S-a constatat că forma A a ADN-ului este de 10 ori mai rezistentă la razele UV decât forma B.
Forma A și forma B sunt numite forme canonice ale ADN-ului.
Formulare C-E sunt, de asemenea, dreptaci, formarea lor poate fi observată numai în experimente speciale și, aparent, nu există in vivo. ADN-ul în formă de C are o structură similară cu ADN-ul B. Numărul de perechi de baze pe tură este de 9,33, lungimea elicei este de 3,1 nm. Perechile de baze sunt înclinate la un unghi de 8 grade față de poziția perpendiculară pe axă. Canelurile au dimensiuni similare canelurilor B-ADN. În acest caz, canelura principală este oarecum mai mică, iar canelura minoră este mai profundă. Polinucleotidele ADN naturale și sintetice pot trece în forma C.
| Tabelul 1. Caracteristicile unor tipuri de structuri ADN | |||
| Tipul spiralat | A | B | Z |
| Pas spiralat | 0,32 nm | 3,38 nm | 4,46 nm |
| Răsucire în spirală | Dreapta | Dreapta | Stânga |
| Perechi de bază pe rând | 11 | 10 | 12 |
| Distanța dintre planurile de bază | 0,256 nm | 0,338 nm | 0,371 nm |
| Conformația legăturii glicozidice | anti | anti | anti-C syn-g |
| Conformația inelului furanoză | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-c |
| Lățimea canelurii mici / mari | 1,11 / 0,22 nm | 0,57 / 1,17 nm | 0,2 / 0,88 nm |
| Adâncimea canelurii, mică / mare | 0,26 / 1,30 nm | 0,82 / 0,85 nm | 1,38 / 0,37 nm |
| Diametrul spiralei | 2,3 nm | 2,0 nm | 1,8 nm |
Elemente structurale ale ADN-ului
(structuri ADN necanonice)
Elementele structurale ale ADN-ului includ structuri neobișnuite, limitate de unele secvențe speciale:
|

ADN în formă de Z a fost descoperit în 1979 în timp ce studia hexanucleotida d (CG) 3 -. A fost descoperit de profesorul MIT Alexander Rich și colegii săi. Forma Z a devenit unul dintre cele mai importante elemente structurale ale ADN datorită faptului că formarea sa a fost observată în regiunile ADN în care purinele alternează cu pirimidinele (de exemplu, 5'-HCGCH-3 '), sau în repetări de 5' -CHCH-3 'conținând citozină metilată. O condiție esențială pentru formarea și stabilizarea ADN-ului Z a fost prezența nucleotidelor purinice în conformația sin, alternând cu bazele pirimidinei în anticonformație.
Moleculele naturale de ADN există în principal în forma B corectă, dacă nu conțin secvențe de tip (CH) n. Cu toate acestea, dacă astfel de secvențe fac parte din ADN, atunci aceste regiuni, atunci când se modifică puterea ionică a soluției sau cationii care neutralizează sarcina negativă pe coloana vertebrală a fosfodiesterului, se pot transforma în forma Z, în timp ce alte regiuni ADN din lanț rămân în forma B clasică. Posibilitatea unei astfel de tranziții indică faptul că cele două lanțuri din dubla helică a ADN-ului se află într-o stare dinamică și se pot relaxa unul față de celălalt, trecând de la forma dreaptă la cea stângă și invers. Consecințele biologice ale unei astfel de labilități, care permit transformări conformaționale ale structurii ADN, nu sunt încă pe deplin înțelese. Se crede că regiunile Z-ADN joacă un rol în reglarea expresiei unor gene și sunt implicate în recombinarea genetică.
Forma Z a ADN-ului este o helix dublu stângaci, în care coloana vertebrală fosfodiesterică este situată în zig-zag de-a lungul axei moleculei. De aici și numele moleculei (zigzag) -ADN. ADN-ul Z este cel mai puțin răsucit (12 perechi de baze pe rând) și cel mai subțire cunoscut în natură. Distanța dintre nucleotidele adiacente este de 0,38 nm, lungimea bobinei este de 4,56 nm, iar diametrul ADN-ului Z este de 1,8 nm. In afara de asta, aspect această moleculă de ADN se distinge prin prezența unui canal.
ADN-ul în formă de Z a fost găsit în celulele procariote și eucariote. În prezent, s-au obținut anticorpi care pot distinge forma Z de forma B a ADN-ului. Acești anticorpi se leagă de regiuni specifice ale cromozomilor uriași ai celulelor glandelor salivare ale Drosophila (Dr. melanogaster). Reacția de legare este ușor de urmărit datorită structurii neobișnuite a acestor cromozomi, în care regiunile mai dense (discuri) contrastează cu regiunile mai puțin dense (interdiscuri). Regiunile ADN-ului Z sunt situate în interbande. Rezultă din aceasta că forma Z există de fapt în condiții naturale, deși dimensiunile secțiunilor individuale ale formei Z sunt încă necunoscute.

(shifters) sunt cele mai cunoscute și frecvent găsite secvențe de bază în ADN. Un palindrom este un cuvânt sau o expresie care citește de la stânga la dreapta și invers în același mod. Exemple de astfel de cuvinte sau fraze sunt: SHALASH, KAZAK, POTOP ȘI ROSE TRĂVITE PE LABA AZOR. Atunci când este aplicat regiunilor ADN, acest termen (palindrom) înseamnă aceeași alternanță de nucleotide de-a lungul lanțului de la dreapta la stânga și de la stânga la dreapta (cum ar fi literele din cuvântul „colibă” etc.).
Un palindrom se caracterizează prin prezența repetărilor inversate ale secvențelor de bază având simetrie de ordinul doi în raport cu două catene de ADN. Astfel de secvențe, dintr-un motiv de înțeles, sunt auto-complementare și tind să formeze ac de păr sau structuri cruciforme (Fig.). Agrafe de păr ajută proteinele reglatoare să recunoască locul în care textul genetic al ADN-ului cromozomial este anulat.
În cazurile în care o repetare inversată este prezentă pe aceeași catenă de ADN, această secvență se numește repetare în oglindă. Repetările în oglindă nu posedă proprietățile de auto-complementaritate și, prin urmare, nu sunt capabile să formeze ac de păr sau structuri cruciforme. Secvențele de acest tip se găsesc în aproape toate moleculele mari de ADN și pot varia de la doar câteva perechi de baze la câteva mii de perechi de baze.
Prezența palindromilor sub formă de structuri cruciforme în celulele eucariote nu a fost dovedită, deși s-au găsit o serie de structuri cruciforme in vivo în celulele E. coli. Prezența secvențelor autocomplementare în ARN sau ADN monocatenar este principalul motiv pentru plierea unui lanț nucleic în soluții într-o anumită structură spațială, caracterizată prin formarea multor „agrafe de păr”.

ADN în formă de H este o helix formată din trei fire ADN - tripla helix ADN. Este un complex al dublei spirale Watson-Crick cu a treia catenă ADN monocatenară, care se potrivește în canelura sa mare, cu formarea așa-numitei perechi Hoogsteen.
Formarea unui astfel de triplex are loc ca urmare a plierii dublei spirale ADN în așa fel încât jumătate din secțiunea sa rămâne sub forma unei spirale duble, iar cealaltă jumătate este deconectată. În acest caz, una dintre spiralele deconectate formează o nouă structură cu prima jumătate a spiralei duble - o spirală triplă, iar a doua se dovedește a fi nestructurată, sub forma unei secțiuni monocatenare. O caracteristică a acestei tranziții structurale este o dependență puternică de pH-ul mediului, ai cărui protoni stabilizează noua structură. Datorită acestei caracteristici, noua structură a fost denumită forma H a ADN-ului, a cărui formare a fost găsită în plasmidele supraînfășurate care conțin regiuni homopurină-homopirimidină, care sunt o repetare în oglindă.
În studii ulterioare, posibilitatea unei tranziții structurale a unor polinucleotide bicatenare homopurină-homopirimidină cu formarea unei structuri cu trei catene care conține:
- o homopurină și două fire de homopirimidină ( Py-Pu-Py triplex) [Interacțiunea Hoogsteen].
Blocurile constitutive ale triplexului Py-Pu-Py sunt triade canonice izomorfe la CGC + și TAT. Stabilizarea triplexului necesită protonarea triadei CGC +; prin urmare, aceste triplexuri depind de pH-ul soluției.
- o homopirimidină și două fire de homopurină ( Py-Pu-Pu triplex) [inversează interacțiunea Hoogsteen].
Blocurile constitutive ale triplexului Py-Pu-Pu sunt izomorfe canonice pentru triade CGG și TAA. O proprietate esențială a triplexurilor Py-Pu-Pu este dependența stabilității lor de prezența ionilor dublu încărcați și sunt necesari ioni diferiți pentru stabilizarea triplexurilor de diferite secvențe. Deoarece formarea triplexurilor Py-Pu-Pu nu necesită protonarea nucleotidelor lor constitutive, astfel de triplexuri pot exista la pH neutru.
Notă: interacțiunea directă și inversă Hoogsteen se explică prin simetria 1-metiltiminei: o rotație de 180 ° duce la faptul că atomul O4 este înlocuit cu atomul O2, în timp ce sistemul de legături de hidrogen este păstrat.
Există două tipuri de spirale triple:
- helice triple paralele, în care polaritatea celui de-al treilea fir coincide cu polaritatea lanțului homopurinic al duplexului Watson-Crick
- helice triple antiparalele, în care polaritățile lanțurilor a treia și homopurină sunt opuse.

G-quadruplex- ADN cu 4 catene. O astfel de structură se formează dacă există patru guanine, care formează așa-numitul G-quadruplex - un dans rotund de patru guanine.
Primele indicii cu privire la posibilitatea formării unor astfel de structuri au fost primite cu mult înainte de lucrarea de revoluție a lui Watson și Crick - în 1910. Apoi, chimistul german Ivar Bang a descoperit că una dintre componentele ADN - acidul guanosic - formează geluri la concentrații mari, în timp ce alte componente ale ADN-ului nu au această proprietate.
În 1962, folosind metoda difracției cu raze X, a fost posibilă stabilirea structurii celulare a acestui gel. S-a dovedit a fi compus din patru reziduuri de guanină, legate între ele într-un cerc și formând un pătrat caracteristic. În centru, legătura este susținută de un ion metalic (Na, K, Mg). Aceleași structuri se pot forma în ADN dacă conține multă guanină. Aceste pătrate plate (cvartete G) sunt stivuite pentru a forma structuri destul de stabile, dense (G-quadruplexe).
Patru catene separate de ADN pot fi împletite în complexe cu patru catene, dar aceasta este mai degrabă o excepție. Mai des, o singură catenă de acid nucleic este pur și simplu legată într-un nod, formând îngroșări caracteristice (de exemplu, la capetele cromozomilor), sau ADN-ul catenar dublu formează un quadruplex local într-o regiune bogată în guanină.
Cea mai studiată este existența cvadruplexelor la capetele cromozomilor - pe telomeri și la oncopromotoare. Cu toate acestea, până acum, nu se cunoaște o înțelegere completă a localizării unui astfel de ADN în cromozomii umani.
Toate aceste structuri neobișnuite de ADN sub formă liniară sunt instabile în comparație cu ADN-ul în formă de B. Cu toate acestea, ADN-ul există adesea într-o formă circulară de stres topologic atunci când are ceea ce se numește supraîncărcare. În aceste condiții, structurile ADN necanonice se formează cu ușurință: forme Z, cruci și agrafe de păr, forme H, quadruplexuri de guanină și motiv i.
- Formă supraînfășurată - se observă atunci când este eliberată din nucleul celular fără a deteriora coloana vertebrală pentoză-fosfat. Are forma unor inele închise super-răsucite. În starea supraînfășurată, dubla helică a ADN-ului este „răsucită asupra sa” cel puțin o dată, adică conține cel puțin o super-bobină (ia forma unei cifre de opt).
- Starea relaxată a ADN-ului este observată cu o singură rupere (ruperea unei fire). În acest caz, super-bobinele dispar și ADN-ul ia forma unui inel închis.
- Formă liniară de ADN - observată atunci când două fire ale unei spirale duble sunt rupte.
Structura terțiară a ADN-ului
Structura terțiară a ADN-ului se formează ca rezultat al răsucirii suplimentare în spațiu a unei molecule cu dublă catenă - supraîncărcarea acesteia. Superînfășurarea moleculei de ADN în celulele eucariote, spre deosebire de procariote, se realizează sub formă de complexe cu proteine.
Aproape tot ADN-ul eucariot se găsește în cromozomii nucleelor, doar o cantitate mică din acesta este conținută în mitocondrii, în plante și în plastide. Principala substanță a cromozomilor celulelor eucariote (inclusiv cromozomii umani) este cromatina, formată din ADN bicatenar, histone și proteine non-histonice.
Proteine histonice de cromatină
Histonele sunt proteine simple care alcătuiesc 50% din cromatină. În toate celulele studiate de animale și plante, s-au găsit cinci clase principale de histone: H1, H2A, H2B, H3, H4, diferind ca mărime, compoziția aminoacizilor și valoarea sarcinii (întotdeauna pozitive).
Histona H1 a mamiferelor constă dintr-un singur lanț polipeptidic de aproximativ 215 aminoacizi; dimensiunile altor histone variază de la 100 la 135 de aminoacizi. Toate acestea sunt spiralizate și răsucite într-o globulă cu un diametru de aproximativ 2,5 nm, conțin o cantitate neobișnuit de mare de aminoacizi încărcați pozitiv lizină și arginină. Histonele pot fi acetilate, metilate, fosforilate, poli (ADP) -ribozilate, iar histonele H2A și H2B sunt legate covalent de ubiquitină. Care este rolul unor astfel de modificări în formarea structurii și îndeplinirea funcțiilor de către histone nu a fost încă pe deplin elucidat. Se presupune că aceasta este capacitatea lor de a interacționa cu ADN-ul și de a oferi unul dintre mecanismele de reglare a acțiunii genelor.
Histonele interacționează cu ADN-ul în principal prin legături ionice (punți de sare) formate între grupări fosfat încărcate negativ de ADN și resturi de lizină și arginină încărcate pozitiv de histone.
Proteine cromatine non-histonice
Spre deosebire de histone, proteinele non-histonice sunt foarte diverse. Au fost izolate până la 590 de fracții diferite de proteine non-histonice care se leagă de ADN. Sunt numite și proteine acide, deoarece aminoacizii acizi predomină în structura lor (sunt polianioni). Reglarea specifică a activității cromatinei este asociată cu o varietate de proteine non-histonice. De exemplu, enzimele necesare pentru replicarea și exprimarea ADN se pot lega temporar de cromatină. Alte proteine, de exemplu, care participă la diverse procese de reglare, se leagă de ADN numai în țesuturi specifice sau în anumite etape de diferențiere. Fiecare proteină este complementară unei secvențe specifice de nucleotide ADN (situs ADN). Acest grup include:
- o familie de proteine deget de zinc specifice site-ului. Fiecare deget de zinc recunoaște un site specific format din 5 perechi de nucleotide.
- familie de proteine site-specific - homodimeri. Un fragment dintr-o astfel de proteină în contact cu ADN-ul are o structură elicoidală-elicoidală.
- proteinele gel cu mobilitate ridicată (proteine HMG) sunt un grup de proteine structurale și de reglare care sunt asociate în mod constant cu cromatina. Au o greutate moleculară mai mică de 30 kDa și se caracterizează printr-un conținut ridicat de aminoacizi încărcați. Datorită greutății lor moleculare mici, proteinele HMG sunt extrem de mobile în timpul electroforezei pe gel de poliacrilamidă.
- enzime de replicare, transcriere și reparare.
Cu participarea proteinelor structurale, de reglare și a enzimelor implicate în sinteza ADN-ului și ARN-ului, catena nucleozomică este transformată într-un complex extrem de condensat de proteine și ADN. Structura rezultată este de 10.000 de ori mai scurtă decât molecula originală de ADN.

Cromatina
Cromatina este un complex de proteine cu ADN nuclear și substanțe anorganice... Cea mai mare parte a cromatinei este inactivă. Conține ADN condensat strâns. Este heterocromatină. Distingeți între cromatina constitutivă, inactivă genetic (ADN satelit), formată din regiuni neexprimate, și facultativă - inactivă în mai multe generații, dar în anumite circumstanțe capabile să se exprime.
Cromatina activă (euchromatin) este necondensată, adică ambalate mai puțin strâns. În diferite celule, conținutul său variază de la 2 la 11%. În celulele creierului este cel mai mult - 10-11%, în celulele ficatului - 3-4 și rinichi - 2-3%. Se constată transcrierea activă a euchromatinei. În același timp, organizarea sa structurală permite ca una și aceeași informație genetică ADN inerentă acestui tip de organism să fie folosită în moduri diferite în celule specializate.
Într-un microscop electronic, imaginea cromatinei seamănă cu o margelă: îngroșări sferice de aproximativ 10 nm în mărime, separate prin punți asemănătoare firului. Aceste îngroșări globulare se numesc nucleozomi. Nucleozomul este o unitate structurală a cromatinei. Fiecare nucleozom conține un segment de ADN supraînfășurat cu o lungime de 146 perechi de baze, înfășurat cu formarea de 1,75 viraje la stânga pe nucleul nucleozomului. Nucleul nucleosomal este un octamer histonic format din histone H2A, H2B, H3 și H4, două molecule de fiecare tip (Fig. 9), care arată ca un disc de 11 nm în diametru și 5,7 nm grosime. A cincea histonă, H1, nu face parte din nucleul nucleozomal și nu este implicată în procesul de înfășurare a ADN-ului pe octamerul histonei. Contactează ADN-ul unde dubla helix intră și iese din nucleul nucleozomului. Acestea sunt regiuni ADN intercorticale (linker), a căror lungime variază în funcție de tipul de celulă de la 40 la 50 de perechi de nucleotide. Ca urmare, lungimea fragmentului de ADN inclus în nucleozom variază de asemenea (de la 186 la 196 perechi de nucleotide).
Nucleozomul conține aproximativ 90% din ADN, restul acestuia este linkerul. Se crede că nucleozomii sunt fragmente de cromatină „tăcută”, iar linkerul este activ. Cu toate acestea, nucleozomii se pot desfășura și deveni liniari. Nucleozomii desfășurați sunt deja cromatină activă. Astfel se manifestă în mod clar dependența funcției de structură. Se poate presupune că cu cât este mai multă cromatină în compoziția nucleozomilor globulari, cu atât este mai puțin activă. Evident, în diferite celule, proporția inegală de cromatină în repaus este asociată cu numărul de astfel de nucleozomi.
Pe fotografiile microscopice electronice, în funcție de condițiile de izolare și de gradul de întindere, cromatina poate arăta nu numai ca un fir lung cu îngroșări - „margele” de nucleozomi, ci și ca o fibrilă (fibră) mai scurtă și mai densă cu un diametru de 30 nm, a cărui formare este observată în timpul interacțiunii histonei H1 legată de regiunea linker a ADN-ului și histonei H3, ceea ce duce la răsucirea suplimentară a unei spirale de șase nucleozomi pe rând cu formarea unui solenoid cu diametrul de 30 nm. În acest caz, o proteină histonică poate interfera cu transcrierea unui număr de gene și astfel poate regla activitatea acestora.
Ca rezultat al interacțiunilor descrise mai sus ale ADN-ului cu histone, un segment al dublei spirale ADN de 186 perechi de baze cu un diametru mediu de 2 nm și o lungime de 57 nm se transformă într-o spirală cu un diametru de 10 nm și o lungime de 5 nm. Comprimarea ulterioară a acestei spirale la o fibră cu un diametru de 30 nm crește gradul de condensare cu un factor de șase.
În cele din urmă, împachetarea unui duplex de ADN cu cinci histone are ca rezultat o condensare de 50 de ori a ADN-ului. Cu toate acestea, chiar și un grad atât de ridicat de condensare nu poate explica densificarea de aproape 50.000 - 100.000 de ori a ADN-ului în cromozomul metafazic. Din păcate, detaliile privind ambalarea ulterioară a cromatinei până la cromozomul metafazic nu sunt încă cunoscute; prin urmare, numai caracteristicile generale ale acestui proces pot fi luate în considerare.
Nivelurile de compactare a ADN-ului în cromozomi
Fiecare moleculă de ADN este ambalată într-un cromozom separat. Celulele diploide umane conțin 46 de cromozomi, care sunt localizați în nucleul celular. Lungimea totală a ADN-ului tuturor cromozomilor unei celule este de 1,74 m, dar diametrul nucleului în care sunt ambalate cromozomii este de milioane de ori mai mic. O astfel de împachetare compactă de ADN în cromozomi și cromozomi din nucleul celulei este asigurată de o varietate de proteine histone și non-histonice care interacționează într-o secvență specifică cu ADN (vezi mai sus). Compactarea ADN-ului în cromozomi face posibilă reducerea dimensiunilor sale liniare de aproximativ 10.000 de ori - în mod convențional de la 5 cm la 5 microni. Există mai multe niveluri de compactare (Fig. 10).


- ADN dublu helix este o moleculă încărcată negativ cu un diametru de 2 nm și o lungime de câțiva cm.
- nivel nucleosomal- cromatina arată într-un microscop electronic ca un lanț de „margele” - nucleozomi - „pe un șir”. Nucleozomul este o unitate structurală universală care se găsește atât în eucromatină, cât și în heterocromatină, în nucleul interfazic și în cromozomii metafazici.
Nivelul nucleozomal de compactare este asigurat de proteine speciale - histone. Opt domenii de histone încărcate pozitiv formează nucleul (nucleul) nucleozomului în jurul căruia este înfășurată molecula de ADN încărcat negativ. Aceasta are ca rezultat o scurtare de 7 ori, în timp ce diametrul crește de la 2 la 11 nm.
- nivelul solenoidului
Nivelul solenoidului de organizare a cromozomilor se caracterizează prin răsucirea filamentului nucleozomal și formarea fibrilelor mai groase cu diametrul de 20-35 nm din acesta - solenoizi sau superbide. Pasul solenoidului este de 11 nm; există aproximativ 6-10 nucleozomi pe tură. Ambalarea solenoidelor este considerată mai probabilă decât superbidă, conform căreia o fibrilă de cromatină cu diametrul de 20-35 nm este un lanț de granule sau superbide, fiecare dintre ele constând din opt nucleozomi. La nivelul solenoidului, dimensiunea liniară a ADN-ului este redusă de 6-10 ori, diametrul crește la 30 nm.
- nivelul buclei
Nivelul buclei este asigurat de proteine care nu se leagă de ADN-ul specific situsului histonei, care recunosc și se leagă de secvențe specifice de ADN, formând bucle de aproximativ 30-300 kb. Bucla oferă expresie genică, adică E. bucla nu este doar o formațiune structurală, ci și funcțională. Scurtarea la acest nivel are loc de 20-30 de ori. Diametrul crește la 300 nm. Structurile de tip buclă de tip „lamp-brush” la ovocitele amfibiene pot fi văzute pe preparatele citologice. Aceste bucle, aparent, sunt supraînfășurate și reprezintă domenii ADN care corespund probabil unităților de transcripție și replicare a cromatinei. Proteinele specifice fixează bazele buclelor și, eventual, unele dintre regiunile lor interioare. Organizarea domeniului în formă de buclă promovează plierea cromatinei în cromozomii metafazici în structuri elicoidale de ordine superioare.
- la nivel de domeniu
Nivelul domeniului de organizare a cromozomilor a fost insuficient studiat. La acest nivel, se constată formarea domeniilor de buclă - structurile firelor (fibrilelor) cu grosimea de 25-30 nm, care conțin 60% proteine, 35% ADN și 5% ARN, sunt practic invizibile în toate fazele celulei ciclu cu excepția mitozei și sunt oarecum distribuite aleator peste nucleul celular. Structurile de tip buclă de tip „lamp-brush” la ovocitele amfibiene pot fi văzute pe preparatele citologice.
Domeniile de buclă cu baza lor sunt atașate la matricea de proteine intranucleare în așa-numitele site-uri de atașament încorporate, adesea denumite secvențe MAR / SAR (MAR, din regiunea engleză asociată cu matricea; SAR, din regiunile de atașare a schelelor engleze) - ADN fragmente de câteva sute de perechi de baze în lungime, care se caracterizează printr-un conținut ridicat (> 65%) perechi de baze A / T. Fiecare domeniu pare să aibă o singură origine a replicării și funcționează ca o unitate superînfășurată autonomă. Orice domeniu în buclă conține multe unități de transcriere, a căror funcționare este probabil coordonată - întregul domeniu este fie într-o stare activă, fie inactivă.
La nivel de domeniu, ca rezultat al împachetării secvențiale a cromatinei, dimensiunile liniare ale ADN scad de aproximativ 200 de ori (700 nm).
- nivel cromozomial
La nivel cromozomial, condensarea cromozomului profazic în cromozomul metafazic are loc cu compactarea domeniilor buclei în jurul cadrului axial al proteinelor non-histonice. Această superînfășurare este însoțită de fosforilarea tuturor moleculelor H1 din celulă. Ca rezultat, cromozomul metafazic poate fi descris ca niște bucle de solenoid strâns înfășurate într-o spirală strânsă. Un cromozom uman tipic poate conține până la 2600 de bucle. Grosimea unei astfel de structuri atinge 1400 nm (două cromatide), în timp ce molecula ADN este scurtată de 104 ori, adică cu 5 cm de ADN întins la 5 μm.
Funcții cromozomiale
În interacțiunea cu mecanismele extracromozomale, cromozomii asigură
- stocarea informațiilor ereditare
- folosind aceste informații pentru a crea și a menține organizarea celulară
- reglementarea citirii informațiilor ereditare
- autodublarea materialului genetic
- transferul materialului genetic de la celula mamă la fiică.
Există dovezi că atunci când regiunea cromatinei este activată, adică în timpul transcrierii, histona H1 este îndepărtată reversibil din ea mai întâi, apoi octetul histonei. Acest lucru determină decondensarea cromatinei, o tranziție secvențială a unei fibrile de cromatină de 30 nanometri într-un filament de 10 nanometri și desfășurarea acesteia în regiuni de ADN liber, adică pierderea structurii nucleozomale.
Știm cu toții că aspectul unei persoane, unele obiceiuri și chiar boli sunt moștenite. Toate aceste informații despre o ființă vie sunt codificate în gene. Deci, cum arată aceste gene notorii, cum funcționează și unde sunt localizate?
Deci, purtătorul tuturor genelor oricărei persoane sau animale este ADN-ul. Acest compus a fost descoperit în 1869 de Johann Friedrich Miescher. Din punct de vedere chimic, ADN-ul este acid dezoxiribonucleic. Ce inseamna asta? Cum poartă acest acid codul genetic al vieții de pe planeta noastră?
Să începem prin a privi unde se află ADN-ul. Într-o celulă umană, există multe organite care îndeplinesc diverse funcții. ADN-ul este localizat în nucleu. Nucleul este un mic organet care este înconjurat de o membrană specială și în care este stocat tot materialul genetic - ADN.
Care este structura unei molecule de ADN?
În primul rând, să ne uităm la ce este ADN-ul. ADN-ul este o moleculă foarte lungă formată din blocuri de construcție - nucleotide. Există 4 tipuri de nucleotide - adenină (A), timină (T), guanină (G) și citozină (C). Lanțul nucleotidic arată schematic astfel: GGAATCTAAG ... Aceasta este secvența de nucleotide care este lanțul ADN.Structura ADN-ului a fost descifrată pentru prima dată în 1953 de James Watson și Francis Crick.
Într-o moleculă de ADN, există două lanțuri de nucleotide care sunt răsucite elicoidal unul în jurul celuilalt. Cum se lipesc aceste lanțuri de nucleotide și se răsucesc într-o spirală? Acest fenomen se datorează proprietății complementarității. Complementaritatea înseamnă că numai anumite nucleotide (complementare) pot fi localizate una față de cealaltă în două catene. Deci, opus adeninei există întotdeauna timină, iar opus guaninei există întotdeauna doar citozină. Astfel, guanina este complementară citozinei, iar adenina este complementară timinei. Astfel de perechi de nucleotide care se confruntă unul cu celălalt în diferite fire sunt numite și complementare.
 Poate fi descris schematic după cum urmează:
Poate fi descris schematic după cum urmează:
G - C
T - A
T - A
C - G
Aceste perechi complementare A - T și G - C formează legătură chimicăîntre nucleotidele perechii, iar legătura dintre G și C este mai puternică decât între A și T. Legătura se formează strict între baze complementare, adică formarea unei legături între G și A necomplementare este imposibilă.
Ambalarea ADN-ului, cum devine un fir ADN un cromozom?
De ce aceste lanțuri de nucleotide ADN se răsucesc, de asemenea, unul în jurul celuilalt? De ce este nevoie de asta? Faptul este că numărul de nucleotide este enorm și este nevoie de mult spațiu pentru a găzdui lanțuri atât de lungi. Din acest motiv, există o răsucire în spirală a două fire ADN în jurul celuilalt. Acest fenomen se numește spiralizare. Ca urmare a spiralizării, firele de ADN sunt scurtate de 5-6 ori.Unele molecule de ADN sunt utilizate în mod activ de către organism, în timp ce altele sunt rareori folosite. Astfel de molecule de ADN utilizate rar, pe lângă spiralizare, suferă o „împachetare” și mai compactă. Acest pachet compact se numește superînfășurare și scurtează firul ADN de 25-30 de ori!
Cum are loc ambalarea catenelor de ADN?
 Pentru supraînfășurare, se utilizează proteine histonice, care au aspectul și structura unei tije sau a unei bobine cu fir. Catenele de ADN spiralizate sunt înfășurate pe aceste „bobine” - proteine histonice. Astfel, firul lung devine foarte compact și ocupă foarte puțin spațiu.
Pentru supraînfășurare, se utilizează proteine histonice, care au aspectul și structura unei tije sau a unei bobine cu fir. Catenele de ADN spiralizate sunt înfășurate pe aceste „bobine” - proteine histonice. Astfel, firul lung devine foarte compact și ocupă foarte puțin spațiu. Dacă este necesar să se utilizeze una sau alta moleculă de ADN, are loc procesul de „desfacere”, adică firul de ADN este „desfăcut” din „bobină” - o proteină histonică (dacă a fost înfășurată pe ea) și desfăcută din o spirală în două lanțuri paralele. Și când molecula de ADN se află într-o stare atât de neîntoarsă, din ea pot fi citite informațiile genetice necesare. Mai mult decât atât, citirea informațiilor genetice are loc numai din șiruri de ADN neîntoarse!
Se numește setul de cromozomi supraînfășurați heterocromatină, și cromozomii disponibili pentru citirea informațiilor - euchromatin.
Ce sunt genele, care este relația lor cu ADN-ul?
Acum să ne uităm la ce sunt genele. Se știe că există gene care determină tipul de sânge, culoarea ochilor, părul, pielea și multe alte proprietăți ale corpului nostru. O genă este o secțiune strict definită a ADN-ului, constând dintr-un anumit număr de nucleotide situate într-o combinație strict definită. Localizarea într-o zonă strict definită a ADN înseamnă că unei gene specifice i s-a atribuit locul și este imposibil să se schimbe acest loc. Este potrivit să se facă o astfel de comparație: o persoană locuiește pe o anumită stradă, într-o anumită casă și apartament, iar o persoană nu se poate muta în mod arbitrar în altă casă, apartament sau pe altă stradă. Un anumit număr de nucleotide dintr-o genă înseamnă că fiecare genă are un număr specific de nucleotide și nu poate deveni mai mult sau mai puțin. De exemplu, gena care codifică producția de insulină are o lungime de 60 de perechi de baze; gena care codifică producția hormonului oxitocină - a 370 perechi de baze. Secvența strictă de nucleotide este unică pentru fiecare genă și este strict definită. De exemplu, secvența AATTAATA este un fragment al unei gene care codifică producția de insulină. Pentru a obține insulină, se folosește doar o astfel de secvență; pentru a obține, de exemplu, adrenalină, se utilizează o combinație diferită de nucleotide. Este important să înțelegem că doar o anumită combinație de nucleotide codifică un anumit „produs” (adrenalină, insulină etc.). O astfel de combinație unică a unui anumit număr de nucleotide, care se află în „locul său” - acesta este genă.
În plus față de gene, așa-numitele „secvențe necodificate” sunt localizate în lanțul ADN. Astfel de secvențe de nucleotide necodificatoare reglează activitatea genelor, ajută la spiralizarea cromozomilor și marchează începutul și sfârșitul unei gene. Cu toate acestea, până în prezent, rolul majorității secvențelor necodificate rămâne neclar.
Ce este un cromozom? Cromozomi sexuali
Colecția genelor unui individ se numește genom. În mod natural, este imposibil să se încadreze întregul genom într-un singur ADN. Genomul este descompus în 46 de perechi de molecule de ADN. O pereche de molecule de ADN se numește cromozom. Deci, acești cromozomi au o persoană care are 46 de bucăți. Fiecare cromozom poartă un set strict definit de gene, de exemplu, cromozomul 18 conține gene care codifică culoarea ochilor etc. Cromozomii diferă între ei prin lungime și formă. Cele mai frecvente forme sunt X sau Y, dar există și altele. O persoană are doi cromozomi de aceeași formă, care se numesc împerecheați (perechi). Datorită unor astfel de diferențe, toți cromozomii asociați sunt numerotați - există 23 de perechi. Aceasta înseamnă că există o pereche de cromozomi # 1, pereche # 2, # 3 etc. Fiecare genă responsabilă de o anumită trăsătură este localizată pe același cromozom. În manualele moderne pentru specialiști, localizarea genei poate fi indicată, de exemplu, după cum urmează: 22 cromozom, braț lung.Care sunt diferențele dintre cromozomi?
 În ce fel diferă cromozomii? Ce înseamnă termenul umăr lung? Să luăm cromozomi de forma X. Intersecția firelor de ADN poate avea loc strict în mijloc (X) sau poate apărea nu central. Când o astfel de intersecție a firelor de ADN nu are loc central, atunci relativ la punctul de trecere, unele capete sunt mai lungi, altele, respectiv, sunt mai scurte. Astfel de capete lungi sunt denumite de obicei brațul lung al cromozomului, iar cele scurte, respectiv, sunt numite brațul scurt. În cromozomii în formă de Y, umerii lungi ocupă majoritatea, iar cei scurți sunt foarte mici (nici măcar nu sunt indicați în imaginea schematică).
În ce fel diferă cromozomii? Ce înseamnă termenul umăr lung? Să luăm cromozomi de forma X. Intersecția firelor de ADN poate avea loc strict în mijloc (X) sau poate apărea nu central. Când o astfel de intersecție a firelor de ADN nu are loc central, atunci relativ la punctul de trecere, unele capete sunt mai lungi, altele, respectiv, sunt mai scurte. Astfel de capete lungi sunt denumite de obicei brațul lung al cromozomului, iar cele scurte, respectiv, sunt numite brațul scurt. În cromozomii în formă de Y, umerii lungi ocupă majoritatea, iar cei scurți sunt foarte mici (nici măcar nu sunt indicați în imaginea schematică). Mărimea cromozomilor variază: cei mai mari sunt cromozomii de perechi # 1 și # 3, cei mai mici sunt cromozomi de perechi # 17, # 19.
În plus față de formă și dimensiune, cromozomii diferă prin funcțiile lor. Din cele 23 de cupluri, 22 sunt somatice și 1 sunt sexuale. Ce înseamnă? Cromozomii somatici determină toate semnele externe ale unui individ, în special al acestuia răspunsuri comportamentale, psihotipul ereditar, adică toate trăsăturile și caracteristicile fiecărei persoane individuale. O pereche de cromozomi sexuali determină sexul unei persoane: bărbat sau femeie. Există două tipuri de cromozomi sexuali umani - X (X) și Y (Y). Dacă sunt combinate ca XX (X - X) - aceasta este o femeie, iar dacă XY (X - Y) - avem un bărbat.
Boli ereditare și leziuni cromozomiale
Cu toate acestea, apar „defecțiuni” ale genomului, apoi bolile genetice sunt detectate la oameni. De exemplu, când există trei cromozomi pe 21 de perechi de cromozomi în loc de doi, se naște o persoană cu sindrom Down.Există multe „defecțiuni” mai mici ale materialului genetic care nu duc la apariția bolii, ci, dimpotrivă, conferă proprietăți bune. Toate „defecțiunile” materialului genetic sunt numite mutații. Mutațiile care duc la boli sau deteriorarea proprietăților organismului sunt considerate negative, iar mutațiile care duc la formarea de noi proprietăți benefice sunt considerate pozitive.
Cu toate acestea, în raport cu majoritatea bolilor pe care oamenii le suferă astăzi, nu este o boală moștenită, ci doar o predispoziție. De exemplu, tatăl unui copil asimilează zahărul încet. Aceasta nu înseamnă că copilul se va naște cu diabet, dar copilul va avea o predispoziție. Aceasta înseamnă că, dacă un copil abuzează de dulciuri și produse din făină, va dezvolta diabet zaharat.
Astăzi, așa-numitul predicativ medicament. În cadrul acestei practici medicale, predispozițiile sunt identificate într-o persoană (pe baza identificării genelor corespunzătoare) și apoi i se oferă recomandări - ce dietă să urmeze, cum să alterneze corect modul de lucru și odihnă, astfel încât să nu se îmbolnăvească.
Cum se citesc informații codificate în ADN?
Cum puteți citi informațiile conținute în ADN? Cum îl folosește propriul ei corp? ADN-ul în sine este un fel de matrice, dar nu simplu, ci codificat. Pentru a citi informații din matricea ADN, acestea sunt mai întâi transferate către un purtător special - ARN. ARN-ul este acid ribonucleic chimic. Se diferențiază de ADN prin faptul că poate trece prin membrana nucleară în celulă, iar ADN-ul este privat de această capacitate (poate fi doar în nucleu). Informațiile codificate sunt utilizate în celula în sine. Deci, ARN este purtătorul informațiilor codificate de la nucleu la celulă.Cum se sintetizează ARN, cum se sintetizează proteinele cu ajutorul ARN?
 Catenele ADN, din care este necesar să „citim” informațiile, să ne relaxăm, o enzimă specială - „constructor” se apropie de ele și sintetizează o catenă complementară de ARN în paralel cu catenă ADN. Molecula de ARN este, de asemenea, formată din 4 tipuri de nucleotide - adenină (A), uracil (Y), guanină (G) și citozină (C). În acest caz, următoarele perechi sunt complementare: adenină - uracil, guanină - citozină. După cum puteți vedea, spre deosebire de ADN, ARN-ul folosește uracil în loc de timină. Adică, enzima „constructor” funcționează după cum urmează: dacă vede A în catena ADN, atunci atașează Y la catena ARN, dacă G, apoi atașează C etc. Astfel, din fiecare genă activă în timpul transcrierii, se formează un șablon - o copie a ARN-ului care poate trece prin membrana nucleară.
Catenele ADN, din care este necesar să „citim” informațiile, să ne relaxăm, o enzimă specială - „constructor” se apropie de ele și sintetizează o catenă complementară de ARN în paralel cu catenă ADN. Molecula de ARN este, de asemenea, formată din 4 tipuri de nucleotide - adenină (A), uracil (Y), guanină (G) și citozină (C). În acest caz, următoarele perechi sunt complementare: adenină - uracil, guanină - citozină. După cum puteți vedea, spre deosebire de ADN, ARN-ul folosește uracil în loc de timină. Adică, enzima „constructor” funcționează după cum urmează: dacă vede A în catena ADN, atunci atașează Y la catena ARN, dacă G, apoi atașează C etc. Astfel, din fiecare genă activă în timpul transcrierii, se formează un șablon - o copie a ARN-ului care poate trece prin membrana nucleară. Cum se produce sinteza unei proteine codificate de o genă specifică?
După părăsirea nucleului, ARN intră în citoplasmă. Deja în citoplasmă, ARN-ul poate fi, ca matrice, încorporat în sisteme enzimatice speciale (ribozomi), care pot sintetiza, ghidat de informațiile ARN, secvența de aminoacizi proteică corespunzătoare. După cum știți, o moleculă proteică este compusă din aminoacizi. Cum reușește ribozomul să afle ce aminoacizi trebuie atașați lanțului proteic în creștere? Acest lucru se face pe baza unui cod triplet. Codul triplet înseamnă că secvența a trei nucleotide ale lanțului ARN ( triplet, de exemplu, HGH) codifică un aminoacid (în acest caz, glicină). Fiecare aminoacid este codificat de un triplet specific. Și astfel, ribozomul „citește” tripletul, determină ce aminoacid ar trebui atașat în continuare, pe măsură ce citește informații din ARN. Atunci când se formează un lanț de aminoacizi, acesta ia o anumită formă spațială și devine o proteină capabilă să îndeplinească funcțiile enzimatice, de construcție, hormonale și alte funcții care îi sunt atribuite.Proteina pentru orice organism viu este produsul unei gene. Proteinele determină toate diferitele proprietăți, calități și manifestări externe ale genelor.
În dreapta este cea mai mare spirală de ADN uman, construită din oameni pe plaja din Varna (Bulgaria), a intrat în Cartea Recordurilor Guinness pe 23 aprilie 2016
Acidul dezoxiribonucleic. Informații generale
ADN-ul (acidul dezoxiribonucleic) este un fel de plan pentru viață, un cod complex care conține date despre informații ereditare. Această macromoleculă complexă este capabilă să stocheze și să transmită informații genetice ereditare de la generație la generație. ADN determină astfel de proprietăți ale oricărui organism viu, cum ar fi ereditatea și variabilitatea. Informațiile codate în acesta setează întregul program pentru dezvoltarea oricărui organism viu. Factorii inerenți genetic predetermină întregul curs al vieții atât a unei persoane, cât și a oricărui alt organism. Efectele artificiale sau naturale ale mediului extern pot afecta ușor gravitatea generală a trăsăturilor genetice individuale sau pot afecta dezvoltarea proceselor programate.
Acidul dezoxiribonucleic(ADN) este o macromoleculă (una dintre cele trei principale, celelalte două sunt ARN și proteine), care asigură stocarea, transmiterea din generație în generație și implementarea programului genetic pentru dezvoltarea și funcționarea organismelor vii. ADN-ul conține informații despre structura diferitelor tipuri de ARN și proteine.
În celulele eucariote (animale, plante și ciuperci), ADN-ul se găsește în nucleul celulei ca parte a cromozomilor, precum și în unele organite celulare (mitocondrii și plastide). În celulele organismelor procariote (bacterii și arhee), o moleculă circulară sau liniară de ADN, așa-numitul nucleoid, este atașată din interior la membrana celulară. Ei și eucariotele inferioare (de exemplu, drojdia) au, de asemenea, mici molecule de ADN mici, predominant circulare, numite plasmide.
Din punct de vedere chimic, ADN-ul este o moleculă lungă de polimer compusă din blocuri repetate - nucleotide. Fiecare nucleotidă este compusă dintr-o bază azotată, un zahăr (dezoxiriboză) și o grupare fosfat. Legăturile dintre nucleotidele din lanț se formează datorită dezoxiribozei ( CU) și fosfat ( F) grupuri (legături fosfodiester).

Orez. 2. Nuclertidul este format dintr-o bază azotată, zahăr (dezoxiriboză) și o grupare fosfat
În majoritatea covârșitoare a cazurilor (cu excepția unor virusuri care conțin ADN monocatenar), o macromoleculă de ADN constă din două lanțuri orientate între ele de baze azotate. Această moleculă dublu catenară este răsucită într-o linie elicoidală.
Există patru tipuri de baze azotate în ADN (adenină, guanină, timină și citozină). Bazele azotate ale unuia dintre lanțuri sunt conectate cu bazele azotate ale celuilalt lanț prin legături de hidrogen conform principiului complementarității: adenina este conectată numai cu timina ( LA), guanină - numai cu citozină ( G-C). Aceste perechi alcătuiesc „barele transversale” ale „scării” în spirală a ADN-ului (vezi: Fig. 2, 3 și 4).

Orez. 2. Bazele azotate
Secvența de nucleotide vă permite să „codificați” informații despre tipuri diferite ARN-urile, dintre care cele mai importante sunt informaționale, sau mesager (ARNm), ribozomal (ARNr) și transport (ARNt). Toate aceste tipuri de ARN sunt sintetizate pe șablonul ADN prin copierea secvenței ADN în secvența ARN sintetizată în timpul procesului de transcriere și sunt implicate în biosinteza proteinelor (procesul de traducere). Pe lângă secvențele de codificare, ADN-ul celular conține secvențe care îndeplinesc funcții de reglare și structurale.

Orez. 3. Replicarea ADN-ului
Localizarea combinațiilor de bază ale compușilor ADN chimici și relațiile cantitative dintre aceste combinații asigură codificarea informațiilor ereditare.
Educaţie ADN nou (replicare)
- Proces de replicare: desfacerea dublei spirale ADN - sinteza catenelor complementare prin ADN polimerază - formarea a două molecule ADN dintr-una.
- Elica dublă „se dezlănțuie” în două ramuri atunci când enzimele rup legătura dintre perechile de baze ale compușilor chimici.
- Fiecare ramură este un element al ADN-ului nou. Noile perechi de baze sunt conectate în aceeași succesiune ca în ramura părinte.
La finalizarea duplicării, se formează două spirale independente, create din compuși chimici ai ADN-ului parental și având același cod genetic cu acesta. În acest fel, ADN-ul este capabil să sape prin informații de la celulă la celulă.
Informații mai detaliate:
STRUCTURA ACIZILOR NUCLEICI

Orez. 4. Baze azotate: adenină, guanină, citozină, timină
Acidul dezoxiribonucleic(ADN) se referă la acizi nucleici. Acizi nucleici este o clasă de biopolimeri neregulați, ai căror monomeri sunt nucleotide.
NUCLEOTIDE constau din baza azotata combinat cu un carbohidrat cu cinci carbon (pentoză) - dezoxiriboză(în cazul ADN-ului) sau riboză(în cazul ARN), care se combină cu reziduul de acid fosforic (H 2 PO 3 -).
Bazele azotate există două tipuri: baze pirimidinice - uracil (numai în ARN), citozină și timină, baze purinice - adenină și guanină.

Orez. 5. Structura nucleotidelor (stânga), localizarea nucleotidei în ADN (jos) și tipurile de baze azotate (dreapta): pirimidină și purină

Atomii de carbon din molecula de pentoză sunt numerotați de la 1 la 5. Fosfatul se combină cu al treilea și al cincilea atom de carbon. Acesta este modul în care nucleotidele se combină pentru a forma un lanț de acid nucleic. Astfel, putem izola capetele 3 'și 5' ale catenei ADN:

Orez. 6. Izolarea capetelor 3 'și 5' ale catenei ADN
Se formează două catene de ADN helix dublu... Aceste lanțuri într-o spirală sunt orientate în direcții opuse. În diferite fire ADN, bazele azotate sunt interconectate de legături de hidrogen... Adenina se combină întotdeauna cu timina, iar citozina cu guanina. Se numeste regula complementarității(cm. principiul complementarității).
Regula complementarității:
| A-T G-C |
De exemplu, dacă ni se dă o catenă ADN cu secvența
3'- ATGTCCTAGCTGCTCG - 5 ',
atunci al doilea lanț va fi complementar acestuia și direcționat în direcția opusă - de la capătul 5 'până la capătul 3':
5'-TACAGGATCGACGAGC-3 '.

Orez. 7. Direcția lanțurilor moleculei de ADN și conexiunea bazelor azotate utilizând legături de hidrogen
REPLICAȚIE ADN
Replicarea ADN-ului este procesul de dublare a unei molecule de ADN prin sinteza matricială. În majoritatea cazurilor de replicare naturală a ADN-uluigrundcăci sinteza ADN este fragment scurt (recreat). Un astfel de primer ribonucleotidic este creat de enzima primază (ADN primază în procariote, ADN polimerază în eucariote) și este ulterior înlocuit cu deoxiribonucleotide polimerază, care îndeplinește în mod normal funcții de reparare (corectarea daunelor chimice și rupturile moleculei ADN).
Replicarea are loc printr-un mecanism semi-conservator. Aceasta înseamnă că dubla spirală a ADN-ului se desfășoară și o nouă catena este completată pe fiecare dintre catena sa conform principiului complementarității. Prin urmare, molecula ADN fiică conține un lanț din molecula părinte și unul nou sintetizat. Replicarea are loc în direcția de la capătul 3 'la capătul 5' al lanțului părinte.

Orez. 8. Replicarea (dublarea) moleculei ADN
Sinteza ADN-ului- acesta nu este un proces atât de complicat pe cât ar putea părea la prima vedere. Dacă vă gândiți la asta, atunci mai întâi trebuie să vă dați seama ce este sinteza. Este procesul de a aduce ceva împreună. Formarea unei noi molecule de ADN are loc în mai multe etape:
1) ADN topoizomeraza, situată în fața furcii de replicare, taie ADN-ul pentru a facilita desfacerea și desfacerea acestuia.
2) ADN helicaza, în urma topoizomerazei, influențează procesul de „desfacere” a helixului ADN.
3) Proteinele care leagă ADN-ul realizează legarea firelor de ADN și, de asemenea, realizează stabilizarea lor, împiedicându-le să se lipească una de cealaltă.
4) ADN polimerază δ(delta) , coordonată cu viteza de mișcare a furcii replicative, realizează sintezaconducerelanţuri filială ADN în direcția 5 "→ 3" de pe șablon matern Catenă ADN în direcția de la capătul său de 3 "la 5" (viteza de până la 100 de perechi de baze pe secundă). Aceste evenimente despre asta matern Catenele ADN sunt limitate.

Orez. 9. Reprezentarea schematică a procesului de replicare a ADN-ului: (1) Catenă laging (catenă lagging), (2) Catenă principală (catena principală), (3) ADN polimerază α (Polα), (4) ADN ligază, (5) ARN -primer, (6) Primază, (7) fragment Okazaki, (8) ADN polimerază δ (Polδ), (9) Helicază, (10) Proteine monocatenare de legare a ADN, (11) Topoisomerază.
În cele ce urmează este descrisă sinteza firului întârziat al ADN-ului fiică (vezi. Sistem funcție de furcă de replicare și enzimă de replicare)
Pentru o explicație mai vizuală a replicării ADN, vezi
5) Imediat după desfacerea și stabilizarea unui alt fir al moleculei părinte,ADN polimerază α(alfa)iar în direcția 5 "→ 3" sintetizează un primer (primer ARN) - o secvență ARN pe un șablon ADN cu lungimea de 10 până la 200 nucleotide. După aceea, enzimaeste îndepărtat din catena de ADN.
In loc de ADN polimerazăα
se atașează la capătul de 3 "al grundului ADN polimerazăε
.
6)
ADN polimerazăε
(epsilon) ca și cum ar continua să prelungească grundul, dar ca substrat se încorporeazădezoxiribonucleotide(în cantitate de 150-200 nucleotide). Rezultatul este un fir solid din două piese -ARN(adică grund) și ADN.
ADN polimerază εfuncționează până când îndeplinește grundul anteriorfragment de Okazaki(sintetizat puțin mai devreme). Această enzimă este apoi îndepărtată din lanț.
7) ADN polimerază β(beta) se ridică în schimbADN polimerază ε,se mișcă în aceeași direcție (5 "→ 3") și îndepărtează ribonucleotidele de grund, în timp ce introduce dezoxiribonucleotide în locul lor. Enzima funcționează până la îndepărtarea completă a primerului, adică până la o dezoxiribonucleotidă (chiar mai devreme sintetizatăADN polimerază ε). Enzima nu este capabilă să conecteze rezultatul muncii sale și ADN-ul din față, așa că părăsește lanțul.
Drept urmare, un fragment de ADN fiică „zace” pe matricea firului mamei. Se numestefragment de Okazaki.
8) ADN ligaza cusături două adiacente fragmente de Okazaki , adică 5 "-end a segmentului sintetizatADN polimerază ε,și capătul circuitului de 3 ", încorporatADN polimerazăβ .
STRUCTURA ARN
Acid ribonucleic(ARN) este una dintre cele trei macromolecule principale (celelalte două sunt ADN și proteine) care se găsesc în celulele tuturor organismelor vii.
La fel ca ADN-ul, ARN-ul este alcătuit dintr-un lanț lung în care se numește fiecare verigă nucleotidă... Fiecare nucleotidă este compusă dintr-o bază azotată, un zahăr riboză și o grupare fosfat. Cu toate acestea, spre deosebire de ADN, ARN-ul nu are de obicei două fire, ci una. Pentozele din ARN sunt reprezentate de riboză, nu de deoxiriboză (riboză are o grupare hidroxil suplimentară pe al doilea atom de carbohidrați). În cele din urmă, ADN-ul diferă de ARN în compoziția bazelor azotate: în loc de timină ( T) uracil ( U) care este, de asemenea, complementară adeninei.
Secvența de nucleotide permite ARN-ului să codifice informații genetice. Toate organismele celulare folosesc ARN (ARNm) pentru a programa sinteza proteinelor.
ARN-urile celulare sunt realizate printr-un proces numit transcriere , adică sinteza ARN-ului pe matricea ADN, realizată de enzime speciale - ARN polimeraze.
Apoi, ARN-urile messenger (ARNm) participă la un proces numit difuzare, acestea. sinteza proteinelor pe matricea ARNm cu participarea ribozomilor. Alți ARN, după transcripție, suferă modificări chimice, iar după formarea structurilor secundare și terțiare, îndeplinesc funcții în funcție de tipul de ARN.

Orez. 10. Diferența dintre ADN și ARN la baza azotată: în loc de timină (T), ARN conține uracil (U), care este, de asemenea, complementar cu adenina.
TRANSCRIERE
Este procesul de sintetizare a ARN-ului pe un șablon de ADN. ADN-ul se desfășoară la unul dintre site-uri. Una dintre catene conține informații care trebuie copiate pe o moleculă de ARN - această catena se numește catena de codificare. A doua catenă de ADN, complementară celei de codificare, se numește șablon. În procesul de transcripție, o catena de ARN complementar este sintetizată pe catena șablon în direcția 3 '- 5' (de-a lungul catenei ADN). Astfel, se creează o copie ARN a firului de codare.
![]()
Orez. 11. Reprezentarea schematică a transcrierii
De exemplu, dacă ni se dă secvența șirului de codare
3'- ATGTCCTAGCTGCTCG - 5 ',
apoi, conform regulii complementarității, lanțul matricial va purta secvența
5'- TACAGGATCGACGAGC- 3 ',
iar ARN sintetizat din acesta este secvența
Transmisie
Luați în considerare mecanismul sinteza proteinei pe matricea ARN, precum și codul genetic și proprietățile sale. De asemenea, pentru claritate, folosind linkul de mai jos, vă recomandăm să vizionați un scurt videoclip despre procesele de transcriere și traducere care au loc într-o celulă vie:

Orez. 12. Procesul de sinteză a proteinelor: ADN codifică ARN, ARN codifică proteinele
COD GENETIC
Cod genetic- o metodă de codificare a secvenței de aminoacizi a proteinelor utilizând o secvență de nucleotide. Fiecare aminoacid este codificat de o secvență de trei nucleotide - un codon sau un triplet.
Cod genetic comun pentru majoritatea pro- și eucariote. Tabelul listează toți cei 64 de codoni și indică aminoacizii corespunzători. Ordinea de bază este de la capătul 5 "până la capătul 3" al ARNm.
Tabelul 1. Cod genetic standard
|
Primul nie |
A 2-a bază |
A treia nie |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe / F) |
U C U |
(Ser / S) |
U A U |
(Tyr / Y) |
U G U |
(Cys / C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu / L) |
U C A |
U A A |
Opriți codonul ** |
U G A |
Opriți codonul ** |
A |
||
|
U U G |
U C G |
U A G |
Opriți codonul ** |
U G G |
(Trp / W) |
G |
|||
|
C |
C U U |
C C U |
(Recuzită) |
C A U |
(His / H) |
C G U |
(Arg / R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln / Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile / I) |
A C U |
(Thr / T) |
A A U |
(Asn / N) |
A G U |
(Ser / S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys / K) |
A G A |
A |
||||
|
A U G |
(Met / M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val / V) |
G C U |
(Ala / A) |
G A U |
(Asp / D) |
G G U |
(Gly / G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu / E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Printre triplete, există 4 secvențe speciale care funcționează ca „semne de punctuație”:
- *Triplet AUG, care codifică și metionina, se numește începe codonul... Sinteza unei molecule de proteină începe de la acest codon. Astfel, în timpul sintezei proteinelor, primul aminoacid din secvență va fi întotdeauna metionina.
- ** Triplete UAA, UAGși UGA sunt numite opriți codoniiși nu codifică un singur aminoacid. La aceste secvențe, sinteza proteinelor se oprește.
Proprietățile codului genetic
1. Triplet... Fiecare aminoacid este codificat de o secvență de trei nucleotide - un triplet sau un codon.
2. Continuitate... Nu există nucleotide suplimentare între triplete, informațiile sunt citite continuu.
3. Non-suprapunere... O nucleotidă nu poate fi inclusă în două triplete în același timp.
4. Neambiguitate... Un codon poate codifica un singur aminoacid.

5. Degenerarea... Un aminoacid poate fi codificat de mai mulți codoni diferiți.

6. Versatilitate... Codul genetic este același pentru toate organismele vii.
Exemplu. Ni se dă secvența lanțului de codare:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Lanțul matricial va avea secvența:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Acum „sintetizăm” ARN-ul informațional din acest lanț:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Sinteza proteinelor merge în direcția 5 '→ 3', prin urmare, trebuie să răsturnăm secvența pentru a „citi” codul genetic:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
Acum să găsim codonul de pornire AUG:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
Să împărțim secvența în triplete:
sună așa: informațiile din ADN sunt transferate la ARN (transcripție), de la ARN - la proteine (traducere). ADN-ul poate fi, de asemenea, duplicat prin replicare, iar procesul de transcriere inversă este, de asemenea, posibil, atunci când ADN-ul este sintetizat din șablonul de ARN, dar acest proces este în principal tipic pentru viruși.

Orez. 13. Dogma centrală biologie moleculara
GENOMUL: GENE ȘI CROMOZOMI
(concepte generale)
Genom - totalitatea tuturor genelor unui organism; setul său complet de cromozomi.
Termenul „genom” a fost propus de G. Winkler în 1920 pentru a descrie un set de gene conținute într-un set haploid de cromozomi ai organismelor unei singure specii biologice. Înțelesul inițial al acestui termen a indicat faptul că conceptul de genom, spre deosebire de genotip, este o caracteristică genetică a speciei în ansamblu și nu a unui individ individual. Odată cu dezvoltarea geneticii moleculare, semnificația acestui termen s-a schimbat. Se știe că ADN-ul, care este purtătorul informațiilor genetice în majoritatea organismelor și, prin urmare, formează baza genomului, include nu numai gene în sensul modern al cuvântului. Majoritatea ADN-ului celulelor eucariote este reprezentat de secvențe de nucleotide necodificatoare („redundante”) care nu conțin informații despre proteine și acizi nucleici. Astfel, partea principală a genomului oricărui organism este întregul ADN al setului său haploid de cromozomi.
Genele sunt secțiuni ale moleculelor de ADN care codifică polipeptide și molecule de ARN
În secolul trecut, înțelegerea noastră asupra genelor s-a schimbat semnificativ. Anterior, genomul a fost numit o secțiune a unui cromozom care codifică sau determină o trăsătură sau fenotipic o proprietate (vizibilă), cum ar fi culoarea ochilor.
În 1940, George Beadle și Edward Tatem au propus o definiție moleculară a genei. Oamenii de știință au tratat sporii fungici Neurospora crassa Razele X și alți agenți care provoacă modificări ale secvenței ADN ( mutații), și au găsit tulpini mutante ale ciupercii care pierduseră anumite enzime specifice, ceea ce în unele cazuri a dus la întreruperea întregii căi metabolice. Beadle și Tatem au concluzionat că o genă este o bucată de material genetic care definește sau codifică o singură enzimă. Așa a apărut ipoteza „O genă - o enzimă”... Acest concept a fost ulterior extins pentru a defini „O genă - o polipeptidă”, deoarece multe gene codifică proteine care nu sunt enzime, iar polipeptida poate fi o subunitate a unui complex proteic complex.
În fig. 14 prezintă o diagramă a modului în care tripletele de nucleotide din ADN determină polipeptida, secvența de aminoacizi a unei proteine, mediată de ARNm. Una dintre firele de ADN joacă rolul unui șablon pentru sinteza ARNm, ale cărui triplete de nucleotide (codoni) sunt complementare tripletelor de ADN. La unele bacterii și multe eucariote, secvențele de codificare sunt întrerupte de regiuni necodificate (așa-numitul introni).
Definiție genetică biochimică modernă chiar mai specific. Genele sunt toate regiuni ale ADN-ului care codifică secvența primară a produselor finale, care includ polipeptide sau ARN-uri care au funcție structurală sau catalitică.
Alături de gene, ADN-ul conține și alte secvențe care îndeplinesc exclusiv o funcție de reglare. Secvențe de reglementare poate indica începutul sau sfârșitul genelor, poate afecta transcripția sau poate indica locul inițierii replicării sau recombinării. Unele gene pot fi exprimate în moduri diferite, aceeași bucată de ADN servind drept șablon pentru formarea diferitelor produse.
Putem calcula aproximativ dimensiunea minimă a genei codificarea proteinei medii. Fiecare aminoacid din lanțul polipeptidic este codificat ca o secvență de trei nucleotide; secvențele acestor triplete (codoni) corespund lanțului de aminoacizi din polipeptida codificată de gena dată. Lanțul polipeptidic de 350 de resturi de aminoacizi (lanț mediu) corespunde unei secvențe de 1050 bp. ( Perechi de baze). Cu toate acestea, multe gene eucariote și unele gene procariote sunt întrerupte de segmente de ADN care nu sunt purtători de informații despre o proteină și, prin urmare, se dovedește a fi mult mai lungă decât arată un simplu calcul.
Câte gene sunt pe un cromozom?
 Orez. 15. Vedere a cromozomilor în celulele procariote (stânga) și eucariote. Histonele sunt o clasă largă de proteine nucleare care îndeplinesc două funcții principale: sunt implicate în ambalarea firelor de ADN din nucleu și în reglarea epigenetică a proceselor nucleare, cum ar fi transcrierea, replicarea și repararea.
Orez. 15. Vedere a cromozomilor în celulele procariote (stânga) și eucariote. Histonele sunt o clasă largă de proteine nucleare care îndeplinesc două funcții principale: sunt implicate în ambalarea firelor de ADN din nucleu și în reglarea epigenetică a proceselor nucleare, cum ar fi transcrierea, replicarea și repararea.
ADN-ul procariotelor este mai simplu: celulele lor nu au nucleu, deci ADN-ul este localizat direct în citoplasmă sub forma unui nucleoid.
 După cum știți, celulele bacteriene au un cromozom sub forma unui fir ADN, ambalat într-o structură compactă - un nucleoid. Cromozomul unui procariot Escherichia coli, al cărui genom a fost complet decodificat, este o moleculă circulară de ADN (de fapt, nu este un cerc obișnuit, ci mai degrabă o buclă fără început și sfârșit), constând din 4 639 675 bp. Această secvență conține aproximativ 4300 de gene pentru proteine și 157 de gene pentru molecule de ARN stabile. V genomul uman aproximativ 3,1 miliarde de perechi de baze, corespunzând a aproape 29.000 de gene situate pe 24 de cromozomi diferiți.
După cum știți, celulele bacteriene au un cromozom sub forma unui fir ADN, ambalat într-o structură compactă - un nucleoid. Cromozomul unui procariot Escherichia coli, al cărui genom a fost complet decodificat, este o moleculă circulară de ADN (de fapt, nu este un cerc obișnuit, ci mai degrabă o buclă fără început și sfârșit), constând din 4 639 675 bp. Această secvență conține aproximativ 4300 de gene pentru proteine și 157 de gene pentru molecule de ARN stabile. V genomul uman aproximativ 3,1 miliarde de perechi de baze, corespunzând a aproape 29.000 de gene situate pe 24 de cromozomi diferiți.
Procariote (bacterii).
 Bacterie E coli are o moleculă de ADN circular dublu catenar. Se compune din 4.639.675 bp. și atinge o lungime de aproximativ 1,7 mm, care depășește lungimea celulei în sine E coli de aproximativ 850 de ori. În plus față de cromozomul circular mare din nucleoid, multe bacterii conțin una sau mai multe molecule mici de ADN circular care sunt localizate liber în citosol. Astfel de elemente extracromozomiale se numesc plasmide(fig. 16).
Bacterie E coli are o moleculă de ADN circular dublu catenar. Se compune din 4.639.675 bp. și atinge o lungime de aproximativ 1,7 mm, care depășește lungimea celulei în sine E coli de aproximativ 850 de ori. În plus față de cromozomul circular mare din nucleoid, multe bacterii conțin una sau mai multe molecule mici de ADN circular care sunt localizate liber în citosol. Astfel de elemente extracromozomiale se numesc plasmide(fig. 16).
Majoritatea plasmidelor constau doar din câteva mii de perechi de baze, unele conțin mai mult de 10.000 bp. Ei poartă informații genetice și se reproduc cu formarea de plasmide fiice, care intră în celulele fiice în timpul diviziunii celulei părinte. Plasmidele se găsesc nu numai în bacterii, ci și în drojdie și alte ciuperci. În multe cazuri, plasmidele nu oferă niciun avantaj celulelor gazdă, iar singura lor sarcină este reproducerea independentă. Cu toate acestea, unele plasmide poartă gene utile gazdei. De exemplu, genele conținute în plasmide pot conferi rezistență la agenți antibacterieni celulelor bacteriene. Plasmidele care poartă gena β-lactamază conferă rezistență la antibiotice β-lactamice precum penicilina și amoxicilina. Plasmidele pot fi transferate de la celule rezistente la antibiotice la alte celule ale aceluiași tip sau ale unui tip diferit de bacterii, determinând și aceste celule să devină rezistente. Utilizarea intensivă a antibioticelor este un factor selectiv puternic care promovează răspândirea plasmidelor care codifică rezistența la antibiotice (precum și transpozonii care codifică gene similare) printre bacteriile patogene și duce la apariția tulpinilor bacteriene cu rezistență la mai multe antibiotice. Medicii încep să înțeleagă pericolele utilizării pe scară largă a antibioticelor și le prescriu doar atunci când este nevoie urgentă. Din motive similare, utilizarea pe scară largă a antibioticelor pentru tratamentul animalelor de fermă este limitată.
Vezi si: Ravin N.V., Shestakov S.V. Genomul procariotelor // Vavilov Journal of Genetics and Selection, 2013. V. 17. No. 4/2. S. 972-984.
Eucariote.
Tabelul 2. ADN, gene și cromozomi ai unor organisme
|
ADN comun, p.n. |
Numărul cromozomului * |
Numărul aproximativ de gene |
|
|
Escherichia coli(bacterie) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(drojdie) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(nematod) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(plantă) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(musculiță de oțet) |
120 367 260 |
20 000 |
|
|
Oryza sativa(orez) |
480 000 000 |
57 000 |
|
|
Mus musculus(șoarece) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens(uman) |
3 070 128 600 |
29 000 |
Notă. Informațiile sunt actualizate constant; pentru informații mai actualizate, consultați site-urile dedicate proiectelor genomice individuale
* Pentru toate eucariotele, cu excepția drojdiei, se dă un set diploid de cromozomi. Diploid trusa cromozomi (din grecesc diploos- dublu și eidos- specie) - set dublu de cromozomi(2n), fiecare dintre acestea având unul omolog.
** Set Haploid. Tulpinile de drojdie sălbatică au de obicei opt (octaploide) sau mai multe seturi de astfel de cromozomi.
*** Pentru femelele cu doi cromozomi X. Bărbații au un cromozom X, dar nu Y, adică există doar 11 cromozomi.
O celulă de drojdie, unul dintre cele mai mici eucariote, are de 2,6 ori mai mult ADN decât o celulă E coli(Masa 2). Celulele mustei fructelor Drosophila, un obiect clasic al cercetării genetice, conține de 35 de ori mai mult ADN și celule umane - de aproximativ 700 de ori mai mult ADN decât celulele E coli. Multe plante și amfibieni conțin și mai mult ADN. Materialul genetic al celulelor eucariote este organizat sub formă de cromozomi. Set diploid de cromozomi (2 n) depinde de tipul de organism (Tabelul 2).
De exemplu, într-o celulă somatică umană există 46 de cromozomi ( orez. 17). Fiecare cromozom al unei celule eucariote, așa cum se arată în Fig. 17, A, conține o moleculă de ADN dublu catenar foarte mare. Douăzeci și patru de cromozomi umani (22 de cromozomi împerecheați și doi cromozomi sexuali X și Y) diferă ca lungime de peste 25 de ori. Fiecare cromozom eucariot conține un set specific de gene.

Orez. 17. Cromozomi eucarioti.A- o pereche de cromatide surori legate și condensate din cromozomul uman. În această formă, cromozomii eucariote rămân după replicare și în metafază în timpul mitozei. b- un set complet de cromozomi din leucocitul unuia dintre autorii cărții. Fiecare celulă somatică normală umană conține 46 de cromozomi.

Mărimea și funcția ADN-ului ca matrice pentru stocarea și transmiterea materialului ereditar explică prezența elementelor structurale speciale în organizarea acestei molecule. În organismele superioare, ADN-ul este distribuit între cromozomi.
Colecția de ADN (cromozomi) a unui organism se numește genom. Cromozomii se găsesc în nucleul celular și formează o structură numită cromatină. Cromatina este un complex de ADN și proteine de bază (histone) într-un raport 1: 1. Lungimea ADN-ului este de obicei măsurată prin numărul de perechi de nucleotide complementare (bp). De exemplu, al treilea cromozom este umansecolul este o moleculă de ADN cu o dimensiune de 160 milioane bp. ADN-ul izolat liniarizat cu o dimensiune de 3 * 10 6 bp. are o lungime de aproximativ 1 mm, prin urmare, molecula liniarizată a celui de-al treilea cromozom uman ar avea o lungime de 5 mm, iar ADN-ul tuturor celor 23 de cromozomi (~ 3 * 10 9 bp, MR = 1,8 * 10 12) al haploidului celula - ovocit sau spermă - în formă liniarizată ar fi de 1 m. Cu excepția celulelor germinale, toate celulele corpului uman (există aproximativ 1013 dintre ele) conțin un set dublu de cromozomi. În timpul diviziunii celulare, toate cele 46 de molecule de ADN sunt reproduse și reorganizate în 46 de cromozomi.
Dacă conectați moleculele ADN ale genomului uman (22 de cromozomi și cromozomi X și Y sau X și X), veți obține o secvență de aproximativ un metru lungime. Notă: Toate mamiferele și alte organisme cu sex masculin heterogametic, femelele au doi cromozomi X (XX), iar masculii au un cromozom X și un cromozom Y (XY).
Majoritatea celulelor umane, prin urmare, lungimea totală a ADN-ului acestor celule este de aproximativ 2m. Un adult are aproximativ 10 14 celule, deci lungimea totală a tuturor moleculelor de ADN este de 2 × 10 11 km. Pentru comparație, circumferința Pământului este de 4 ・ 10 4 km, iar distanța de la Pământ la Soare este de 1,5 ・ 10 8 km. Acesta este cât de surprinzător este ADN-ul ambalat compact în celulele noastre!
În celulele eucariote, există alte organite care conțin ADN - mitocondrii și cloroplaste. Au fost prezentate multe ipoteze cu privire la originea ADN-ului mitocondrial și cloroplast. Punctul de vedere general acceptat astăzi este că sunt rudimentele cromozomilor bacteriilor antice care au intrat în citoplasma celulelor gazdă și au devenit precursorii acestor organite. Codurile ADN mitocondrial pentru ARNt și ARNr mitocondrial, precum și mai multe proteine mitocondriale. Peste 95% din proteinele mitocondriale sunt codificate de ADN-ul nuclear.
STRUCTURA GENELOR
Luați în considerare structura genei în procariote și eucariote, asemănările și diferențele lor. În ciuda faptului că o genă este o bucată de ADN care codifică o singură proteină sau ARN, pe lângă partea care codifică direct, include și elemente de reglare și alte elemente structurale care au structuri diferite în procariote și eucariote.
Secvența de codificare- principala unitate structurală și funcțională a genei, este în ea că tripletele nucleotidelor care codificăsecvența de aminoacizi. Începe cu un codon de pornire și se termină cu un codon de oprire.
Înainte și după secvența de codare sunt secvențe 5 'și 3' netraduse... Ei îndeplinesc funcții de reglare și auxiliare, de exemplu, asigură aterizarea ribozomului pe ARN-m.
Secvențele netraduse și codificatoare constituie o unitate de transcripție - o secțiune ADN transcrisă, adică o secțiune ADN din care are loc sinteza ARN-ului.
Terminator- regiune ADN netranscrisă la sfârșitul genei, unde se oprește sinteza ARN.
La începutul genei este zona de reglementare inclusiv promotorși operator.
Promotor- secvența de care se leagă polimeraza în timpul inițierii transcrierii. Operator este o regiune la care proteinele speciale se pot lega - represori, care poate reduce activitatea sintezei ARN-ului din această genă - cu alte cuvinte, reduce-o expresie.
Structura genei în procariote
Structura generală a genelor din procariote și eucariote nu diferă - ambele conțin o regiune reglatoare cu un promotor și operator, o unitate de transcripție cu secvențe de codificare și netraduse și un terminator. Cu toate acestea, organizarea genelor în procariote și eucariote este diferită.

Orez. 18. Schema structurii genei în procariote (bacterii) -imaginea este mărită
La începutul și la sfârșitul operonului, există regiuni de reglare comune pentru mai multe gene structurale. O moleculă de ARNm este citită din regiunea transcrisă a operonului, care conține mai multe secvențe de codificare, fiecare dintre acestea având propriul său codon de pornire și oprire. Din fiecare dintre aceste site-uri cuo proteină este întreruptă. Prin urmare, mai multe molecule de proteine sunt sintetizate dintr-o moleculă de i-ARN.
Procariotele se caracterizează prin combinarea mai multor gene într-o singură unitate funcțională - operon... Lucrarea operonului poate fi reglată de alte gene care pot fi eliminate în mod vizibil din operon în sine - regulatori... Proteina tradusă din această genă se numește represor... Se leagă de operatorul operonului, reglând expresia tuturor genelor conținute în el deodată.
Fenomenul este caracteristic și procariotelor pereche transcriere și traducere.
![]()
Orez. 19 Fenomenul conjugării transcrierii și traducerii în procariote - imaginea este mărită
O astfel de conjugare nu are loc în eucariote datorită prezenței unui înveliș nuclear care separă citoplasma, unde are loc traducerea, de materialul genetic pe care are loc transcrierea. La procariote, în timpul sintezei ARN pe șablonul ADN, ribozomul se poate lega imediat de molecula de ARN sintetizată. Astfel, traducerea începe chiar înainte de finalizarea transcrierii. Mai mult, mai mulți ribozomi se pot lega simultan de o moleculă de ARN, sintetizând simultan mai multe molecule ale unei proteine.
Structura genei în eucariote
Genele și cromozomii eucariotelor sunt organizate foarte complex
Multe specii de bacterii au un singur cromozom și, în aproape toate cazurile, există o copie a fiecărei gene pe fiecare cromozom. Doar câteva gene, cum ar fi genele ARNr, sunt conținute în mai multe copii. Genele și secvențele reglatoare alcătuiesc practic întregul genom al procariotelor. Mai mult, aproape fiecare genă corespunde strict secvenței de aminoacizi (sau secvenței ARN) pe care o codifică (Fig. 14).
Organizarea structurală și funcțională a genelor eucariote este mult mai complexă. Studiul cromozomilor eucariote și secvențierea ulterioară a secvențelor complete ale genomului eucariot au adus multe surprize. Multe, dacă nu chiar majoritatea, genele eucariote au o caracteristică interesantă: secvențele lor de nucleotide conțin una sau mai multe regiuni ADN care nu codifică secvența de aminoacizi a produsului polipeptidic. Astfel de inserții netraduse rup corespondența directă între secvența nucleotidică a genei și secvența de aminoacizi a polipeptidei codificate. Aceste segmente netraduse de gene se numesc introni, sau încorporat secvențe iar segmentele de codificare sunt exoni... La procariote, doar câteva gene conțin introni.
Deci, în eucariote, practic nu există o combinație de gene în operoni, iar secvența de codare a genei eucariote este cel mai adesea împărțită în regiuni traduse. - exoni, și secțiuni netraduse - introni.
În majoritatea cazurilor, funcția intronilor nu a fost stabilită. În general, doar aproximativ 1,5% din ADN-ul uman „codifică”, adică transportă informații despre proteine sau ARN. Cu toate acestea, luând în considerare intronii mari, se dovedește că 30% din ADN-ul uman este format din gene. Deoarece genele alcătuiesc o proporție relativ mică din genomul uman, o parte semnificativă a ADN-ului rămâne neevaluată.

Orez. 16. Schema structurii genei în eucariote - imaginea este mărită
Din fiecare genă este sintetizat mai întâi imatur sau pre-ARN, care conține atât introni, cât și exoni.
După aceasta, are loc un proces de îmbinare, în urma căruia sunt excizate regiunile intron și se formează un ARNm matur, din care proteina poate fi sintetizată.

Orez. 20. Procesul de îmbinare alternativă - imaginea este mărită
O astfel de organizare a genelor face posibilă, de exemplu, realizarea când diferite forme ale unei proteine pot fi sintetizate dintr-o genă, datorită faptului că în procesul de îmbinare exonii pot fi cusute în diferite secvențe.

Orez. 21. Diferențe în structura genelor de procariote și eucariote - imaginea este mărită
MUTAȚII ȘI MUTAGENEZE
Mutaţie se numește o schimbare persistentă a genotipului, adică o modificare a secvenței nucleotidice.
Se numește procesul care duce la apariția mutațiilor mutageneză, și corpul, toate ale cărei celule poartă aceeași mutație - mutant.
Teoria mutației a fost formulată pentru prima dată de Hugo de Vries în 1903. Versiunea sa modernă include următoarele prevederi:
1. Mutațiile apar brusc, în salturi.
2. Mutațiile sunt transmise din generație în generație.
3. Mutațiile pot fi benefice, dăunătoare sau neutre, dominante sau recesive.
4. Probabilitatea de a detecta mutații depinde de numărul de indivizi examinați.
5. Mutații similare pot apărea în mod repetat.
6. Mutațiile nu sunt vizate.
Mutațiile pot apărea datorită diverșilor factori. Distingeți mutațiile care au apărut sub influență mutagen impacturi: fizice (de exemplu, ultraviolete sau radiații), chimice (de exemplu, colchicină sau specii reactive de oxigen) și biologice (de exemplu, viruși). De asemenea, mutațiile pot fi cauzate de erori de replicare.
În funcție de condițiile de apariție, mutațiile sunt împărțite în spontan- adică mutații care au apărut în condiții normale și induse- adică mutații care au apărut în condiții speciale.
Mutațiile pot apărea nu numai în ADN-ul nuclear, ci și, de exemplu, în ADN-ul mitocondriilor sau plastidelor. În consecință, putem distinge nuclearși citoplasmatic mutații.
Noile alele pot apărea adesea ca urmare a mutațiilor. Dacă o alelă mutantă suprimă acțiunea unei alele normale, mutația este numită dominant... Dacă o alelă normală suprimă un mutant, se numește o astfel de mutație recesiv... Majoritatea mutațiilor care duc la apariția de noi alele sunt recesive.
Prin efect, se disting mutațiile adaptativ ducând la o creștere a adaptării corpului la mediu, neutru care nu afectează supraviețuirea, dăunătoare care reduc adaptabilitatea organismelor la condițiile de mediu și mortal ducând la moartea organismului în stadiile incipiente ale dezvoltării.
Conform consecințelor, mutațiile se disting, ducând la pierderea funcției proteice, mutații care duc la ieșirea proteina are o nouă funcție, precum și mutații care modificați doza geneiși, în consecință, doza de proteină sintetizată din aceasta.
O mutație poate apărea în orice celulă din corp. Dacă apare o mutație în celula germinală, aceasta se numește germinal(germinal sau generativ). Astfel de mutații nu apar în organismul în care au apărut, dar duc la apariția mutanților la descendenți și sunt moștenite, prin urmare sunt importante pentru genetică și evoluție. Dacă apare o mutație în orice altă celulă, se numește somatic... O astfel de mutație se poate manifesta, într-un grad sau altul, în organismul în care a apărut, de exemplu, poate duce la formarea tumorilor canceroase. Cu toate acestea, această mutație nu este moștenită și nu afectează descendenții.
Mutațiile pot afecta regiuni ale genomului de diferite dimensiuni. Aloca genă, cromozomialeși genomică mutații.
Mutații genetice
Mutațiile care apar la o scară mai mică decât o genă sunt numite genetic, sau punct (punct)... Astfel de mutații duc la modificarea uneia sau mai multor nucleotide din secvență. Printre mutațiile genetice, existăînlocuiri conducând la înlocuirea unui nucleotid cu altul,ștergeri ducând la pierderea uneia dintre nucleotide,inserții ducând la adăugarea unui nucleotid suplimentar la secvență.

Orez. 23. Mutații genetice (punct)
Conform mecanismului de acțiune asupra proteinelor, mutațiile genetice sunt împărțite în:sinonim, care (ca urmare a degenerării codului genetic) nu duc la o modificare a compoziției aminoacizilor produsului proteic,mutații fără sens care duc la substituirea unui aminoacid cu altul și pot afecta structura proteinei sintetizate, deși adesea se dovedesc a fi nesemnificative,mutații aiurea conducând la înlocuirea codonului de codificare cu un codon de oprire,mutații care duc la tulburare de îmbinare:

Orez. 24. Scheme de mutații
De asemenea, conform mecanismului de acțiune asupra proteinei, mutațiile sunt izolate, ducând la schimbarea cadrului citiri de exemplu, inserții și ștergeri. Astfel de mutații, cum ar fi mutațiile fără sens, deși apar la un moment al unei gene, afectează adesea întreaga structură a unei proteine, ceea ce poate duce la o schimbare completă a structurii sale. când secțiunea cromozomială este rotită cu 180 de grade, Orez. 28. Translocare
când secțiunea cromozomială este rotită cu 180 de grade, Orez. 28. Translocare
Orez. 29. Cromozom înainte și după duplicare
Mutații genomice
In cele din urma, mutații genomice afectează întregul genom în ansamblu, adică se modifică numărul cromozomilor. Alocați poliploidia - o creștere a ploidiei celulare și a aneuploidiei, adică o modificare a numărului de cromozomi, de exemplu, trisomia (prezența unui omolog suplimentar într-unul dintre cromozomi) și monozomia (absența unui omolog într-un cromozom).
Videoclipuri ADN
REPLICAȚIE ADN, CODIFICARE ARN, SINTEZĂ PROTEINICĂ
(Dacă videoclipul nu este afișat, acesta este disponibil până la
Conform structurii sale chimice, ADN-ul ( Acidul dezoxiribonucleic) este un biopolimer, ale căror monomeri sunt nucleotide... Adică ADN-ul este polinucleotidă... Mai mult, o moleculă de ADN constă de obicei din două lanțuri, răsucite unele față de altele de-a lungul unei linii elicoidale (adesea numite „răsucite spiralat”) și conectate prin legături de hidrogen.
Lanțurile pot fi răsucite atât în partea stângă, cât și în partea dreaptă (cel mai adesea).
Unii viruși au o singură catenă de ADN.
Fiecare nucleotidă ADN constă din 1) o bază azotată, 2) dezoxiriboză, 3) un reziduu de acid fosforic.
Elice ADN duble dreapta
ADN-ul include următoarele: adenină, guanină, timinași citozină... Adenina și guanina aparțin purinam, și timină și citozină - până la pirimidină... Uneori uracilul face parte din ADN, care este de obicei caracteristic ARN-ului, unde înlocuiește timina.
Bazele azotate ale unui lanț al moleculei de ADN sunt conectate cu bazele azotate ale celuilalt strict conform principiului complementarității: adenina numai cu timină (formează două legături de hidrogen între ele) și guanina numai cu citozină (trei legături) .
Baza azotată din nucleotidă este conectată la primul atom de carbon de formă ciclică dezoxiriboză care este pentoză (un carbohidrat cu cinci atomi de carbon). Legătura este covalentă, glicozidică (C-N). Spre deosebire de riboză, deoxiribozei îi lipsește una dintre grupările hidroxil. Inelul de dezoxiriboză este format din patru atomi de carbon și un atom de oxigen. Al cincilea atom de carbon se află în afara inelului și este conectat printr-un atom de oxigen cu restul de acid fosforic. De asemenea, prin atomul de oxigen de la al treilea atom de carbon, este atașat reziduul de acid fosforic al nucleotidei adiacente.
Astfel, într-o singură catenă de ADN, nucleotidele adiacente sunt legate prin legături covalente între dezoxiriboză și acid fosforic (legătură fosfodiester). Se formează o coloană vertebrală fosfat-dezoxiriboză. În mod perpendicular pe acesta, spre cealaltă catenă de ADN, sunt direcționate bazele azotate, care sunt conectate la bazele celui de-al doilea lanț prin legături de hidrogen.
Structura ADN-ului este astfel încât coloanele vertebrale ale lanțurilor legate de hidrogen sunt direcționate în direcții diferite (se spune „multidirecțional”, „antiparalel”). Pe lateral, unde unul se termină cu acid fosforic legat de al cincilea atom de carbon al dezoxiribozei, celălalt se termină cu un al treilea atom de carbon "liber". Adică, scheletul unui lanț este răsturnat față de celălalt. Astfel, în structura lanțurilor ADN, se disting capetele 5 "și 3".
La replicarea (dublarea) ADN-ului, sinteza de noi catene merge întotdeauna de la capătul 5 la cel de-al treilea, deoarece noile nucleotide se pot atașa doar la capătul al treilea liber.
În cele din urmă (indirect prin ARN), la fiecare trei nucleotide consecutive din lanțul ADN codifică un aminoacid al proteinei.
Descoperirea structurii moleculei de ADN a avut loc în 1953 grație lucrărilor lui F. Crick și D. Watson (care a fost facilitată și de lucrările timpurii ale altor oameni de știință). Deși cum Substanta chimica ADN-ul era cunoscut încă din secolul al XIX-lea. În anii 40 ai secolului XX, a devenit clar că ADN-ul este purtătorul informațiilor genetice.
Helixul dublu este considerat o structură secundară a moleculei de ADN. În celulele eucariote, cantitatea covârșitoare de ADN este localizată în cromozomi, unde este legat de proteine și alte substanțe și, de asemenea, suferă un ambalaj mai dens.
Acidul dezoxiribonucleic sau ADN este un purtător de informații genetice. Majoritatea ADN-ului din celule este concentrat în nucleu. Este componenta principală a cromozomilor. În eucariote, ADN-ul se găsește și în mitocondrii și plastide. ADN-ul este format din mononucleotide legate covalent între ele, reprezentând un polimer lung, neramificat. Mononucleotidele care alcătuiesc ADN-ul sunt compuse din dezoxiriboză, una dintre cele 4 baze azotate (adenină, guanină, citozină și timină) și un reziduu de acid fosforic. Numărul acestor mononucleotide este foarte mare. De exemplu, în celulele procariote care conțin un singur cromozom, ADN-ul este o singură macromoleculă cu o greutate moleculară mai mare de 2 x 10 9.
Mononucleotidele unei catene de ADN sunt conectate în serie între ele datorită formării legături fosfodiester covalente între grupul OH al dezoxiribozei unui mononucleotid și restul de acid fosforic al altuia. Pe o parte a coloanei vertebrale formate a unei catene de ADN, se află baze azotate. Ele pot fi comparate cu patru mărgele diferite purtate pe același fir, deoarece par a fi strânse pe un lanț zahăr-fosfat.
Se pune întrebarea, cum poate acest lung lanț polinucleotidic să codifice un program pentru dezvoltarea unei celule sau chiar a unui organism întreg? Răspunsul la această întrebare poate fi obținut prin înțelegerea modului în care se formează structura spațială a ADN-ului. Structura acestei molecule a fost descifrată și descrisă de J. Watson și F. Crick în 1953.
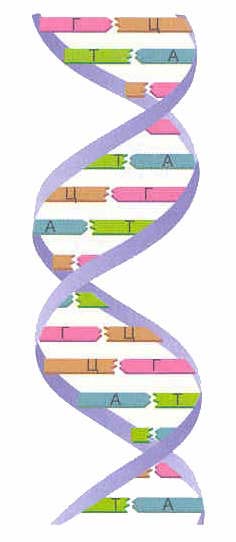
Moleculele ADN sunt două catene care se desfășoară paralel între ele și se formează spirala dreapta
... Lățimea acestei spirale este de aproximativ 2 nm, dar lungimea sa poate ajunge la sute de mii de nanometri. Watson și Crick au propus un model ADN, conform căruia toate bazele ADN sunt situate în interiorul helixului, cu o coloană vertebrală de zahăr-fosfat în exterior. Astfel, bazele unui lanț sunt cât mai apropiate de bazele celuilalt,
prin urmare, între ele se formează legături de hidrogen. Structura helixului ADN este astfel încât lanțurile polinucleotidice care îl alcătuiesc pot fi separate numai după ce a fost desfăcută.
Datorită apropierii maxime a două fire ADN, compoziția sa conține aceeași cantitate de baze azotate de un tip (adenină și guanină) și baze azotate de alt tip (timină și citozină), adică formula este valabilă: A + G = T + C... Acest lucru se datorează dimensiunii bazelor azotate, și anume, lungimea structurilor care se formează datorită apariției unei legături de hidrogen între perechile adenină-timină și guanină-citozină este de aproximativ 1,1 nm. Dimensiunile totale ale acestor perechi corespund dimensiunilor părții interioare a helixului ADN. Pentru a forma o spirală pereche C-T ar fi prea mic și pereche A-G dimpotrivă, este prea mare. Adică, baza azotată a primei catene de ADN definește baza care este situată în același loc pe cealaltă catenă de ADN. A fost numită corespondența strictă a nucleotidelor localizate în molecula ADN în lanțuri pereche paralele între ele complementaritate (adiționalitate). Reproducere exactă sau replicare informațiile genetice sunt posibile tocmai datorită acestei caracteristici a moleculei de ADN.
În ADN, informațiile biologice sunt înregistrate în așa fel încât să poată fi exact copiate și transmise celulelor descendente. Înainte de a se produce divizarea celulară în ea replicare (auto-dublare ) ADN. Deoarece fiecare catenă conține o secvență de nucleotide care este complementară secvenței lanțului partener, ele poartă de fapt aceleași informații genetice. Dacă separați catenele și le utilizați ca șablon (șablon) pentru a construi a doua catenă, veți obține două noi catene de ADN identice. Exact așa se produce duplicarea ADN-ului într-o celulă.


