DNK nimadan iborat? Nukleotid nima? DNK zanjiridagi nukleotidlarning tarkibi, tuzilishi, soni va ketma-ketligi DNKning qo'sh spiralini barqarorlashtiruvchi ionlar
Nuklein kislotalar mononukleotidlardan tashkil topgan yuqori molekulyar moddalar bo'lib, ular polimer zanjirida bir-biri bilan 3 ", 5" - fosfodiester bog'lari yordamida bog'langan va hujayralarga ma'lum tarzda qadoqlangan.
Nuklein kislotalar ikki turdagi biopolimerlar: ribonuklein kislotasi (RNK) va deoksiribonuklein kislotasi (DNK). Har bir biopolimer uglevod qoldig'i (riboza, dezoksiriboza) va azotli asoslardan biri (urasil, timin) bilan farq qiluvchi nukleotidlardan iborat. Ushbu farqlarga ko'ra, nuklein kislotalar o'z nomini oldi.
Dezoksiribonuklein kislota tuzilishi
Nuklein kislotalar birlamchi, ikkilamchi va uchinchi darajali tuzilishga ega.
Birlamchi DNK tuzilishi
DNKning birlamchi tuzilishi chiziqli polinukleotid zanjiri deb ataladi, unda mononukleotidlar 3 ", 5" -fosfodiester bog'lari bilan bog'langan. Hujayradagi nuklein kislotalar zanjirini yig'ish uchun boshlang'ich material nukleosid 5 "-trifosfat bo'lib, b va g fosforik kislota qoldiqlarini olib tashlash natijasida boshqa nukleozidning 3" uglerod atomini biriktirishga qodir. . Shunday qilib, bitta dezoksiribozaning 3 "uglerod atomi bir fosfor kislotasi qoldig'i orqali boshqa dezoksiribozaning 5" uglerod atomiga kovalent bog'lanadi va chiziqli polinukleotid nuklein kislotasi zanjirini hosil qiladi. Shuning uchun nomi: 3 ", 5" -fosfodiester bog'lari. Azotli asoslar bir zanjirning nukleotidlari birikmasida qatnashmaydi (1.-rasm).
Bir nukleotidning fosfor kislotasi molekulasining qolgan qismi va boshqasining uglevodlari o'rtasidagi bunday bog'lanish polinukleotid molekulasining pentoza-fosfat skeletining hosil bo'lishiga olib keladi, buning ustiga azotli asoslar birin-ketin yon tomondan biriktiriladi. Ularning nuklein kislota molekulalarining zanjirlarida joylashish ketma-ketligi qat'iy ravishda turli organizmlarning hujayralari uchun xosdir, ya'ni. o'ziga xos xususiyatga ega (Chargaff qoidasi).
Uzunligi zanjirga kiritilgan nukleotidlar soniga bog'liq bo'lgan chiziqli DNK zanjirining ikkita uchi bor: biri 3 "uchi" deb ataladi va erkin gidroksilni o'z ichiga oladi, ikkinchisi, 5" uchida fosfor kislotasi mavjud. qoldiq. Zanjir polarizatsiyalangan va 5 "-> 3" va 3 "-> 5" yo'nalishiga ega bo'lishi mumkin. Istisno - bu dumaloq DNK.
DNKning genetik "matn"i kod "so'zlari" - kodonlar deb ataladigan nukleotidlarning tripletlaridan iborat. RNKning barcha turlarining birlamchi tuzilishi haqidagi ma'lumotlarni o'z ichiga olgan DNK hududlari strukturaviy genlar deb ataladi.
Polinukleodit DNK zanjirlari ulkan o'lchamlarga etadi, shuning uchun ular hujayrada ma'lum bir tarzda o'ralgan.
DNK tarkibini o'rganib, Chargaff (1949) alohida DNK asoslari tarkibiga oid muhim qonunlarni o'rnatdi. Ular DNKning ikkilamchi tuzilishini ochishga yordam berdi. Bu naqshlar Chargaff qoidalari deb ataladi. Chargaff qoidalari
Ushbu qoidalar shuni ko'rsatadiki, DNKni qurishda purin va pirimidin asoslari o'rtasida umuman qat'iy muvofiqlik (juftlik) kuzatilmasligi kerak, lekin ayniqsa adenin bilan timin va guanin bilan sitozin. Ushbu qoidalarga asoslanib, jumladan, 1953 yilda Uotson va Krik DNKning ikkilamchi strukturasining qo'sh spiral deb nomlangan modelini taklif qildilar (rasm). |

DNKning ikkilamchi tuzilishi
DNKning ikkilamchi strukturasi qoʻsh spiral boʻlib, uning modeli 1953-yilda D.Uotson va F.Krik tomonidan taklif qilingan.
DNK modelini yaratish uchun zarur shartlar
Dastlabki tahlillar shuni ko'rsatdiki, har qanday kelib chiqishi DNK to'rtta nukleotidni teng molyar miqdorda o'z ichiga oladi. Biroq 1940-yillarda E.Chargaff va uning hamkorlari turli organizmlardan ajratilgan DNKni tahlil qilish natijasida ularda azotli asoslar turli miqdoriy nisbatlarda mavjudligini yaqqol ko'rsatdi. Chargaff shuni aniqladiki, bu nisbatlar bir xil turdagi organizmlarning barcha hujayralarining DNKlari uchun bir xil bo'lsa-da, har xil turdagi DNKlar ma'lum nukleotidlar tarkibida sezilarli darajada farq qilishi mumkin. Bu azotli asoslar nisbatidagi farqlar qandaydir biologik kod bilan bog'liq bo'lishi mumkinligini ko'rsatdi. Turli DNK namunalarida individual purin va pirimidin asoslarining nisbati har xil bo'lsa-da, tahlil natijalarini solishtirganda ma'lum bir qonuniyat paydo bo'ldi: barcha namunalarda purinlarning umumiy miqdori pirimidinlarning umumiy miqdoriga teng edi (A + G). = T + C), adenin miqdori timin miqdoriga (A = T) va guanin miqdori - sitozin miqdoriga (G = C) teng edi. Sutemizuvchilar hujayralaridan ajratilgan DNK odatda adenin va timinga boyroq, guanin va sitozinga nisbatan kambag'al bo'lsa, bakteriyalarda DNK guanin va sitozinga boyroq, adenin va timinga nisbatan kambag'al edi. Ushbu ma'lumotlar faktik materialning muhim qismini tashkil etdi, ular asosida keyinchalik Uotson-Krik DNK tuzilishi modeli qurilgan.
DNKning mumkin bo'lgan tuzilishining yana bir muhim bilvosita belgisi L. Paulingning oqsil molekulalarining tuzilishi haqidagi ma'lumotlari edi. Pauling oqsil molekulasidagi aminokislotalar zanjirining turli xil barqaror konfiguratsiyalari mumkinligini ko'rsatdi. Peptid zanjirining umumiy konfiguratsiyalaridan biri a-spiral muntazam spiralsimon strukturadir. Bunday tuzilish bilan zanjirning qo'shni burilishlarida joylashgan aminokislotalar o'rtasida vodorod aloqalarining shakllanishi mumkin. Pauling 1950 yilda polipeptid zanjirining a-spiral konfiguratsiyasini tasvirlab berdi va DNK molekulalari ham, ehtimol, vodorod bog'lari bilan mustahkamlangan spiral tuzilishga ega bo'lishini taklif qildi.
Biroq, DNK molekulasining tuzilishi haqidagi eng qimmatli ma'lumotlar rentgen strukturaviy tahlil natijalari bilan ta'minlandi. DNK kristalidan o'tuvchi rentgen nurlari difraksiyaga uchraydi, ya'ni ular ma'lum bir yo'nalishda buriladi. Nurlarning burilish darajasi va tabiati molekulalarning tuzilishiga bog'liq. Rentgen nurlari diffraktsiyasi (3-rasm) tajribali ko'zga tekshirilayotgan moddaning molekulalarining tuzilishiga oid bir qator bilvosita ko'rsatmalar beradi. DNKning rentgen nurlari diffraktsiya naqshlarini tahlil qilish, azotli asoslar (tekis shaklga ega) plastinkalar to'plami kabi yig'ilgan degan xulosaga keldi. Rentgen nurlari diffraktsiyasi kristalli DNK tuzilishining uchta asosiy davrini aniqladi: 0,34, 2 va 3,4 nm.

Watson-Crick DNK modeli
Chargaffning analitik ma'lumotlariga asoslanib, Uilkins tomonidan olingan rentgen nurlari difraksiyasi naqshlari va molekuladagi atomlar orasidagi aniq masofalar, berilgan atomning bog'lanishlari orasidagi burchaklar va atomlarning o'lchamlari haqida ma'lumot bergan kimyogarlarning tadqiqotlari. Uotson va Krik DNK molekulasining alohida tarkibiy qismlarining fizik modellarini ma'lum miqyosda qurishni boshladilar va ularni bir-biriga "moslashtirdilar", natijada olingan tizim turli xil eksperimental ma'lumotlarga mos keladi. [ko'rsatish] .
DNK zanjiridagi qo'shni nukleotidlar bir nukleotidning 5'-uglerodli dezoksiriboza atomini keyingi nukleotidning 3'-uglerodli dezoksiriboza atomi bilan bog'laydigan fosfodiester ko'priklar orqali bog'langanligi avvalroq ma'lum edi. Uotson va Krik 0,34 nm davri DNK zanjiridagi ketma-ket nukleotidlar orasidagi masofaga mos kelishiga shubha qilishmagan. Bundan tashqari, 2 nm davri zanjir qalinligiga to'g'ri keladi deb taxmin qilish mumkin. Qaysi haqiqiy tuzilma 3,4 nm davriga to'g'ri kelishini tushuntirish uchun, Uotson va Krik, xuddi Pauling singari, zanjirning spiral shaklida o'ralganligini (yoki aniqrog'i, spiral chiziq hosil qiladi, chunki a spiral bu so'z qat'iy ma'noda burilishlar kosmosda silindrsimon sirtni emas, balki konusni hosil qilganda olinadi). Keyin 3,4 nm davri ushbu spiralning ketma-ket burilishlari orasidagi masofaga to'g'ri keladi. Bunday spiral juda zich yoki biroz cho'zilgan bo'lishi mumkin, ya'ni uning burilishlari yumshoq yoki tik bo'lishi mumkin. 3,4 nm davri ketma-ket nukleotidlar orasidagi masofadan (0,34 nm) roppa-rosa 10 barobar ko'p bo'lganligi sababli, spiralning har bir to'liq aylanishida 10 ta nukleotid borligi aniq. Ushbu ma'lumotlardan Uotson va Krik burilishlar orasidagi masofa 3,4 nm ga teng bo'lgan diametri 2 nm bo'lgan spiralga o'ralgan polinukleotid zanjirining zichligini hisoblashga muvaffaq bo'ldi. Ma'lum bo'lishicha, bunday zanjirning zichligi allaqachon ma'lum bo'lgan DNKning haqiqiy zichligining yarmiga teng bo'ladi. Men DNK molekulasi ikkita zanjirdan iborat deb taxmin qilishim kerak edi - bu nukleotidlarning qo'sh spiralidir.
Keyingi vazifa, albatta, qo'sh spiralni tashkil etuvchi ikkita kontur o'rtasidagi fazoviy munosabatni aniqlashtirish edi. Uotson va Krik bir qator zanjir sxemalarini o'zlarining jismoniy modelida sinab ko'rganlaridan so'ng, barcha mavjud ma'lumotlar ikkita polinukleotid spirallari qarama-qarshi yo'nalishda harakatlanadigan variantga eng mos kelishini aniqladilar; bu holda shakar va fosfat qoldiqlaridan tashkil topgan zanjirlar qo'sh spiral sirtini hosil qiladi va purinlar va pirimidinlar ichida joylashgan. Ikki zanjirga mansub, bir-biriga qarama-qarshi joylashgan asoslar vodorod bog'lari orqali juft bo'lib bog'langan; Aynan shu vodorod aloqalari zanjirlarni bir-biriga bog'lab turadi va shu bilan molekulaning umumiy konfiguratsiyasini mustahkamlaydi.
DNKning qo'sh spiralini spiral shaklidagi arqon narvon sifatida tasavvur qilish mumkin, shunda uning zinapoyalari gorizontal holatda qoladi. Keyin ikkita uzunlamasına arqon shakar va fosfat qoldiqlari zanjirlariga to'g'ri keladi va shpallar vodorod aloqalari bilan bog'langan azotli asoslarning juftlariga mos keladi.
Mumkin bo'lgan modellarni keyingi o'rganish natijasida Watson va Crick har bir "bar" bitta purin va bitta pirimidindan iborat bo'lishi kerak degan xulosaga kelishdi; 2 nm davri bilan (bu qo'sh spiralning diametriga to'g'ri keladi), ikkita purin uchun etarli joy bo'lmaydi va ikkita pirimidin to'g'ri vodorod aloqalarini hosil qilish uchun bir-biriga etarlicha yaqin bo'lmaydi. Batafsil modelni chuqur o'rganish shuni ko'rsatdiki, adenin va sitozin o'lchamlari bo'yicha mos kombinatsiyani tashkil etsa ham, ular o'rtasida vodorod aloqalarini hosil qiladigan tarzda joylasha olmaydi. Shunga o'xshash hisobotlar guanin-timin birikmasini istisno qilishga majbur qildi, adenin-timin va guanin-sitozin birikmalari esa juda maqbul edi. Vodorod aloqalarining tabiati shundayki, adenin timin bilan, guanin esa sitozin bilan juft hosil qiladi. Maxsus asoslar juftligining bu kontseptsiyasi "Chargaff qoidasini" tushuntirishga imkon berdi, unga ko'ra har qanday DNK molekulasida adenin miqdori doimo timin tarkibiga, guanin miqdori esa sitozin miqdoriga teng bo'ladi. Adenin va timin o'rtasida ikkita vodorod bog'i, guanin va sitozin o'rtasida uchta vodorod bog'i hosil bo'ladi. Bir zanjirdagi har bir adeninga nisbatan vodorod bog'larini hosil qilishda ana shunday o'ziga xoslik tufayli, ikkinchisida timin topiladi; xuddi shunday, har bir guaninga qarshi faqat sitozin topilishi mumkin. Shunday qilib, zanjirlar bir-birini to'ldiradi, ya'ni bir zanjirdagi nukleotidlar ketma-ketligi ularning ikkinchisida ketma-ketligini o'ziga xos tarzda belgilaydi. Ikki zanjir qarama-qarshi yo'nalishda harakat qiladi va ularning terminal fosfat guruhlari qo'sh spiralning qarama-qarshi uchlarida joylashgan.
O'z tadqiqotlari natijasida 1953 yilda Uotson va Krik DNK molekulasi strukturasi modelini taklif qildilar (3-rasm), bu hozirgi kungacha dolzarbligicha qolmoqda. Modelga ko'ra, DNK molekulasi ikkita qo'shimcha polinukleotid zanjiridan iborat. Har bir DNK zanjiri bir necha o'n minglab nukleotidlardan iborat polinukleotiddir. Unda qo'shni nukleotidlar fosfor kislotasi qoldig'i va dezoksiribozaning kuchli kovalent bog' bilan bog'lanishi tufayli muntazam pentoza-fosfat magistralini hosil qiladi. Bunda bir polinukleotid zanjirining azotli asoslari ikkinchisining azotli asoslariga nisbatan qat’iy belgilangan tartibda joylashadi. Polinukleotid zanjirida azotli asoslarning almashinishi tartibsizdir.
DNK zanjiridagi azotli asoslarning joylashuvi komplementar (yunoncha "to'ldiruvchi" - qo'shilishdan), ya'ni. adeninga (A) qarshi doimo timin (T) va guaninga (G) qarshi faqat sitozin (C) mavjud. Bu A va T, shuningdek, G va C bir-biriga qat'iy mos kelishi bilan bog'liq, ya'ni. bir-birini to‘ldiradi. Bu yozishmalar purin va pirimidin juftligida vodorod aloqalarini hosil qilish imkonini beruvchi asoslarning kimyoviy tuzilishi bilan berilgan. A va T o'rtasida ikkita, G va C o'rtasida uchta bog'lanish mavjud. Bu bog'lanishlar DNK molekulasining kosmosda qisman barqarorlashuvini ta'minlaydi. Bunday holda, qo'sh spiralning barqarorligi A = T bog'lanishlari bilan solishtirganda barqarorroq bo'lgan G≡C bog'lanishlari soniga to'g'ridan-to'g'ri proportsionaldir.
Bir DNK zanjiridagi nukleotidlarning joylashishining ma'lum ketma-ketligi komplementarlik printsipiga ko'ra boshqa zanjirning nukleotidlarini o'rnatishga imkon beradi.
Bundan tashqari, suvli eritmada aromatik tuzilishga ega bo'lgan azotli asoslar bir-birining ustiga joylashib, go'yo tangalar to'plamini tashkil etishi aniqlandi. Bu staklarni shakllantirish jarayoni organik molekulalar staking deb ataladi. Ko'rib chiqilayotgan Uotson-Krik modelining DNK molekulasining polinukleotid zanjirlari o'xshash fizik-kimyoviy holatga ega, ularning azotli asoslari tangalar to'plami shaklida joylashgan bo'lib, ularning tekisliklari orasida van der Vaals o'zaro ta'siri (stacking o'zaro ta'sirlari) paydo bo'ladi.
Van-der-Vaals kuchlari (vertikal) ta’sirida polinukleotid zanjiridagi asoslar tekisliklari o‘rtasidagi komplementar asoslar (gorizontal) va stacking o‘zaro ta’siri o‘rtasidagi vodorod aloqalari DNK molekulasini kosmosda qo‘shimcha barqarorlashtirishni ta’minlaydi.
Ikkala zanjirning qand-fosfat umurtqalari tashqi tomonga, asoslari esa bir-biriga qaragan. DNKdagi zanjirlarning yo'nalishi antiparallel (ulardan biri 5 "-> 3" yo'nalishiga ega, ikkinchisi - 3 "-> 5", ya'ni 3 "bir zanjirning uchi ikkinchisining 5" uchiga qarama-qarshidir. .). Zanjirlar umumiy o'qga ega bo'lgan o'ng qo'lli spirallarni hosil qiladi. Spiralning bir burilishi 10 ta nukleotid, bobinning o'lchami 3,4 nm, har bir nukleotidning balandligi 0,34 nm va spiralning diametri 2,0 nm. Bir ipning ikkinchi ip atrofida aylanishi natijasida DNK qo'sh spiralining katta yivi (diametri taxminan 20 Å) va kichik yiv (taxminan 12 Å) hosil bo'ladi. Uotson-Krik qo'sh spiralining bu shakli keyinchalik B-shakli deb nomlandi. Hujayralarda DNK odatda eng barqaror bo'lgan B shaklida mavjud.
DNK funktsiyalari
Taklif etilayotgan model dezoksiribonuklein kislotasining ko'plab biologik xususiyatlarini, shu jumladan genetik ma'lumotni saqlash va 4 nukleotidning turli xil ketma-ket birikmalari bilan ta'minlangan genlarning xilma-xilligini va genetik kodning mavjudligi, o'z-o'zini ko'paytirish qobiliyatini tushuntirdi. replikatsiya jarayoni va genetik ma'lumotni amalga oshirish orqali ta'minlangan genetik ma'lumotni oqsillar shaklida, shuningdek ferment oqsillari yordamida hosil bo'lgan har qanday boshqa birikmalar shaklida o'tkazish.
DNKning asosiy funktsiyalari.
- DNK genetik ma'lumotlarning tashuvchisi bo'lib, bu genetik kodning mavjudligi fakti bilan ta'minlanadi.
- Hujayralar va organizmlarning avlodlarida genetik ma'lumotlarni ko'paytirish va uzatish. Bu funksiya replikatsiya jarayoni bilan ta'minlanadi.
- Proteinlar, shuningdek, oqsil-fermentlar yordamida hosil bo'lgan boshqa har qanday birikmalar shaklida genetik ma'lumotni amalga oshirish. Bu funksiya transkripsiya va tarjima jarayonlari bilan ta'minlanadi.

Ikki zanjirli DNKni tashkil qilish shakllari
DNK bir necha turdagi qo'sh spiral hosil qilishi mumkin (4-rasm). Hozirgi vaqtda oltita shakl allaqachon ma'lum (A dan E va Z-shakl).
Rozalind Franklin aniqlaganidek, DNKning strukturaviy shakllari nuklein kislota molekulasidagi suv bilan toʻyinganligiga bogʻliq. Rentgen strukturaviy tahlil yordamida DNK tolalarini tadqiq qilishda rentgen nurlarining diffraktsiya sxemasi qanday nisbiy namlikda, ushbu tolaning suv bilan to'yinganligi darajasida tajriba o'tkazilishiga tubdan bog'liq ekanligi ko'rsatildi. Agar tola suv bilan etarlicha to'yingan bo'lsa, u holda bitta rentgen nuri olingan. Quritganda, namlik yuqori bo'lgan tolaning rentgenogrammasidan juda farq qiladigan butunlay boshqa rentgen tasviri paydo bo'ldi.
Yuqori namlikdagi DNK molekulasi B-formasi deb ataladi... Fiziologik sharoitda (tuzning past konsentratsiyasi, yuqori darajada gidratlanish) DNKning dominant strukturaviy turi B-shakli (ikki zanjirli DNKning asosiy shakli Uotson-Krik modelidir). Bunday molekulaning spiral qadami 3,4 nm. "Tangalar" - azotli asoslarning o'ralgan stakalari ko'rinishida navbatma-navbat 10 ta qo'shimcha juftlik mavjud. Staklar stackning ikkita qarama-qarshi "tangalari" orasidagi vodorod aloqalari bilan ushlab turiladi va o'ng qo'l spiraliga o'ralgan fosfodiester skeletining ikkita lentasiga "o'ralgan". Azotli asoslar tekisliklari spiral o'qiga perpendikulyar. Qo'shni to'ldiruvchi juftliklar bir-biriga nisbatan 36 ° ga aylantiriladi. Spiralning diametri 20 Å, purin nukleotidi 12 Å va pirimidin nukleotidi 8 Å.
Pastroq namlikdagi DNK molekulasi A-formasi deb ataladi... A-shakli kamroq yuqori hidratsiya sharoitida va Na + yoki K + ionlarining yuqori miqdori bilan hosil bo'ladi. Bu kengroq o'ng qo'l konformatsiyasi har bir burilishda 11 ta asosiy juftlikka ega. Azotli asoslar tekisliklari spiral o'qga nisbatan kuchliroq moyillikka ega, ular odatdagidan spiral o'qga 20 ° ga buriladi. Demak, diametri 5 Å bo'lgan ichki bo'shliqning mavjudligi. Qo'shni nukleotidlar orasidagi masofa 0,23 nm, bobin uzunligi 2,5 nm, spiral diametri 2,3 nm.
Dastlab, DNKning A-shakli unchalik muhim emas deb hisoblangan. Biroq, keyinchalik DNK ning A-shakli, shuningdek, B-shaklining juda katta biologik ahamiyati borligi ma'lum bo'ldi. Shablon-primer kompleksidagi RNK-DNK spiral A-shakliga, shuningdek, RNK-RNK spiraliga va RNK soch turmagi tuzilmalariga ega (ribozaning 2'-gidroksil guruhi RNK molekulalarining B-shaklini hosil qilishiga imkon bermaydi) . DNK ning A shakli munozaralarda topilgan. DNKning A-shakli B-shakliga qaraganda UV nurlariga 10 marta chidamli ekanligi aniqlandi.
A-shakl va B-shakl deyiladi kanonik shakllar DNK.
C-E shakllari shuningdek, o'ng qo'llar, ularning shakllanishi faqat maxsus tajribalarda kuzatilishi mumkin va, aftidan, ular in vivo mavjud emas. C shaklidagi DNK B-DNKga o'xshash tuzilishga ega. Har bir burilishda asosiy juftliklar soni 9,33, spiral uzunligi 3,1 nm. Asosiy juftliklar o'qga perpendikulyar holatga nisbatan 8 graduslik burchak ostida egiladi. Yivlar hajmi jihatidan B-DNK yivlariga o'xshaydi. Bunday holda, asosiy truba biroz sayozroq, kichik truba esa chuqurroqdir. Tabiiy va sintetik DNK polinukleotidlari C shakliga o'tishi mumkin.
| Jadval 1. DNK tuzilmalarining ayrim turlarining xarakteristikalari | |||
| Spiral turi | A | B | Z |
| Spiral qadam | 0,32 nm | 3,38 nm | 4,46 nm |
| Spiral burilish | To'g'ri | To'g'ri | Chapga |
| Har bir burilish uchun asosiy juftliklar | 11 | 10 | 12 |
| Asosiy tekisliklar orasidagi masofa | 0,256 nm | 0,338 nm | 0,371 nm |
| Glikozid bog'lanish konformatsiyasi | qarshi | qarshi | anti-C sin-g |
| Furanoz siklining konformatsiyasi | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-c |
| Yiv kengligi kichik / katta | 1,11 / 0,22 nm | 0,57 / 1,17 nm | 0,2 / 0,88 nm |
| Chuqurlik chuqurligi, kichik / katta | 0,26 / 1,30 nm | 0,82 / 0,85 nm | 1,38 / 0,37 nm |
| Spiral diametri | 2,3 nm | 2,0 nm | 1,8 nm |
DNKning strukturaviy elementlari
(kanonik bo'lmagan DNK tuzilmalari)
DNKning strukturaviy elementlari ba'zi maxsus ketma-ketliklar bilan chegaralangan g'ayrioddiy tuzilmalarni o'z ichiga oladi:
|

Z shaklidagi DNK 1979 yilda geksanukleotid d (CG) 3 - ni o'rganish paytida kashf etilgan. Uni MIT professori Aleksandr Rich va uning hamkasblari kashf etgan. Z-shakli DNKning eng muhim tuzilmaviy elementlaridan biriga aylandi, chunki uning shakllanishi purinlar pirimidinlar bilan almashinadigan DNK mintaqalarida (masalan, 5'-HCGCH-3 ') yoki 5' takrorida kuzatilgan. -CHCH-3' tarkibida metillangan sitozin. Z-DNKning shakllanishi va barqarorlashuvining muhim sharti sin-konformatsiyada purin nukleotidlarining mavjudligi, antikonformatsiyada pirimidin asoslari bilan almashinishi edi.
Tabiiy DNK molekulalari odatda to'g'ri B-shaklda mavjud bo'lsa, ularda (CH) n tipidagi ketma-ketliklar bo'lmasa. Ammo, agar bunday ketma-ketliklar DNK tarkibiga kiritilgan bo'lsa, u holda eritmaning ion kuchi yoki fosfodiester magistralidagi manfiy zaryadni neytrallovchi kationlar o'zgarganda, Z-shakliga aylanishi mumkin, zanjirdagi boshqa DNK hududlari esa DNKda qoladi. klassik B shakli. Bunday o'tish imkoniyati shuni ko'rsatadiki, DNK qo'sh spiralidagi ikkita zanjir dinamik holatda bo'lib, o'ng shakldan chapga va aksincha, bir-biriga nisbatan bo'shashishi mumkin. DNK strukturasining konformatsion o'zgarishlariga imkon beruvchi bunday labillikning biologik oqibatlari hali to'liq tushunilmagan. Z-DNK hududlari ba'zi genlarning ifodalanishini tartibga solishda rol o'ynaydi va genetik rekombinatsiyada ishtirok etadi, deb ishoniladi.
DNK ning Z-shakli chap qo'l qo'sh spiral bo'lib, unda fosfodiester magistral molekula o'qi bo'ylab zigzag shaklida joylashgan. Shuning uchun molekulaning nomi (zigzag) -DHK. Z-DNK tabiatda ma'lum bo'lgan eng kam burilishli (har bir burilishda 12 ta asosiy juft) va eng nozik hisoblanadi. Qo‘shni nukleotidlar orasidagi masofa 0,38 nm, bobin uzunligi 4,56 nm, Z-DNK diametri esa 1,8 nm. Bundan tashqari, tashqi ko'rinish bu DNK molekulasi bitta yiv mavjudligi bilan ajralib turadi.
DNKning Z-shakli prokaryotik va eukaryotik hujayralarda topilgan. Hozirgi vaqtda Z-shaklini DNKning B-shaklidan ajrata oladigan antikorlar olindi. Bu antikorlar Drosophila (Dr. melanogaster) tuprik bezlari hujayralarining gigant xromosomalarining o'ziga xos hududlariga bog'lanadi. Ushbu xromosomalarning g'ayrioddiy tuzilishi tufayli bog'lanish reaktsiyasini kuzatish oson, ularda zichroq hududlar (disklar) kamroq zichroq mintaqalar (disklararo) bilan farq qiladi. Z-DNK hududlari interbandlarda joylashgan. Bundan kelib chiqadiki, Z-shakli aslida tabiiy sharoitda mavjud, ammo Z-shaklning alohida bo'limlarining o'lchamlari hali ham noma'lum.

(shifters) DNKdagi eng mashhur va tez-tez topiladigan asosiy ketma-ketliklardir. Palindrom - chapdan o'ngga va aksincha, xuddi shu tarzda o'qiladigan so'z yoki ibora. Bunday so‘z yoki iboralarga misol bo‘la oladi: SHALASH, QOZOQ, POTOP VA AZOR panjasiga atirgul tushdi. DNK mintaqalariga nisbatan qo'llanilganda, bu atama (palindrom) zanjir bo'ylab o'ngdan chapga va chapdan o'ngga ("kulba" so'zidagi harflar kabi) nukleotidlarning bir xil almashinuvini anglatadi.
Palindrom ikkita DNK zanjiriga nisbatan ikkinchi darajali simmetriyaga ega bo'lgan asosiy ketma-ketliklarning teskari takrorlanishi bilan tavsiflanadi. Bunday ketma-ketliklar, to'liq tushunarli sabablarga ko'ra, o'z-o'zini to'ldiradi va soch turmagi yoki xoch shaklidagi tuzilmalarni shakllantirishga moyildir (rasm). Soch iplari tartibga soluvchi oqsillarga xromosoma DNKsining genetik matni yozilgan joyni aniqlashga yordam beradi.
Xuddi shu DNK zanjirida teskari takrorlanish mavjud bo'lgan hollarda, bu ketma-ketlik oyna takrori deb ataladi. Ko'zgu takrorlashlari o'z-o'zini to'ldirish xususiyatlariga ega emas va shuning uchun soch turmagi yoki xoch shaklidagi tuzilmalarni shakllantirishga qodir emas. Ushbu turdagi ketma-ketliklar deyarli barcha yirik DNK molekulalarida uchraydi va bir necha tayanch juftlikdan bir necha ming tayanch juftlikgacha bo'lishi mumkin.
Eukaryotik hujayralardagi xochsimon tuzilmalar koʻrinishidagi palindromlarning mavjudligi isbotlanmagan, garchi bir qancha xochsimon tuzilmalar in vivo jonli holda E. coli hujayralarida topilgan. RNK yoki bir zanjirli DNKda o'z-o'zini to'ldiruvchi ketma-ketliklarning mavjudligi eritmalardagi nuklein zanjirining ma'lum fazoviy tuzilishga burmalanishining asosiy sababi bo'lib, ko'plab "soch tolalari" hosil bo'lishi bilan tavsiflanadi.

H shaklidagi DNK uch DNK zanjiridan hosil bo'lgan spiraldir - DNK uch spiral. Bu Uotson-Krik qo'sh spiral kompleksi bo'lib, uchinchi bir zanjirli DNK zanjiri bo'lib, uning katta yiviga to'g'ri keladi va Hoogsteen juftligi deb ataladi.
Bunday tripleksning hosil bo'lishi DNK qo'sh spiralining shunday buklanishi natijasida sodir bo'ladi, uning kesimining yarmi qo'sh spiral shaklida qoladi, ikkinchi yarmi esa uzilib qoladi. Bunday holda, ajratilgan spirallardan biri qo'sh spiralning birinchi yarmi bilan yangi tuzilmani hosil qiladi - uch spiral, ikkinchisi esa tuzilmagan bo'lib, bir ipli qism shaklida chiqadi. Ushbu tizimli o'tishning o'ziga xos xususiyati protonlari yangi tuzilmani barqarorlashtiradigan muhitning pH qiymatiga keskin bog'liqlikdir. Ushbu xususiyat tufayli yangi tuzilma DNKning H-shakli deb nomlandi, uning shakllanishi ko'zgu takrori bo'lgan gomopurin-gomopirimidin hududlarini o'z ichiga olgan superoillangan plazmidlarda topilgan.
Keyingi tadqiqotlarda ba'zi bir gomopurin-homopirimidin ikki zanjirli polinukleotidlarning uchta zanjirli tuzilish hosil bo'lishi bilan tarkibiy o'tish imkoniyati mavjud:
- bitta gomopurin va ikkita gomopirimidin zanjiri ( Py-Pu-Py tripleksi) [Hoogsteen shovqini].
Py-Pu-Py tripleksining tarkibiy bloklari CGC + va TAT ga izomorf kanonik triadalardir. Tripleksni barqarorlashtirish CGC + triadaning protonatsiyasini talab qiladi, shuning uchun bu triplekslar eritmaning pH darajasiga bog'liq.
- bitta gomopirimidin va ikkita gomopurin zanjiri ( Py-Pu-Pu tripleksi) [teskari Hoogsteen shovqini].
Py-Pu-Pu tripleksining tarkibiy bloklari CGG va TAA triadalari uchun kanonik izomorfikdir. Py-Pu-Pu triplekslarining muhim xususiyati ularning barqarorligining ikki marta zaryadlangan ionlar mavjudligiga bog'liqligi va turli ketma-ketlikdagi triplekslarni barqarorlashtirish uchun turli ionlar talab qilinadi. Py-Pu-Pu triplekslarining hosil bo'lishi ularni tashkil etuvchi nukleotidlarning protonatsiyasini talab qilmaganligi sababli, bunday triplekslar neytral pH da mavjud bo'lishi mumkin.
Eslatma: Hoogsteenning to'g'ridan-to'g'ri va teskari o'zaro ta'siri 1-metiltiminning simmetriyasi bilan izohlanadi: 180 ° aylanish O4 atomining O2 atomi bilan almashtirilishiga olib keladi, shu bilan birga vodorod aloqalari tizimi saqlanib qoladi.
Ikki turdagi uch spiral mavjud:
- uchinchi ipning qutbliligi Watson-Crick dupleksining gomopurin zanjirining qutbliligiga to'g'ri keladigan parallel uchburchaklar.
- antiparallel uch spirallar, ularda uchinchi va gomopurin zanjirlarining qutblari qarama-qarshidir.

G-to'rtlik- 4 zanjirli DNK. Bunday tuzilma, agar to'rtta guanin bo'lsa, hosil bo'ladi, ular G-quadrupleks deb ataladigan - to'rtta guaninning dumaloq raqsi.
Bunday tuzilmalarning paydo bo'lishi mumkinligi haqidagi birinchi maslahatlar Uotson va Krikning yutuq ishlaridan ancha oldin - 1910 yilda olingan. Keyin nemis kimyogari Ivar Bang DNK tarkibiy qismlaridan biri - guanosik kislota yuqori konsentratsiyalarda jellar hosil qilishini, DNKning boshqa komponentlarida esa bunday xususiyatga ega emasligini aniqladi.
1962 yilda rentgen nurlari diffraktsiya usulidan foydalanib, ushbu gelning hujayra tuzilishini o'rnatish mumkin edi. U to'rtta guanin qoldig'idan iborat bo'lib, bir-birini aylana shaklida bog'lab, xarakterli kvadrat hosil qiladi. Markazda bog'lanish metall ioni (Na, K, Mg) tomonidan quvvatlanadi. Xuddi shu tuzilmalar DNKda ko'p guanin bo'lsa, paydo bo'lishi mumkin. Bu tekis kvadratchalar (G-kvartetlar) ancha barqaror, zich tuzilmalarni (G-to'rtlik) hosil qilish uchun yig'iladi.
DNKning to'rtta alohida zanjiri to'rt zanjirli komplekslarga birlashtirilishi mumkin, ammo bu istisno. Ko'pincha nuklein kislotaning bir zanjiri oddiygina tugunga bog'lanib, xarakterli qalinlashuvlarni hosil qiladi (masalan, xromosomalarning uchlarida) yoki ikki zanjirli DNK guaninga boy bo'lgan ba'zi mintaqalarda mahalliy to'rtburchak hosil qiladi.
Eng ko'p o'rganilgani xromosomalarning uchlarida - telomerlarda va onkopromotorlarda to'rtliklarning mavjudligi. Biroq, hozirgi kunga qadar, inson xromosomalarida bunday DNKning lokalizatsiyasini to'liq tushunish ma'lum emas.
Chiziqli shakldagi DNKning barcha g'ayrioddiy tuzilmalari DNKning B shakliga nisbatan beqarordir. Biroq, DNK ko'pincha topologik stressning aylana shaklida mavjud bo'lsa, unda supercoiling deb ataladi. Bunday sharoitda kanonik bo'lmagan DNK tuzilmalari osongina hosil bo'ladi: Z-shakllari, xochlar va soch turmalari, H-shakllari, guanin kvadruplekslari va i-motiflar.
- Supero'ralgan shakl - pentoza-fosfat magistraliga zarar bermasdan, hujayra yadrosidan ajratilganda qayd etiladi. U o'ta o'ralgan yopiq halqalar shakliga ega. Supero'ralgan holatda DNK qo'sh spiral kamida bir marta "o'z-o'zidan o'raladi", ya'ni u kamida bitta superoilni o'z ichiga oladi (sakkizinchi raqam shaklini oladi).
- DNKning bo'shashgan holati bitta uzilish (bir ipning uzilishi) bilan kuzatiladi. Bunday holda, superkoillar yo'qoladi va DNK yopiq halqa shaklini oladi.
- DNK ning chiziqli shakli - qo'sh spiralning ikkita zanjiri uzilganda kuzatiladi.
DNKning uchinchi darajali tuzilishi
DNKning uchinchi darajali tuzilishi ikki zanjirli molekulaning bo'shliqda qo'shimcha buralishi - uning o'ta burilishi natijasida hosil bo'ladi. Eukaryotik hujayralardagi DNK molekulasining superkoillanishi, prokariotlardan farqli o'laroq, oqsillar bilan komplekslar shaklida amalga oshiriladi.
Deyarli barcha eukaryotik DNK yadrolarning xromosomalarida joylashgan bo'lib, uning ozgina qismi mitoxondriyalarda, o'simliklarda va plastidlarda mavjud. Eukaryotik hujayralar (shu jumladan, inson xromosomalari) xromosomalarining asosiy moddasi xromatin bo'lib, ikki zanjirli DNK, giston va giston bo'lmagan oqsillardan iborat.
Xromatin giston oqsillari
Gistonlar oddiy oqsillar bo'lib, xromatinning 50% ni tashkil qiladi. Hayvonlar va o'simliklarning barcha o'rganilgan hujayralarida gistonlarning beshta asosiy klassi topildi: H1, H2A, H2B, H3, H4, hajmi, aminokislotalar tarkibi va zaryad qiymati (har doim ijobiy).
Sutemizuvchilar gistoni H1 taxminan 215 aminokislotadan iborat bitta polipeptid zanjiridan iborat; boshqa gistonlarning o'lchamlari 100 dan 135 aminokislotagacha o'zgarib turadi. Ularning barchasi spirallangan va diametri taxminan 2,5 nm bo'lgan globulaga o'ralgan bo'lib, juda ko'p miqdordagi musbat zaryadlangan aminokislotalar lizin va argininni o'z ichiga oladi. Gistonlar atsetillangan, metillangan, fosforlangan, poli (ADP) -ribosillangan bo'lishi mumkin va H2A va H2B gistonlari ubiquitin bilan kovalent bog'langan. Tuzilishning shakllanishida va gistonlar tomonidan funktsiyalarni bajarishda bunday modifikatsiyalarning roli hali to'liq aniqlanmagan. Bu ularning DNK bilan o'zaro ta'sir qilish qobiliyati va genlar ta'sirini tartibga solish mexanizmlaridan birini ta'minlashi taxmin qilinadi.
Gistonlar DNK bilan asosan DNKning manfiy zaryadlangan fosfat guruhlari va gistonlarning musbat zaryadlangan lizin va arginin qoldiqlari oʻrtasida hosil boʻlgan ion bogʻlari (tuz koʻprigi) orqali oʻzaro taʼsir qiladi.
Giston bo'lmagan xromatin oqsillari
Gistonlardan farqli o'laroq, giston bo'lmagan oqsillar juda xilma-xildir. DNK bilan bog'langan giston bo'lmagan oqsillarning 590 tagacha turli fraktsiyalari ajratilgan. Ular kislotali oqsillar deb ham ataladi, chunki ularning tuzilishida kislotali aminokislotalar ustunlik qiladi (ular polianionlardir). Xromatin faolligini o'ziga xos tartibga solish giston bo'lmagan turli xil oqsillar bilan bog'liq. Masalan, DNK replikatsiyasi va ekspressiyasi uchun zarur bo'lgan fermentlar vaqtinchalik xromatin bilan bog'lanishi mumkin. Boshqa oqsillar, masalan, turli xil tartibga solish jarayonlarida ishtirok etib, DNK bilan faqat ma'lum to'qimalarda yoki differentsiatsiyaning ma'lum bosqichlarida bog'lanadi. Har bir oqsil ma'lum bir DNK nukleotidlar ketma-ketligini (DNK joyi) to'ldiradi. Bu guruhga quyidagilar kiradi:
- saytga xos sink barmoq oqsillari oilasi. Har bir sink barmog'i 5 juft nukleotiddan tashkil topgan ma'lum bir joyni taniydi.
- saytga xos oqsillar oilasi - homodimerlar. Bunday oqsilning DNK bilan aloqada bo'lgan qismi spiral-burilish-spiral tuzilishga ega.
- yuqori harakatchanlikli gel oqsillari (HMG oqsillari) doimiy ravishda xromatin bilan bog'liq bo'lgan tarkibiy va tartibga soluvchi oqsillar guruhidir. Ularning molekulyar og'irligi 30 kDa dan kam bo'lib, zaryadlangan aminokislotalarning yuqori miqdori bilan ajralib turadi. Past molekulyar og'irligi tufayli HMG oqsillari poliakrilamid gel elektroforezi paytida juda harakatchan.
- replikatsiya, transkripsiya va ta'mirlash fermentlari.
DNK va RNK sintezida ishtirok etuvchi strukturaviy, tartibga soluvchi oqsillar va fermentlar ishtirokida nukleosoma zanjiri oqsillar va DNKning yuqori kondensatsiyalangan kompleksiga aylanadi. Olingan struktura dastlabki DNK molekulasidan 10 000 marta qisqa.

Xromatin
Xromatin yadro DNKsi bilan oqsillar majmuasi va noorganik moddalar... Xromatinning katta qismi faol emas. U zich o'ralgan, kondensatsiyalangan DNKni o'z ichiga oladi. Bu heteroxromatin. Konstitutsiyaviy, genetik jihatdan faol bo'lmagan xromatin (sun'iy yo'ldosh DNK), ifodalanmagan hududlardan iborat va ixtiyoriy - bir qator avlodlarda faol bo'lmagan, ammo ma'lum sharoitlarda ifodalashga qodir.
Faol kromatin (euchromatin) kondensatsiyalanmagan, ya'ni. kamroq mahkamlangan. Turli hujayralarda uning miqdori 2 dan 11% gacha. Miya hujayralarida u eng ko'p - 10-11%, jigar hujayralarida - 3-4 va buyraklar - 2-3% ni tashkil qiladi. Euchromatinning faol transkripsiyasi qayd etilgan. Shu bilan birga, uning strukturaviy tashkil etilishi ma'lum bir turdagi organizmga xos bo'lgan bir xil DNK genetik ma'lumotlarini ixtisoslashgan hujayralarda turli usullarda qo'llash imkonini beradi.
Elektron mikroskopda xromatinning tasviri munchoqqa o'xshaydi: o'lchami taxminan 10 nm bo'lgan sferik qalinlashuvlar, ipli ko'priklar bilan ajratilgan. Bu globulyar qalinlashuvlar nukleosomalar deb ataladi. Nukleosoma xromatinning strukturaviy birligidir. Har bir nukleosomada uzunligi 146 ta tayanch juft bo'lgan, nukleosoma yadrosida 1,75 chap burilish hosil bo'lgan DNKning supero'ralgan segmenti mavjud. Nukleosoma yadrosi H2A, H2B, H3 va H4 gistonlaridan, har bir turdagi ikkita molekuladan tashkil topgan giston oktameridir (9-rasm), diametri 11 nm va qalinligi 5,7 nm bo'lgan diskka o'xshaydi. Beshinchi giston, H1, nukleosoma yadrosining bir qismi emas va giston oktamerida DNKni o'rash jarayonida ishtirok etmaydi. U qo'sh spiralning nukleosoma yadrosiga kirib, undan chiqadigan DNK bilan aloqa qiladi. Bular kortikal (bog'lovchi) DNK hududlari bo'lib, ularning uzunligi hujayra turiga qarab 40 dan 50 ta asosiy juftgacha o'zgaradi. Natijada, nukleosoma tarkibiga kiradigan DNK fragmentining uzunligi ham o'zgarib turadi (186 dan 196 juft nukleotidgacha).
Nukleosoma DNKning taxminan 90% ni o'z ichiga oladi, qolgan qismi bog'lovchi hisoblanadi. Nukleosomalar "jim" xromatinning bo'laklari bo'lib, bog'lovchi faol ekanligiga ishoniladi. Biroq, nukleosomalar ochilib, chiziqli bo'lishi mumkin. Ochilmagan nukleosomalar allaqachon faol xromatindir. Funktsiyaning strukturaga bog'liqligi ana shunday aniq namoyon bo'ladi. Taxmin qilish mumkinki, globulyar nukleosomalar tarkibida xromatin qancha ko'p bo'lsa, u shunchalik faol emas. Shubhasiz, turli hujayralarda tinch xromatinning teng bo'lmagan nisbati bunday nukleosomalar soni bilan bog'liq.
Elektron mikroskopik fotosuratlarda, izolyatsiya sharoitlari va cho'zilish darajasiga qarab, xromatin nafaqat qalinlashgan uzun ip - nukleosomalarning "boncuklari" sifatida, balki diametri 30 bo'lgan qisqaroq va zichroq fibrilla (tola) sifatida ham ko'rinishi mumkin. nm bo'lib, uning shakllanishi H1 gistonining DNK va H3 gistonining bog'lovchi hududiga bog'langan o'zaro ta'sirida kuzatiladi, bu esa diametri 30 nm bo'lgan solenoid hosil bo'lishi bilan oltita nukleosomadan iborat spiralning qo'shimcha burilishiga olib keladi. Bunday holda, giston oqsili bir qator genlarning transkripsiyasiga xalaqit berishi va shu bilan ularning faoliyatini tartibga solishi mumkin.
DNK ning gistonlar bilan yuqorida tavsiflangan o'zaro ta'siri natijasida o'rtacha diametri 2 nm va uzunligi 57 nm bo'lgan 186 ta asosiy juftlikdan iborat DNK qo'sh spiralining segmenti diametri 10 nm va bir spiralga aylanadi. uzunligi 5 nm. Keyinchalik bu spiralning diametri 30 nm bo'lgan tolaga siqilishi bilan kondensatsiya darajasi olti marta ortadi.
Oxir oqibat, DNK dupleksini besh giston bilan qadoqlash DNKning 50 marta kondensatsiyasiga olib keladi. Ammo bunday yuqori darajadagi kondensatsiya ham metafaza xromosomasida DNKning deyarli 50 000 - 100 000 marta zichlashishini tushuntirib bera olmaydi. Afsuski, xromatinning metafaza xromosomasiga qadar keyingi qadoqlanishi tafsilotlari hali ma'lum emas, shuning uchun bu jarayonning faqat umumiy xususiyatlarini ko'rib chiqish mumkin.
Xromosomalardagi DNKning siqilish darajalari
Har bir DNK molekulasi alohida xromosomaga o'ralgan. Insonning diploid hujayralarida 46 ta xromosoma mavjud bo'lib, ular hujayra yadrosida joylashgan. Hujayraning barcha xromosomalari DNKsining umumiy uzunligi 1,74 m, lekin xromosomalar to'plangan yadroning diametri millionlab marta kichikroqdir. Hujayra yadrosidagi xromosomalar va xromosomalardagi DNKning bunday ixcham to'plami DNK bilan ma'lum bir ketma-ketlikda o'zaro ta'sir qiluvchi turli xil giston va giston bo'lmagan oqsillar tomonidan ta'minlanadi (yuqoriga qarang). Xromosomalarda DNKning siqilishi uning chiziqli o'lchamlarini taxminan 10 000 marta - shartli ravishda 5 sm dan 5 mikrongacha kamaytirish imkonini beradi. Siqilishning bir necha darajalari mavjud (10-rasm).


- DNK qo'sh spiral diametri 2 nm va uzunligi bir necha sm bo'lgan manfiy zaryadlangan molekuladir.
- nukleosoma darajasi- xromatin elektron mikroskopda "munchoqlar" zanjiri - nukleosomalar - "ipda" ko'rinadi. Nukleosoma universal strukturaviy birlik bo'lib, u evromatin va geteroxromatinda, interfaza yadrosi va metafaza xromosomalarida joylashgan.
Siqilishning nukleosoma darajasi maxsus oqsillar - gistonlar tomonidan ta'minlanadi. Sakkizta musbat zaryadlangan giston domenlari manfiy zaryadlangan DNK molekulasi o'ralgan nukleosomaning yadrosini (yadrosini) tashkil qiladi. Bu 7 marta qisqarishga olib keladi, diametri esa 2 dan 11 nm gacha oshadi.
- solenoid darajasi
Xromosomalar tashkil etilishining solenoid darajasi nukleosoma filamentining buralishi va undan diametri 20-35 nm bo'lgan qalinroq fibrillalar - solenoidlar yoki superbidlar hosil bo'lishi bilan tavsiflanadi. Solenoidning balandligi 11 nm, har bir burilishda taxminan 6-10 nukleosoma mavjud. Solenoid o'rash superbiddan ko'ra ko'proq hisoblanadi, unga ko'ra diametri 20-35 nm bo'lgan xromatin fibrillasi har biri sakkiz nukleosomadan iborat bo'lgan granulalar zanjiri yoki superbidlardir. Solenoid darajasida DNKning chiziqli o'lchami 6-10 marta kamayadi, diametri 30 nm gacha oshadi.
- halqa darajasi
Loop darajasi taxminan 30-300 kb gacha bo'lgan halqalarni tashkil etuvchi, ma'lum DNK ketma-ketliklarini taniydigan va bog'laydigan giston bo'lmagan joyga xos DNKni bog'laydigan oqsillar tomonidan ta'minlanadi. Loop gen ifodasini ta'minlaydi, ya'ni E. loop nafaqat strukturaviy, balki funktsional shakllanishdir. Ushbu darajadagi qisqarish 20-30 marta sodir bo'ladi. Diametri 300 nm gacha oshadi. Sitologik preparatlarda amfibiya oositlarida "chiroq-cho'tka" tipidagi halqaga o'xshash tuzilmalarni ko'rish mumkin. Ko'rinishidan, bu halqalar o'ta o'ralgan va DNK domenlarini ifodalaydi, ular, ehtimol, xromatin transkripsiyasi va replikatsiyasi birliklariga mos keladi. Maxsus oqsillar halqalarning asoslarini va, ehtimol, ularning ba'zi ichki qismlarini mahkamlaydi. Halqa shaklidagi domen tashkiloti metafaza xromosomalaridagi xromatinning yuqori darajadagi spiral tuzilmalarga katlanishiga yordam beradi.
- domen darajasi
Xromosomalar tashkil etilishining domen darajasi etarli darajada o'rganilmagan. Ushbu darajada halqa domenlarining shakllanishi qayd etilgan - qalinligi 25-30 nm bo'lgan, tarkibida 60% protein, 35% DNK va 5% RNK bo'lgan iplar (fibrillalar) tuzilmalari hujayraning barcha fazalarida deyarli ko'rinmaydi. mitozdan tashqari tsikl va hujayra yadrosi bo'ylab tasodifiy taqsimlanadi. Sitologik preparatlarda amfibiya oositlarida "chiroq-cho'tka" tipidagi halqaga o'xshash tuzilmalarni ko'rish mumkin.
Loop domenlari o'z asoslarini yadro ichidagi oqsil matritsasiga o'rnatilgan biriktirish joylari deb ataladi, ko'pincha MAR / SAR ketma-ketliklari deb ataladi (MAR, ingliz matritsasi bilan bog'langan hududdan; SAR, inglizcha iskala biriktirma mintaqalaridan) - bir nechta DNK fragmentlari yuzlab uzunlikdagi tayanch juftlari, ular yuqori tarkibli (> 65%) A / T tayanch juftliklari bilan tavsiflanadi. Har bir domen replikatsiyaning yagona kelib chiqishiga ega bo'lib ko'rinadi va o'z-o'zidan o'ta o'ralgan birlik sifatida ishlaydi. Har qanday halqali domen ko'plab transkripsiya birliklarini o'z ichiga oladi, ularning ishlashi, ehtimol, muvofiqlashtirilgan - butun domen faol yoki nofaol holatda.
Domen darajasida, xromatinning ketma-ket qadoqlanishi natijasida DNKning chiziqli o'lchamlari taxminan 200 marta (700 nm) ga kamayadi.
- xromosoma darajasi
Xromosoma darajasida profilaktika xromosomasining birinchi metafazaga kondensatsiyasi giston bo'lmagan oqsillarning eksenel ramkasi atrofida halqali domenlarning siqilishi bilan sodir bo'ladi. Bu o'ta sarg'ish hujayradagi barcha H1 molekulalarining fosforlanishi bilan birga keladi. Natijada, metafaza xromosomasini qattiq spiralga o'ralgan qattiq o'ralgan solenoid halqalar sifatida tasvirlash mumkin. Odatiy inson xromosomasi 2600 tagacha halqadan iborat bo'lishi mumkin. Bunday strukturaning qalinligi 1400 nm (ikki xromatid) ga etadi, DNK molekulasi esa 104 marta qisqartiriladi, ya'ni. 5 sm gacha cho'zilgan DNK bilan 5 mkm.
Xromosoma funktsiyalari
Ekstraxromosoma mexanizmlari bilan o'zaro ta'sirda xromosomalar ta'minlaydi
- irsiy ma'lumotlarni saqlash
- uyali aloqani yaratish va saqlash uchun ushbu ma'lumotlardan foydalanish
- irsiy ma'lumotlarni o'qishni tartibga solish
- genetik materialning o'z-o'zidan ikki baravar ko'payishi
- genetik materialni ona hujayradan qizga o'tkazish.
Xromatin hududi faollashganda, ya'ni, dalillar mavjud. transkripsiya vaqtida undan avval giston H1, keyin esa giston okteti teskari ravishda chiqariladi. Bu xromatin dekondensatsiyasiga, 30 nanometrli xromatin fibrillasining 10 nanometrli filamentga ketma-ket o'tishiga va uning keyinchalik erkin DNK hududlariga tarqalishiga olib keladi, ya'ni. nukleosoma tuzilishini yo'qotish.
Hammamizga ma'lumki, insonning tashqi ko'rinishi, ba'zi odatlari va hatto kasalliklari meros bo'lib qoladi. Tirik mavjudot haqidagi bu ma'lumotlarning barchasi genlarda kodlangan. Xo'sh, bu maqol genlari qanday ko'rinishga ega, ular qanday ishlaydi va ular qayerda joylashgan?
Demak, har qanday odam yoki hayvonning barcha genlarining tashuvchisi DNKdir. Bu birikma 1869 yilda Iogan Fridrix Misher tomonidan kashf etilgan.Kimyoviy jihatdan DNK dezoksiribonuklein kislotadir. Bu qanday ma'nono bildiradi? Qanday qilib bu kislota sayyoramizdagi barcha hayotning genetik kodini olib yuradi?
Keling, DNK qayerda joylashganligini ko'rib chiqaylik. Inson hujayrasida turli funktsiyalarni bajaradigan ko'plab organellalar mavjud. DNK yadroda joylashgan. Yadro kichik organella bo'lib, u barcha genetik materialni - DNKni saqlaydigan maxsus membrana bilan o'ralgan.
DNK molekulasining tuzilishi qanday?
Avvalo, DNK nima ekanligini ko'rib chiqaylik. DNK - bu qurilish bloklari - nukleotidlardan tashkil topgan juda uzun molekula. Nukleotidlarning 4 turi mavjud - adenin (A), timin (T), guanin (G) va sitozin (C). Nukleotid zanjiri sxematik tarzda shunday ko'rinadi: GGAATCTAAG ... Bu DNK zanjiri bo'lgan nukleotidlar ketma-ketligi.DNK tuzilishi birinchi marta 1953 yilda Jeyms Uotson va Frensis Krik tomonidan ochilgan.
Bitta DNK molekulasida ikkita nukleotid zanjiri mavjud bo'lib, ular bir-birining atrofida spiral tarzda o'ralgan. Qanday qilib bu nukleotid zanjirlari bir-biriga yopishadi va spiralga aylanadi? Bu hodisa to'ldiruvchilik xususiyati bilan bog'liq. Komplementarlik deganda faqat ma'lum nukleotidlar (to'ldiruvchi) ikkita ipda bir-biriga qarama-qarshi joylashishi mumkin. Demak, adeninning qarshisida doimo timin, guaninning qarshisida esa faqat sitozin bo'ladi. Demak, guanin sitozinni, adenin esa timinni to'ldiruvchidir.Har xil iplardagi bir-biriga qarama-qarshi joylashgan bunday juft nukleotidlar ham komplementar deyiladi.
 Uni sxematik tarzda quyidagicha tasvirlash mumkin:
Uni sxematik tarzda quyidagicha tasvirlash mumkin:
G - C
T - A
T - A
C - G
Bu to'ldiruvchi juftliklar A - T va G - C hosil qiladi kimyoviy bog'lanish juftning nukleotidlari oʻrtasida boʻladi va G va S oʻrtasidagi bogʻlanish A va T ga qaraganda kuchliroqdir. Bogʻlanish qatʼiy ravishda komplementar asoslar orasida hosil boʻladi, yaʼni toʻldiruvchi boʻlmagan G va A oʻrtasida bogʻ hosil boʻlishi mumkin emas.
DNK "qadoqlash", DNK zanjiri qanday qilib xromosomaga aylanadi?
Nima uchun bu DNK nukleotid zanjirlari ham bir-birining atrofida aylanadi? Bu nima uchun kerak? Gap shundaki, nukleotidlar soni juda ko'p va bunday uzun zanjirlarni joylashtirish uchun juda ko'p joy talab etiladi. Shu sababli, ikkita DNK zanjirining bir-birining atrofida spiral burmasi mavjud. Bu hodisa spirallanish deyiladi. Spiralizatsiya natijasida DNK zanjirlari 5-6 marta qisqaradi.Ba'zi DNK molekulalari organizm tomonidan faol ishlatiladi, boshqalari esa kamdan-kam qo'llaniladi. Bunday kamdan-kam qo'llaniladigan DNK molekulalari spiralizatsiyadan tashqari, yanada ixchamroq "qadoqlash" dan o'tadi. Ushbu ixcham paket supercoiling deb ataladi va DNK zanjirini 25-30 marta qisqartiradi!
DNK zanjirining qadoqlanishi qanday sodir bo'ladi?
 Supercoiling uchun giston oqsillari qo'llaniladi, ular tashqi ko'rinishga va novda yoki ipli g'altakning tuzilishiga ega. Spirallashtirilgan DNK iplari ushbu "spirallar" - giston oqsillariga o'ralgan. Shunday qilib, uzun ip juda ixcham o'ralgan bo'lib, juda kam joy egallaydi.
Supercoiling uchun giston oqsillari qo'llaniladi, ular tashqi ko'rinishga va novda yoki ipli g'altakning tuzilishiga ega. Spirallashtirilgan DNK iplari ushbu "spirallar" - giston oqsillariga o'ralgan. Shunday qilib, uzun ip juda ixcham o'ralgan bo'lib, juda kam joy egallaydi. Agar u yoki bu DNK molekulasini ishlatish zarur bo'lsa, "echish" jarayoni sodir bo'ladi, ya'ni DNK zanjiri "spirtli" - giston oqsilidan (agar u o'ralgan bo'lsa) "ochiladi" va undan ochiladi. spiral ikkita parallel zanjirga aylanadi. Va DNK molekulasi shunday burilmagan holatda bo'lsa, undan kerakli genetik ma'lumotni o'qish mumkin. Bundan tashqari, genetik ma'lumotni o'qish faqat burilmagan DNK iplaridan sodir bo'ladi!
Yuqori o'ralgan xromosomalar to'plami deyiladi geterokromatin, va ma'lumotni o'qish uchun mavjud xromosomalar - evromatin.
Genlar nima, ularning DNK bilan aloqasi qanday?
Endi genlar nima ekanligini ko'rib chiqaylik. Ma'lumki, tanamizning qon guruhi, ko'z rangi, sochlari, terisi va boshqa ko'plab xususiyatlarini aniqlaydigan genlar mavjud. Gen - qat'iy belgilangan kombinatsiyada joylashgan ma'lum miqdordagi nukleotidlardan tashkil topgan DNKning qat'iy belgilangan bo'limi. DNKning qat'iy belgilangan hududida joylashishi ma'lum bir genga o'z o'rni berilganligini anglatadi va bu joyni o'zgartirish mumkin emas. Bunday qiyoslash o‘rinlidir: odam ma’lum bir ko‘chada, ma’lum bir uyda va xonadonda yashaydi va odam o‘zboshimchalik bilan boshqa uyga, xonadonga yoki boshqa ko‘chaga ko‘chishi mumkin emas. Gendagi nukleotidlarning ma'lum soni har bir genda ma'lum miqdordagi nukleotidlar mavjudligini va ko'p yoki kamroq bo'lishi mumkin emasligini anglatadi. Masalan, insulin ishlab chiqarish genining uzunligi 60 ta tayanch juft; oksitotsin gormoni ishlab chiqarishni kodlovchi gen - 370 ta asosiy juftlik. Nukleotidlarning qat'iy ketma-ketligi har bir gen uchun o'ziga xosdir va qat'iy belgilangan. Masalan, AATTAATA ketma-ketligi insulin ishlab chiqarishni kodlaydigan genning bir qismidir. Insulinni olish uchun aynan shunday ketma-ketlik qo'llaniladi, masalan, adrenalin olish uchun nukleotidlarning boshqa kombinatsiyasi qo'llaniladi. Nukleotidlarning faqat ma'lum bir birikmasi ma'lum bir "mahsulot" ni (adrenalin, insulin va boshqalar) kodlashini tushunish muhimdir. Bu "o'z o'rnida" turgan ma'lum miqdordagi nukleotidlarning noyob birikmasidir - bu gen.
Genlardan tashqari, DNK zanjirida "kodlanmagan ketma-ketliklar" joylashgan. Bunday kodlanmagan nukleotidlar ketma-ketligi genlarning ishini tartibga soladi, xromosomalarning spirallanishiga yordam beradi va genning boshlanishi va oxirini belgilaydi. Biroq, hozirgi kunga qadar ko'pchilik kodlanmagan ketma-ketliklarning roli noaniq bo'lib qolmoqda.
Xromosoma nima? Jinsiy xromosomalar
Individning genlari to'plamiga genom deyiladi. Tabiiyki, butun genomni bitta DNKga sig'dirib bo'lmaydi. Genom 46 juft DNK molekulasiga bo'linadi. Bir juft DNK molekulasi xromosoma deb ataladi. Shunday qilib, odamda 46 ta xromosomalar mavjud. Har bir xromosoma qat'iy belgilangan genlar to'plamini olib yuradi, masalan, 18-xromosomada ko'z rangini kodlaydigan genlar mavjud va hokazo. Xromosomalar uzunligi va shakli bo'yicha bir-biridan farq qiladi. Eng keng tarqalgan shakllar X yoki Y, lekin boshqalar ham bor. Odamda bir xil shakldagi ikkita xromosoma mavjud bo'lib, ular juft (juft) deb ataladi. Bunday farqlar tufayli barcha juftlashgan xromosomalar raqamlangan - ularning 23 tasi bor. Bu shuni anglatadiki, bir juft xromosomalar # 1, juft # 2, # 3 va boshqalar mavjud. Muayyan belgi uchun mas'ul bo'lgan har bir gen bir xil xromosomada joylashgan. Mutaxassislar uchun zamonaviy ko'rsatmalarda genning lokalizatsiyasi, masalan, quyidagicha ko'rsatilishi mumkin: 22 xromosoma, uzun qo'l.Xromosomalar o'rtasidagi farqlar qanday?
 Xromosomalar yana qanday farq qiladi? Uzun elka atamasi nimani anglatadi? X shaklidagi xromosomalarni oling. DNK zanjirlarining kesishishi qat'iy o'rtada (X) sodir bo'lishi mumkin yoki u markaziy bo'lmagan holda ham sodir bo'lishi mumkin. Agar DNK zanjirlarining bunday kesishishi markaziy tarzda sodir bo'lmasa, u holda kesishish nuqtasiga nisbatan ba'zi uchlari uzunroq, boshqalari esa mos ravishda qisqaroq bo'ladi. Bunday uzun uchlari odatda xromosomaning uzun qo'li deb ataladi va qisqa bo'lganlar mos ravishda qisqa qo'l deb ataladi. Y shaklidagi xromosomalarda ularning ko'p qismini uzun yelkalar egallaydi, qisqalari esa juda kichikdir (ular hatto sxematik rasmda ham ko'rsatilmagan).
Xromosomalar yana qanday farq qiladi? Uzun elka atamasi nimani anglatadi? X shaklidagi xromosomalarni oling. DNK zanjirlarining kesishishi qat'iy o'rtada (X) sodir bo'lishi mumkin yoki u markaziy bo'lmagan holda ham sodir bo'lishi mumkin. Agar DNK zanjirlarining bunday kesishishi markaziy tarzda sodir bo'lmasa, u holda kesishish nuqtasiga nisbatan ba'zi uchlari uzunroq, boshqalari esa mos ravishda qisqaroq bo'ladi. Bunday uzun uchlari odatda xromosomaning uzun qo'li deb ataladi va qisqa bo'lganlar mos ravishda qisqa qo'l deb ataladi. Y shaklidagi xromosomalarda ularning ko'p qismini uzun yelkalar egallaydi, qisqalari esa juda kichikdir (ular hatto sxematik rasmda ham ko'rsatilmagan). Xromosomalarning kattaligi har xil: eng kattasi № 1 va № 3 juft xromosomalar, eng kichiklari № 17, № 19 juft xromosomalardir.
Xromosomalar shakli va hajmiga qo'shimcha ravishda o'z vazifalarida ham farqlanadi. 23 ta juftlikdan 22 tasi somatik, 1 tasi jinsiy. Bu nima degani? Somatik xromosomalar shaxsning barcha tashqi belgilarini, ayniqsa uning xususiyatlarini aniqlaydi xulq-atvor reaktsiyalari, irsiy psixotip, ya'ni har bir alohida shaxsning barcha xususiyatlari va xususiyatlari. Bir juft jinsiy xromosomalar insonning jinsini aniqlaydi: erkak yoki ayol. Inson jinsiy xromosomalarining ikki turi mavjud - X (X) va Y (Y). Agar ular XX (X - X) kabi birlashtirilgan bo'lsa - bu ayol, agar XY (X - Y) bo'lsa - bizning oldimizda erkak bor.
Irsiy kasalliklar va xromosomalarning shikastlanishi
Biroq, genomning "buzilishi" sodir bo'ladi, keyin esa odamlarda genetik kasalliklar aniqlanadi. Masalan, 21 juft xromosomada ikkita emas, uchta xromosoma bo'lsa, odam Daun sindromi bilan tug'iladi.Genetik materialning ko'plab kichikroq "buzilishlari" mavjud bo'lib, ular kasallikning boshlanishiga olib kelmaydi, aksincha, yaxshi xususiyatlarni beradi. Genetik materialning barcha "parchalanishi" mutatsiyalar deb ataladi. Kasallikka yoki organizm xususiyatlarining yomonlashishiga olib keladigan mutatsiyalar salbiy, yangi foydali xususiyatlarning shakllanishiga olib keladigan mutatsiyalar esa ijobiy hisoblanadi.
Biroq, bugungi kunda odamlar azob chekayotgan kasalliklarning ko'pchiligiga nisbatan, bu irsiy kasallik emas, balki faqat moyillikdir. Misol uchun, shakar bolaning otasi tomonidan sekin so'riladi. Bu bolaning qandli diabet bilan tug'ilishini anglatmaydi, lekin bolaning moyilligi bo'ladi. Bu shuni anglatadiki, agar bola shirinlik va un mahsulotlarini suiiste'mol qilsa, unda qandli diabet paydo bo'ladi.
Bugungi kunda, deb atalmish predikativ dori. Ushbu tibbiy amaliyot doirasida odamda moyillik aniqlanadi (tegishli genlarni aniqlash asosida), so'ngra unga tavsiyalar beriladi - qanday parhezga rioya qilish kerak, ish va dam olish rejimini qanday qilib to'g'ri almashtirish kerak. kasal bo'lmaslik uchun.
DNKda kodlangan ma'lumotlarni qanday o'qish mumkin?
DNK tarkibidagi ma'lumotlarni qanday o'qiy olasiz? O'z tanasi undan qanday foydalanadi? DNKning o'zi matritsaning bir turi, lekin oddiy emas, lekin kodlangan. DNK matritsasidan ma'lumotni o'qish uchun u birinchi navbatda maxsus tashuvchiga - RNKga o'tkaziladi. RNK kimyoviy jihatdan ribonuklein kislotadir. Uning DNKdan farqi shundaki, u yadro membranasi orqali hujayraga o'ta oladi va DNK bu qobiliyatdan mahrum (u faqat yadroda bo'lishi mumkin). Kodlangan ma'lumot hujayraning o'zida ishlatiladi. Shunday qilib, RNK yadrodan hujayragacha kodlangan ma'lumotni tashuvchisidir.RNK qanday sintezlanadi, RNK yordamida oqsil qanday sintezlanadi?
 Ma'lumotni "o'qish", bo'shatish kerak bo'lgan DNK zanjirlari, ularga maxsus ferment - "quruvchi" yaqinlashadi va DNK zanjiriga parallel ravishda komplementar RNK zanjirini sintez qiladi. RNK molekulasi shuningdek, 4 turdagi nukleotidlardan - adenin (A), urasil (U), guanin (G) va sitozin (C) dan iborat. Bunda quyidagi juftliklar bir-birini to'ldiradi: adenin - urasil, guanin - sitozin. Ko'rib turganingizdek, DNKdan farqli o'laroq, RNK timin o'rniga urasildan foydalanadi. Ya'ni, "quruvchi" ferment quyidagicha ishlaydi: agar u DNK zanjirida A ni ko'rsa, u RNK zanjiriga Y ni, agar G bo'lsa, C ni biriktiradi va hokazo. Shunday qilib, transkripsiya paytida har bir faol gendan shablon hosil bo'ladi - yadro membranasidan o'tishi mumkin bo'lgan RNK nusxasi.
Ma'lumotni "o'qish", bo'shatish kerak bo'lgan DNK zanjirlari, ularga maxsus ferment - "quruvchi" yaqinlashadi va DNK zanjiriga parallel ravishda komplementar RNK zanjirini sintez qiladi. RNK molekulasi shuningdek, 4 turdagi nukleotidlardan - adenin (A), urasil (U), guanin (G) va sitozin (C) dan iborat. Bunda quyidagi juftliklar bir-birini to'ldiradi: adenin - urasil, guanin - sitozin. Ko'rib turganingizdek, DNKdan farqli o'laroq, RNK timin o'rniga urasildan foydalanadi. Ya'ni, "quruvchi" ferment quyidagicha ishlaydi: agar u DNK zanjirida A ni ko'rsa, u RNK zanjiriga Y ni, agar G bo'lsa, C ni biriktiradi va hokazo. Shunday qilib, transkripsiya paytida har bir faol gendan shablon hosil bo'ladi - yadro membranasidan o'tishi mumkin bo'lgan RNK nusxasi. Muayyan gen tomonidan kodlangan oqsil sintezi qanday sodir bo'ladi?
Yadrodan chiqqandan keyin RNK sitoplazmaga kiradi. Sitoplazmada allaqachon RNK matritsa sifatida maxsus ferment tizimlariga (ribosomalar) o'rnatilgan bo'lishi mumkin, ular RNK ma'lumotlariga asoslanib, tegishli protein aminokislotalar ketma-ketligini sintez qila oladi. Ma'lumki, oqsil molekulasi aminokislotalardan iborat. Ribosoma o'sib borayotgan oqsil zanjiriga qaysi aminokislota bog'lanishi kerakligini qanday aniqlashga muvaffaq bo'ladi? Bu triplet kod asosida amalga oshiriladi. Triplet kod RNK zanjirining uchta nukleotidlari ketma-ketligini bildiradi ( uchlik, masalan, HGH) bitta aminokislotani kodlaydi (bu holda, glisin). Har bir aminokislota ma'lum bir triplet tomonidan kodlangan. Shunday qilib, ribosoma tripletni "o'qiydi", RNKdagi ma'lumotlarni o'qiyotganda keyingi qaysi aminokislota biriktirilishi kerakligini aniqlaydi. Aminokislotalar zanjiri hosil bo'lganda, u ma'lum fazoviy shaklga ega bo'lib, unga yuklangan fermentativ, qurilish, gormonal va boshqa funktsiyalarni bajarishga qodir bo'lgan oqsilga aylanadi.Har qanday tirik organizm uchun protein gen mahsulotidir. Bu genlarning barcha xilma-xil xususiyatlarini, sifatlarini va tashqi ko'rinishlarini aniqlaydigan oqsillardir.
O'ng tomonda 2016 yil 23 aprelda Ginnesning rekordlar kitobiga kiritilgan Varna (Bolgariya) plyajidagi odamlardan qurilgan inson DNKsining eng katta spirali.
Deoksiribonuklein kislotasi. Umumiy ma'lumot
DNK (dezoksiribonuklein kislotasi) - bu hayotning o'ziga xos rejasi, irsiy ma'lumotlarga oid ma'lumotlarni o'z ichiga olgan murakkab kod. Bu murakkab makromolekula nasldan naslga irsiy genetik axborotni saqlash va uzatishga qodir. DNK har qanday tirik organizmning irsiyat va o'zgaruvchanlik kabi xususiyatlarini aniqlaydi. Unda kodlangan ma'lumotlar har qanday tirik organizmning rivojlanishi uchun butun dasturni belgilaydi. Genetik jihatdan o'ziga xos omillar insonning ham, boshqa organizmning ham butun hayotini oldindan belgilab beradi. Tashqi muhitning sun'iy yoki tabiiy ta'siri individual genetik xususiyatlarning umumiy jiddiyligiga ozgina ta'sir qilishi yoki dasturlashtirilgan jarayonlarning rivojlanishiga ta'sir qilishi mumkin.
Deoksiribonuklein kislotasi(DNK) - makromolekula (uchta asosiydan biri, qolgan ikkitasi RNK va oqsillar), u saqlash, avloddan avlodga o'tish va tirik organizmlarning rivojlanishi va faoliyatining genetik dasturini amalga oshirishni ta'minlaydi. DNK turli xil RNK va oqsillarning tuzilishi haqida ma'lumotni o'z ichiga oladi.
Eukaryotik hujayralarda (hayvonlar, o'simliklar va zamburug'lar) DNK xromosomalarning bir qismi sifatida hujayra yadrosida, shuningdek, ba'zi hujayra organellalarida (mitoxondriyalar va plastidlar) mavjud. Prokaryotik organizmlar hujayralarida (bakteriyalar va arxeyalar) hujayra membranasiga ichki tomondan nukleoid deb ataladigan dumaloq yoki chiziqli DNK molekulasi biriktirilgan. Ular va quyi eukariotlar (masalan, xamirturush) ham plazmidlar deb ataladigan kichik, avtonom, asosan aylana shaklidagi DNK molekulalariga ega.
Kimyoviy nuqtai nazardan, DNK takrorlanuvchi bloklar - nukleotidlardan tashkil topgan uzun polimer molekulasidir. Har bir nukleotid azotli asos, shakar (dezoksiriboza) va fosfat guruhidan iborat. Zanjirdagi nukleotidlar orasidagi bog'lanishlar dezoksiriboza tufayli hosil bo'ladi ( BILAN) va fosfat ( F) guruhlar (fosfodiester bog'lari).

Guruch. 2. Nukletid azotli asos, qand (dezoksiriboza) va fosfat guruhidan iborat.
Aksariyat hollarda (bir zanjirli DNKni o'z ichiga olgan ba'zi viruslardan tashqari) DNK makromolekulasi azotli asoslar bilan bir-biriga yo'naltirilgan ikkita zanjirdan iborat. Bu ikki zanjirli molekula spiral chiziq bo'ylab o'ralgan.
DNKda to'rt xil azotli asoslar mavjud (adenin, guanin, timin va sitozin). Zanjirlardan birining azotli asoslari ikkinchi zanjirning azotli asoslari bilan komplementarlik printsipiga ko'ra vodorod bog'lari bilan bog'langan: adenin faqat timin bilan bog'langan ( DA), guanin - faqat sitozin bilan ( G-C). Aynan shu juftliklar DNKning spiral "zinapoyasi" ning "chorbalarini" tashkil qiladi (qarang: 2, 3 va 4-rasm).

Guruch. 2. Azotli asoslar
Nukleotidlar ketma-ketligi haqida ma'lumotni "kodlash" imkonini beradi turli xil turlari RNK lar, ulardan eng muhimi informatsion yoki xabarchi (mRNK), ribosoma (rRNK) va transport (tRNK). Bu barcha turdagi RNKlar DNK ketma-ketligini transkripsiya jarayonida sintez qilingan RNK ketma-ketligiga nusxalash orqali DNK shablonida sintezlanadi va oqsillar biosintezida (translatsiya jarayoni) ishtirok etadi. Hujayra DNKsi kodlash ketma-ketliklaridan tashqari tartibga solish va tizimli funktsiyalarni bajaradigan ketma-ketlikni o'z ichiga oladi.

Guruch. 3. DNK replikatsiyasi
Kimyoviy DNK birikmalarining asosiy birikmalarining joylashuvi va bu birikmalar orasidagi miqdoriy munosabatlar irsiy ma'lumotlarning kodlanishini ta'minlaydi.
Ta'lim yangi DNK (replikatsiya)
- Replikatsiya jarayoni: DNK qo'sh spiralining ochilishi - DNK polimeraza tomonidan komplementar zanjirlarning sintezi - bittadan ikkita DNK molekulasining hosil bo'lishi.
- Fermentlar kimyoviy birikmalarning tayanch juftlari orasidagi bog‘lanishni buzganda, qo‘sh spiral ikki shoxchaga «ochiladi».
- Har bir filial yangi DNKning elementidir. Yangi tayanch juftliklar ota-ona filialidagi kabi ketma-ketlikda ulanadi.
Duplikatsiya tugagandan so'ng, ota-ona DNKsining kimyoviy birikmalaridan yaratilgan va u bilan bir xil genetik kodga ega bo'lgan ikkita mustaqil spiral hosil bo'ladi. Shunday qilib, DNK hujayradan hujayraga ma'lumotni hazm qila oladi.
Batafsil ma'lumot:
NUDLEIN KISLOTALARNING TUZILISHI

Guruch. 4 . Azotli asoslar: adenin, guanin, sitozin, timin
Deoksiribonuklein kislotasi(DNK) nuklein kislotalarni nazarda tutadi. Nuklein kislotalar monomerlari nukleotidlar bo'lgan tartibsiz biopolimerlar sinfidir.
NIKLEOTIDLAR dan iborat azotli asos besh uglerodli uglevod (pentoza) bilan birlashtirilgan - deoksiriboza(DNK holatida) yoki riboza(RNK holatida), bu fosfor kislotasi qoldig'i (H 2 PO 3 -) bilan birlashadi.
Azotli asoslar ikki xil bo'ladi: pirimidin asoslari - urasil (faqat RNKda), sitozin va timin, purin asoslari - adenin va guanin.

Guruch. 5. Nukleotidlarning tuzilishi (chapda), nukleotidning DNKdagi joylashuvi (pastda) va azotli asoslar turlari (o'ngda): pirimidin va purin.

Pentoza molekulasidagi uglerod atomlari 1 dan 5 gacha raqamlangan. Fosfat uchinchi va beshinchi uglerod atomlari bilan birlashadi. Shunday qilib nukleotidlar birlashib, nuklein kislota zanjirini hosil qiladi. Shunday qilib, biz DNK zanjirining 3 va 5 uchlarini ajratib olishimiz mumkin:

Guruch. 6. DNK zanjirining 3 «va 5» uchlarini ajratib olish
Ikkita DNK zanjiri hosil bo'ladi ikki tomonlama spiral... Spiraldagi bu zanjirlar qarama-qarshi yo'nalishda yo'naltirilgan. Turli DNK zanjirlarida azotli asoslar o'zaro bog'langan vodorod aloqalari... Adenin doimo timin bilan, sitozin esa guanin bilan birlashadi. U deyiladi bir-birini to'ldirish qoidasi(sm. bir-birini to'ldirish printsipi).
To'ldiruvchi qoida:
| A-T G-C |
Misol uchun, agar bizga ketma-ketlik bilan DNK zanjiri berilsa
3'- ATGTCCTAGCTGCTCG - 5 ',
keyin ikkinchi zanjir unga to'ldiruvchi bo'ladi va teskari yo'nalishda - 5'-uchidan 3'-uchiga yo'naltiriladi:
5'-TACAGGATCGACGAGC-3 '.

Guruch. 7. DNK molekulasi zanjirlarining yo‘nalishi va azotli asoslarning vodorod bog‘lari yordamida bog‘lanishi.
DNK replikatsiyasi
DNK replikatsiyasi matritsa sintezi yordamida DNK molekulasini ikki barobar oshirish jarayonidir. Ko'p hollarda tabiiy DNK replikatsiyasiastarDNK sintezi uchun qisqa parcha (qayta yaratilgan). Bunday ribonukleotid primeri primaza fermenti (prokaryotlarda DNK primazasi, eukariotlarda DNK polimeraza) tomonidan yaratiladi va keyinchalik odatda ta'mirlash funktsiyalarini bajaradigan dezoksiribonukleotid polimeraza bilan almashtiriladi (DNK molekulasidagi kimyoviy shikastlanish va uzilishlarni to'g'rilaydi).
Replikatsiya yarim konservativ mexanizm bilan sodir bo'ladi. Bu shuni anglatadiki, DNK ning qo'sh spirali bo'shatiladi va uning har bir zanjirida to'ldiruvchilik printsipiga ko'ra yangi ip tugallanadi. Shunday qilib, qiz DNK molekulasi ota-molekuladan bitta zanjirni va bitta yangi sintezlangan zanjirni o'z ichiga oladi. Replikatsiya asosiy zanjirning 3 “5” oxirigacha bo'lgan yo'nalishda sodir bo'ladi.

Guruch. 8. DNK molekulasining replikatsiyasi (ikki marta ko'payishi).
DNK sintezi- bu birinchi qarashda ko'rinadigan darajada murakkab jarayon emas. Agar siz bu haqda o'ylab ko'rsangiz, avval sintez nima ekanligini aniqlab olishingiz kerak. Bu biror narsani birlashtirish jarayonidir. Yangi DNK molekulasining shakllanishi bir necha bosqichda sodir bo'ladi:
1) replikatsiya vilkasi oldida joylashgan DNK topoizomerazasi DNKni ochish va ochishni osonlashtirish uchun uni kesib tashlaydi.
2) DNK helikazasi, topoizomerazdan keyin DNK spiralining "burilish" jarayoniga ta'sir qiladi.
3) DNK-bog'lovchi oqsillar DNK zanjirlarini bog'lashni amalga oshiradi, shuningdek, ularning bir-biriga yopishib qolishiga yo'l qo'ymasdan, ularning barqarorlashuvini amalga oshiradi.
4) DNK polimeraza d(delta) , replikativ vilkaning harakat tezligi bilan muvofiqlashtirilgan, sintezni amalga oshiradiyetakchizanjirlar sho'ba korxonasi Shablondagi 5 "→ 3" yo'nalishidagi DNK onalik DNK zanjiri o'zining 3 "-uchidan 5" uchigacha yo'nalishda (tezlik sekundiga 100 ta asosiy juftgacha). Bu voqealar haqida onalik DNK zanjirlari cheklangan.

Guruch. 9. DNK replikatsiyasi jarayonining sxematik tasviri: (1) orqada qolgan zanjir (kechikuvchi zanjir), (2) yetakchi zanjir (etakchi zanjir), (3) DNK polimeraza a (Pola), (4) DNK ligaza, (5) RNK -primer, (6) Primaza, (7) Okazaki fragmenti, (8) DNK polimeraza d (Polo), (9) Helikaz, (10) Bir zanjirli DNKni bog'lovchi oqsillar, (11) Topoizomeraz.
Quyida qiz DNKning orqada qolgan zanjirining sintezi tasvirlangan (qarang. Sxema replikatsiya vilkasi va replikatsiya fermenti funktsiyasi)
DNK replikatsiyasini ko'proq vizual tushuntirish uchun qarang
5) Ota-molekulaning boshqa ipi echib, barqarorlashgandan so'ng darhol;DNK polimeraza a(alfa)va 5 "→ 3" yo'nalishi bo'yicha primer (RNK primeri) - uzunligi 10 dan 200 nukleotidgacha bo'lgan DNK shablonidagi RNK ketma-ketligini sintez qiladi. Shundan so'ng, fermentDNK zanjiridan chiqariladi.
Ning o'rniga DNK polimerazaα
primerning 3 "uchiga biriktiriladi DNK polimerazaε
.
6)
DNK polimerazaε
(epsilon) go'yo u primerni uzaytirishda davom etadi, lekin substrat sifatida u ko'madideoksiribonukleotidlar(150-200 nukleotid miqdorida). Natijada, ikki qismdan qattiq ip hosil bo'ladi -RNK(ya'ni, primer) va DNK.
DNK polimeraza eoldingi primerga mos kelguncha ishlaydiOkazaki parchasi(bir oz oldin sintez qilingan). Keyin bu ferment zanjirdan chiqariladi.
7) DNK polimeraza b(beta) o'rniga o'rnidan turadiDNK polimeraza e,bir xil yo'nalishda (5 "→ 3") harakat qiladi va ularning o'rniga deoksiribonukleotidlarni joylashtirganda primer ribonukleotidlarni olib tashlaydi. Enzim primerni to'liq olib tashlamaguncha ishlaydi, ya'ni. deoksiribonukleotidga qadar (hatto ilgari sintez qilingan).DNK polimeraza e). Ferment o'z ishining natijasini va uning oldidagi DNKni bog'lay olmaydi, shuning uchun u zanjirni tark etadi.
Natijada, ona ipining matritsasida qiz DNKning bir qismi "yotadi". U deyiladiOkazaki parchasi.
8) DNK ligazasi ikkita qo'shni tikuv Okazaki parchalari , ya'ni. 5 "-sintezlangan segmentning oxiriDNK polimeraza e,va 3 "-sxemaning oxiri, o'rnatilganDNK polimerazaβ .
RNK TUZILISHI
Ribonuklein kislotasi(RNK) barcha tirik organizmlarning hujayralarida joylashgan uchta asosiy makromolekulalardan biri (qolgan ikkitasi DNK va oqsillar).
Xuddi DNK singari, RNK ham har bir bo'g'in deb ataladigan uzun zanjirdan iborat nukleotid... Har bir nukleotid azotli asos, riboza shakar va fosfat guruhidan iborat. Biroq, DNKdan farqli o'laroq, RNK odatda ikkita emas, balki bitta zanjirga ega. RNKdagi pentoza deoksiriboza emas, riboza bilan ifodalanadi (riboza ikkinchi uglevod atomida qo'shimcha gidroksil guruhiga ega). Nihoyat, DNK azotli asoslar tarkibida RNK dan farq qiladi: timin o'rniga ( T) urasil ( U) Bu ham adeninni to'ldiruvchi hisoblanadi.
Nukleotidlar ketma-ketligi RNKga genetik ma'lumotni kodlash imkonini beradi. Barcha hujayrali organizmlar oqsil sintezini dasturlash uchun RNK (mRNK) dan foydalanadi.
Hujayra RNKlari deb ataladigan jarayon orqali hosil bo'ladi transkripsiya , ya'ni maxsus fermentlar tomonidan amalga oshiriladigan DNK matritsasida RNK sintezi - RNK polimerazalari.
Keyin xabarchi RNKlar (mRNKlar) chaqirilgan jarayonda ishtirok etadilar eshittirish, bular. ribosomalar ishtirokida mRNK matritsasida oqsil sintezi. Boshqa RNKlar transkripsiyadan so'ng kimyoviy modifikatsiyadan o'tadi va ikkilamchi va uchinchi darajali tuzilmalar hosil bo'lgandan so'ng ular RNK turiga qarab funktsiyalarni bajaradilar.

Guruch. 10. Azotli asosda DNK va RNK o'rtasidagi farq: RNKda timin (T) o'rniga adeninni ham to'ldiruvchi urasil (U) mavjud.
TRANSKRIPSIYA
Bu DNK shablonida RNK sintezi jarayonidir. DNK saytlardan birida ochiladi. Iplardan biri RNK molekulasiga ko'chirilishi kerak bo'lgan ma'lumotlarni o'z ichiga oladi - bu zanjir kodlash zanjiri deb ataladi. Kodlovchiga to'ldiruvchi ikkinchi DNK zanjiri shablon deb ataladi. Shablon zanjirida 3 '- 5' yo'nalishi bo'yicha (DNK zanjiri bo'ylab) transkripsiya jarayonida komplementar RNK zanjiri sintezlanadi. Shunday qilib, kodlash zanjirining RNK nusxasi yaratiladi.
![]()
Guruch. 11. Transkripsiyaning sxematik tasviri
Misol uchun, agar bizga kodlash chizig'ining ketma-ketligi berilsa
3'- ATGTCCTAGCTGCTCG - 5 ',
keyin, to'ldiruvchilik qoidasiga ko'ra, matritsa zanjiri ketma-ketlikni olib yuradi
5'- TACAGGATCGACGAGC- 3 ',
va undan sintez qilingan RNK ketma-ketlikdir
Translyatsiya
Mexanizmni ko'rib chiqing oqsil sintezi RNK matritsasida, shuningdek, genetik kod va uning xususiyatlari. Shuningdek, aniqlik uchun quyidagi havoladan foydalanib, tirik hujayrada sodir bo'ladigan transkripsiya va tarjima jarayonlari haqida qisqacha video tomosha qilishni tavsiya etamiz:

Guruch. 12. Protein sintez jarayoni: DNK RNKni, RNK oqsilni kodlaydi
GENETIK KOD
Genetik kod- nukleotidlar ketma-ketligi yordamida oqsillarning aminokislotalar ketma-ketligini kodlash usuli. Har bir aminokislota uchta nukleotidlar ketma-ketligi bilan kodlangan - kodon yoki triplet.
Ko'pgina pro- va eukariotlarga xos bo'lgan genetik kod. Jadvalda barcha 64 ta kodon ko'rsatilgan va tegishli aminokislotalar ko'rsatilgan. Asosiy tartib mRNKning 5-“3” oxirigacha.
Jadval 1. Standart genetik kod
|
1 yo'q |
2-tayanch |
3 yo'q |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe / F) |
U C U |
(Ser / S) |
U A U |
(Tyr / Y) |
U G U |
(Cys / C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu / L) |
U C A |
U A A |
Kodonni to'xtating ** |
U G A |
Kodonni to'xtating ** |
A |
||
|
U U G |
U C G |
U A G |
Kodonni to'xtating ** |
U G G |
(Trp / Vt) |
G |
|||
|
C |
C U U |
C C U |
(Pro / P) |
C A U |
(Uning / H) |
C G U |
(Arg / R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln / Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile / I) |
A C U |
(Thr / T) |
A A U |
(Asn / N) |
A G U |
(Ser / S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys / K) |
A G A |
A |
||||
|
A U G |
(Met / M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val / V) |
G C U |
(Ala / A) |
G A U |
(Asp / D) |
G G U |
(Gly / G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu / E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Uchlik orasida "tinish belgilari" vazifasini bajaradigan 4 ta maxsus ketma-ketlik mavjud:
- * Uchlik AVG, shuningdek, metioninni kodlovchi, deyiladi kodonni boshlang... Oqsil molekulasining sintezi shu kodondan boshlanadi. Shunday qilib, oqsil sintezi jarayonida ketma-ketlikdagi birinchi aminokislota har doim metionin bo'ladi.
- ** Uch nafar UAA, UAG va UGA deyiladi kodonlarni to'xtatish va bitta aminokislotani kodlamang. Ushbu ketma-ketlikda oqsil sintezi to'xtaydi.
Genetik kodning xususiyatlari
1. Uchlik... Har bir aminokislota uchta nukleotidlar ketma-ketligi bilan kodlangan - triplet yoki kodon.
2. Davomiylik... Tripletlar orasida qo'shimcha nukleotidlar yo'q, ma'lumot doimiy ravishda o'qiladi.
3. Bir-biriga yopishmaslik... Bitta nukleotid bir vaqtning o'zida ikkita tripletga kira olmaydi.
4. Aniqlik... Bitta kodon faqat bitta aminokislotani kodlashi mumkin.

5. Degeneratsiya... Bitta aminokislota bir nechta turli kodonlar tomonidan kodlanishi mumkin.

6. Ko'p qirralilik... Genetik kod barcha tirik organizmlar uchun bir xil.
Misol. Bizga kodlash zanjirining ketma-ketligi berilgan:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Matritsa zanjiri quyidagi ketma-ketlikka ega bo'ladi:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Endi biz ushbu zanjirdan informatsion RNKni "sintezlaymiz":
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Protein sintezi 5 "→ 3" yo'nalishi bo'yicha ketadi, shuning uchun biz genetik kodni "o'qish" uchun ketma-ketlikni aylantirishimiz kerak:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
Endi AUG start kodonini topamiz:
5’- AU AVG CUAGCUGCACGUUAGCC- 3’.
Keling, ketma-ketlikni uchlikka ajratamiz:
shunday eshitiladi: DNKdan olingan ma'lumotlar RNKga (transkripsiya), RNKdan - oqsilga (translyatsiya) o'tkaziladi. DNK replikatsiya yo'li bilan ham ko'paytirilishi mumkin va teskari transkripsiya jarayoni ham mumkin, DNK RNK shablonidan sintez qilinganda, lekin bu jarayon asosan viruslar uchun xosdir.

Guruch. 13. Markaziy dogma molekulyar biologiya
GENOM: GENLAR va XROMOSOMLAR
(umumiy tushunchalar)
Genom - organizmning barcha genlarining yig'indisi; uning to'liq xromosoma to'plami.
"Genom" atamasi 1920 yilda G.Vinkler tomonidan bitta biologik turga mansub organizmlar xromosomalarining haploid to'plamida joylashgan genlar to'plamini tavsiflash uchun taklif qilingan. Ushbu atamaning asl ma'nosi shuni ko'rsatdiki, genom tushunchasi, genotipdan farqli o'laroq, alohida shaxsning emas, balki butun turning genetik xususiyati hisoblanadi. Molekulyar genetikaning rivojlanishi bilan bu atamaning ma'nosi o'zgardi. Ma'lumki, ko'pchilik organizmlarda genetik ma'lumotlarning tashuvchisi bo'lgan va shuning uchun genomning asosini tashkil etuvchi DNK so'zning zamonaviy ma'nosida nafaqat genlarni o'z ichiga oladi. Katta qismi Eukaryotik hujayralarning DNKsi oqsillar va nuklein kislotalar haqida ma'lumotga ega bo'lmagan kodlanmagan ("ortiqcha") nukleotidlar ketma-ketligi bilan ifodalanadi. Shunday qilib, har qanday organizm genomining asosiy qismi uning haploid xromosomalar to'plamining butun DNKsidir.
Genlar - polipeptidlar va RNK molekulalarini kodlaydigan DNK molekulalarining bo'limlari
O'tgan asrda bizning genlar haqidagi tushunchamiz sezilarli darajada o'zgardi. Ilgari genom xromosomaning bitta xususiyatni kodlaydigan yoki aniqlaydigan qismi deb atalgan. fenotipik ko'z rangi kabi (ko'rinadigan) xususiyat.
1940 yilda Jorj Bidl va Edvard Tatem genning molekulyar ta'rifini taklif qilishdi. Olimlar qo'ziqorin sporalarini davolashdi Neyrospora crassa Rentgen nurlari va DNK ketma-ketligini o'zgartirishga olib keladigan boshqa vositalar ( mutatsiyalar), va ba'zi o'ziga xos fermentlarni yo'qotgan qo'ziqorinning mutant shtammlarini topdi, bu esa ba'zi hollarda butun metabolik yo'lning buzilishiga olib keldi. Beadle va Tatem gen - bu bitta fermentni belgilaydigan yoki kodlaydigan genetik materialning bir qismi degan xulosaga kelishdi. Gipoteza shunday paydo bo'ldi "Bir gen - bitta ferment"... Keyinchalik bu kontseptsiyani aniqlash uchun kengaytirildi "Bir gen - bitta polipeptid", chunki ko'p genlar fermentlar bo'lmagan oqsillarni kodlaydi va polipeptid murakkab protein kompleksining pastki birligi bo'lishi mumkin.
Shaklda. 14 - DNKdagi nukleotidlarning tripletlari polipeptidni, mRNK vositachiligida oqsilning aminokislotalar ketma-ketligini qanday aniqlashining diagrammasi. DNK zanjirlaridan biri mRNK sintezi uchun shablon rolini o'ynaydi, uning nukleotid tripletlari (kodonlari) DNK tripletlarini to'ldiradi. Ba'zi bakteriyalar va ko'plab eukaryotlarda kodlash ketma-ketligi kodlanmaydigan hududlar (deb ataladigan) tomonidan uziladi. intronlar).
Zamonaviy biokimyoviy gen ta'rifi yanada aniqroq. Genlar DNKning barcha bo'limlari bo'lib, yakuniy mahsulotlarning birlamchi ketma-ketligini kodlaydi, ular tarkibiga strukturaviy yoki katalitik funktsiyaga ega bo'lgan polipeptidlar yoki RNK kiradi.
Genlar bilan bir qatorda DNKda faqat tartibga solish funktsiyasini bajaradigan boshqa ketma-ketliklar ham mavjud. Normativ ketma-ketliklar genlarning boshlanishi yoki oxirini bildirishi, transkripsiyaga ta'sir qilishi yoki replikatsiya yoki rekombinatsiya boshlangan joyni ko'rsatishi mumkin. Ba'zi genlar turli yo'llar bilan ifodalanishi mumkin, bir xil DNK bo'lagi turli xil mahsulotlarning shakllanishi uchun shablon bo'lib xizmat qiladi.
Biz taxminan hisoblashimiz mumkin minimal gen hajmi o'rtacha proteinni kodlash. Polipeptid zanjiridagi har bir aminokislota uchta nukleotidlar ketma-ketligi sifatida kodlangan; bu tripletlarning (kodonlarning) ketma-ketligi berilgan gen tomonidan kodlangan polipeptiddagi aminokislotalar zanjiriga mos keladi. 350 ta aminokislota qoldig'idan iborat polipeptid zanjiri (o'rta zanjir) 1050 bp ketma-ketlikka to'g'ri keladi. ( tayanch juftliklar). Biroq, ko'pgina eukaryotik genlar va ba'zi prokaryotik genlar DNK segmentlari tomonidan uzilib qoladi. axborot tashuvchilar oqsil haqida, va shuning uchun oddiy hisob ko'rsatadi ancha uzoq bo'lib chiqadi.
Bitta xromosomada nechta gen bor?
 Guruch. 15. Prokarit (chapda) va eukaryotik hujayralardagi xromosomalarning ko'rinishi. Gistonlar ikkita asosiy funktsiyani bajaradigan yadro oqsillarining keng sinfidir: ular yadrodagi DNK zanjirlarini o'rashda va transkripsiya, replikatsiya va ta'mirlash kabi yadro jarayonlarini epigenetik tartibga solishda ishtirok etadilar.
Guruch. 15. Prokarit (chapda) va eukaryotik hujayralardagi xromosomalarning ko'rinishi. Gistonlar ikkita asosiy funktsiyani bajaradigan yadro oqsillarining keng sinfidir: ular yadrodagi DNK zanjirlarini o'rashda va transkripsiya, replikatsiya va ta'mirlash kabi yadro jarayonlarini epigenetik tartibga solishda ishtirok etadilar.
Prokariotlarning DNKsi oddiyroq: ularning hujayralari yadroga ega emas, shuning uchun DNK bevosita sitoplazmada nukleoid shaklida joylashgan.
 Ma'lumki, bakteriya hujayralari ixcham tuzilishga - nukleoidga o'ralgan DNK zanjiri ko'rinishidagi xromosomaga ega. Prokaryot xromosomasi Escherichia coli, genomi to'liq dekodlangan, aylana shaklidagi DNK molekulasidir (aslida u emas. to'g'ri doira, balki boshi va oxiri bo'lmagan halqa), 4 639 675 bp dan iborat. Bu ketma-ketlikda oqsillar uchun taxminan 4300 gen va barqaror RNK molekulalari uchun 157 gen mavjud. V inson genomi taxminan 3,1 milliard tayanch jufti, bu 24 xil xromosomalarda joylashgan 29 000 ga yaqin genga mos keladi.
Ma'lumki, bakteriya hujayralari ixcham tuzilishga - nukleoidga o'ralgan DNK zanjiri ko'rinishidagi xromosomaga ega. Prokaryot xromosomasi Escherichia coli, genomi to'liq dekodlangan, aylana shaklidagi DNK molekulasidir (aslida u emas. to'g'ri doira, balki boshi va oxiri bo'lmagan halqa), 4 639 675 bp dan iborat. Bu ketma-ketlikda oqsillar uchun taxminan 4300 gen va barqaror RNK molekulalari uchun 157 gen mavjud. V inson genomi taxminan 3,1 milliard tayanch jufti, bu 24 xil xromosomalarda joylashgan 29 000 ga yaqin genga mos keladi.
Prokaryotlar (bakteriyalar).
 Bakteriya E. coli bitta ikkita zanjirli dumaloq DNK molekulasiga ega. U 4 639 675 bp dan iborat. va uzunligi taxminan 1,7 mm ga etadi, bu hujayraning o'zi uzunligidan oshadi E. coli taxminan 850 marta. Nukleoiddagi katta doiraviy xromosomadan tashqari, ko'plab bakteriyalarda sitozolda erkin joylashgan bir yoki bir nechta kichik dumaloq DNK molekulalari mavjud. Bunday ekstraxromosoma elementlari deyiladi plazmidlar(16-rasm).
Bakteriya E. coli bitta ikkita zanjirli dumaloq DNK molekulasiga ega. U 4 639 675 bp dan iborat. va uzunligi taxminan 1,7 mm ga etadi, bu hujayraning o'zi uzunligidan oshadi E. coli taxminan 850 marta. Nukleoiddagi katta doiraviy xromosomadan tashqari, ko'plab bakteriyalarda sitozolda erkin joylashgan bir yoki bir nechta kichik dumaloq DNK molekulalari mavjud. Bunday ekstraxromosoma elementlari deyiladi plazmidlar(16-rasm).
Aksariyat plazmidlar atigi bir necha ming tayanch juftlikdan iborat bo'lib, ba'zilarida 10 000 bp dan ortiq. Ular genetik ma'lumotni olib yuradilar va ona hujayraning bo'linishi paytida qiz hujayralarga kiradigan qiz plazmidlarning shakllanishi bilan ko'payadilar. Plazmidlar nafaqat bakteriyalarda, balki xamirturush va boshqa zamburug'larda ham uchraydi. Ko'p hollarda plazmidlar xost hujayralariga hech qanday afzallik bermaydi va ularning yagona vazifasi mustaqil ko'payishdir. Biroq, ba'zi plazmidlar xost uchun foydali genlarni olib yuradi. Masalan, plazmidlar tarkibidagi genlar bakteriya hujayralariga antibakterial vositalarga qarshilik ko'rsatishi mumkin. B-laktamaza genini tashuvchi plazmidlar penitsillin va amoksitsillin kabi b-laktam antibiotiklariga qarshilik ko'rsatadi. Plazmidlar antibiotiklarga chidamli hujayralardan bir xil yoki boshqa turdagi bakteriyalarning boshqa hujayralariga o'tkazilishi mumkin, bu hujayralarni ham chidamli qiladi. Antibiotiklardan intensiv foydalanish patogen bakteriyalar orasida antibiotiklarga chidamliligini kodlovchi plazmidlarning (shuningdek, shunga o'xshash genlarni kodlovchi transpozonlar) tarqalishiga yordam beruvchi kuchli selektiv omil bo'lib, bir nechta antibiotiklarga chidamli bakterial shtammlarning paydo bo'lishiga olib keladi. Shifokorlar antibiotiklarni keng qo'llash xavfini tushunishni boshlaydilar va ularni faqat shoshilinch zarurat tug'ilganda yozadilar. Shu kabi sabablarga ko'ra qishloq xo'jaligi hayvonlarini davolash uchun antibiotiklardan keng foydalanish cheklangan.
Shuningdek qarang: Ravin N.V., Shestakov S.V. Prokaryotlar genomi // Vavilov jurnali Genetika va seleksiya, 2013. V. 17. No 4/2. S. 972-984.
Eukariotlar.
2-jadval. Ayrim organizmlarning DNKsi, genlari va xromosomalari
|
Umumiy DNK, p.n. |
Xromosoma soni * |
Genlarning taxminiy soni |
|
|
Escherichia coli(bakteriya) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(xamirturush) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(nematod) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(o'simlik) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(meva chivinlari) |
120 367 260 |
20 000 |
|
|
Oryza sativa(guruch) |
480 000 000 |
57 000 |
|
|
Mushak mushaklari(sichqoncha) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens(Odam) |
3 070 128 600 |
29 000 |
Eslatma. Ma'lumotlar doimiy ravishda yangilanadi; qo'shimcha ma'lumot olish uchun individual genomik loyihalarga bag'ishlangan saytlarga murojaat qiling
* Xamirturushdan tashqari barcha eukariotlar uchun xromosomalarning diploid to'plami berilgan. Diploid to'plam xromosomalar (yunoncha diploos- qo'sh va eidos- turlardan) - ikki xil xromosomalar to'plami(2n), ularning har biri gomologga ega.
** Gaploid to'plami. Yovvoyi xamirturush shtammlari odatda sakkizta (oktaploid) yoki undan ko'p bunday xromosomalarga ega.
*** Ikkita X xromosomali ayollar uchun. Erkaklarda X xromosoma bor, lekin Y yo'q, ya'ni faqat 11 xromosoma mavjud.
Eng kichik eukariotlardan biri bo'lgan xamirturush hujayrasi hujayradan 2,6 baravar ko'p DNKga ega. E. coli(2-jadval). Meva chivinlari hujayralari Drosophila, genetik tadqiqotning klassik ob'ekti, DNK 35 baravar ko'p, inson hujayralari esa hujayralarga qaraganda taxminan 700 baravar ko'p DNKni o'z ichiga oladi. E. coli. Ko'pgina o'simliklar va amfibiyalarda ko'proq DNK mavjud. Eukaryotik hujayralarning genetik materiali xromosomalar shaklida tashkil etilgan. Xromosomalarning diploid to'plami (2 n) organizmning turiga bog'liq (2-jadval).
Masalan, odamning somatik hujayrasida 46 ta xromosoma mavjud ( guruch. 17). Eukaryotik hujayraning har bir xromosomasi, rasmda ko'rsatilganidek. 17, a, tarkibida bitta juda katta ikki zanjirli DNK molekulasi mavjud. Yigirma to'rtta odam xromosomalari (22 juft xromosoma va ikkita jinsiy xromosoma X va Y) uzunligi bo'yicha 25 martadan ko'proq farq qiladi. Har bir eukaryotik xromosoma o'ziga xos genlarni o'z ichiga oladi.

Guruch. 17. Eukaryotik xromosomalar.a- odam xromosomasidan bir juft bog'langan va kondensatsiyalangan opa-singil xromatidlar. Ushbu shaklda eukaryotik xromosomalar replikatsiyadan keyin va mitoz paytida metafazada qoladi. b- kitob mualliflaridan birining leykotsitidan xromosomalarning to'liq to'plami. Har bir oddiy odam somatik hujayrasida 46 ta xromosoma mavjud.

DNKning irsiy materialni saqlash va uzatish matritsasi sifatidagi hajmi va vazifasi ushbu molekulaning tashkil etilishida maxsus strukturaviy elementlarning mavjudligini tushuntiradi. Yuqori organizmlarda DNK xromosomalar orasida taqsimlanadi.
Organizmning DNK (xromosomalari) to'plamiga genom deyiladi. Xromosomalar hujayra yadrosida joylashgan bo'lib, xromatin deb ataladigan tuzilishni hosil qiladi. Xromatin 1: 1 nisbatda DNK va asosiy oqsillar (gistonlar) majmuasidir. DNK uzunligi odatda komplementar nukleotid juftlari soni (bp) bilan o'lchanadi. Masalan, 3-xromosoma insondirasr 160 million bp hajmiga ega bo'lgan DNK molekulasi.3 * 10 6 bp o'lchamli izolyatsiyalangan chiziqli DNK. uzunligi taxminan 1 mm ni tashkil qiladi, shuning uchun 3-inson xromosomasining chiziqli molekulasi uzunligi 5 mm va haploidning barcha 23 xromosomasining DNKsi (~ 3 * 10 9 bp, MR = 1,8 * 10 12) bo'ladi. hujayra - tuxumhujayra yoki sperma - chiziqli shaklda 1 m bo'ladi Jinsiy hujayralar bundan mustasno, inson tanasining barcha hujayralari (taxminan 1013 tasi bor) ikkita xromosoma to'plamini o'z ichiga oladi. Hujayra bo'linishi jarayonida barcha 46 DNK molekulasi replikatsiya qilinadi va 46 xromosomaga qayta tashkil etiladi.
Agar siz inson genomining DNK molekulalarini (22 xromosoma va X va Y yoki X va X xromosomalari) bog'lasangiz, siz taxminan bir metr uzunlikdagi ketma-ketlikni olasiz. Eslatma: Barcha sutemizuvchilar va geterogametik erkak jinsga ega bo'lgan boshqa organizmlar, urg'ochilarda ikkita X xromosoma (XX), erkaklarda bitta X xromosoma va bitta Y xromosoma (XY) mavjud.
Ko'pgina inson hujayralari, shuning uchun bunday hujayralar DNKsining umumiy uzunligi taxminan 2 m ni tashkil qiladi. Voyaga etgan odamda taxminan 10 14 hujayra mavjud, shuning uchun barcha DNK molekulalarining umumiy uzunligi 2 ・ 10 11 km ni tashkil qiladi. Taqqoslash uchun, Yerning aylanasi 4・ 10 4 km, Yerdan Quyoshgacha bo'lgan masofa 1,5 ・ 10 8 km. Hujayralarimizda ixcham o'ralgan DNK mana shunday hayratlanarli!
Eukaryotik hujayralarda DNKni o'z ichiga olgan boshqa organellalar - mitoxondriyalar va xloroplastlar mavjud. Mitoxondriyal va xloroplast DNKsining kelib chiqishi haqida ko'plab farazlar ilgari surilgan. Bugungi kunda umumiy qabul qilingan nuqtai nazar shundan iboratki, ular mezbon hujayralar sitoplazmasiga kirgan va bu organellalarning kashshoflari bo'lgan qadimgi bakteriyalar xromosomalarining rudimentlari. Mitoxondriyal DNK mitoxondriyal tRNK va rRNKni, shuningdek, bir nechta mitoxondriyal oqsillarni kodlaydi. Mitoxondriyal oqsillarning 95% dan ortig'i yadroviy DNK tomonidan kodlangan.
GENLARNING TUZILISHI
Prokaryotlar va eukariotlardagi genning tuzilishini, ularning o'xshash va farqlarini ko'rib chiqing. Gen faqat bitta protein yoki RNKni kodlaydigan DNK bo'lagi bo'lishiga qaramay, u to'g'ridan-to'g'ri kodlash qismidan tashqari, prokaryotlar va eukariotlarda boshqa tuzilishga ega bo'lgan tartibga soluvchi va boshqa tarkibiy elementlarni ham o'z ichiga oladi.
Kodlash ketma-ketligi- genning asosiy tarkibiy va funktsional birligi, unda nukleotidlarning tripletlari kodlanadi.aminokislotalar ketma-ketligi. U boshlang'ich kodoni bilan boshlanadi va to'xtash kodon bilan tugaydi.
Kodlash ketma-ketligidan oldin va keyin tarjima qilinmagan 5'- va 3'-ketliklar... Ular tartibga solish va yordamchi funktsiyalarni bajaradilar, masalan, ribosomaning m-RNKga tushishini ta'minlaydilar.
Tarjima qilinmagan va kodlovchi ketma-ketliklar transkripsiya birligini - transkripsiyalangan DNK bo'limini, ya'ni m-RNK sintezi sodir bo'ladigan DNK bo'limini tashkil qiladi.
Terminator- gen oxiridagi transkripsiyalanmagan DNK mintaqasi, bu erda RNK sintezi to'xtaydi.
Genning boshida tartibga solish sohasi shu jumladan targ'ibotchi va operator.
Promouter- transkripsiya boshlanishida polimeraza bog'lanish ketma-ketligi. Operator maxsus oqsillar bog'lanishi mumkin bo'lgan mintaqa - repressorlar, bu gendan RNK sintezining faolligini kamaytirishi mumkin - boshqacha qilib aytganda, uni kamaytiradi ifoda.
Prokariotlarda gen tuzilishi
Prokaryotlar va eukariotlardagi genlarning umumiy tuzilishi bir-biridan farq qilmaydi - ularning ikkalasi ham promotor va operatorga ega bo'lgan tartibga soluvchi mintaqani, kodlash va tarjima qilinmagan ketma-ketliklarga ega bo'lgan transkripsiya birligini va terminatorni o'z ichiga oladi. Biroq, prokaryotlar va eukariotlarda genlarning tashkil etilishi boshqacha.

Guruch. 18. Prokariotlarda (bakteriyalarda) gen tuzilishi sxemasi -tasvir kattalashtiriladi
Operonning boshida va oxirida bir nechta strukturaviy genlar uchun umumiy tartibga soluvchi hududlar mavjud. Bitta mRNK molekulasi operonning transkripsiyalangan hududidan o'qiladi, u bir nechta kodlash ketma-ketligini o'z ichiga oladi, ularning har biri o'zining boshlang'ich va to'xtash kodoniga ega. Ushbu saytlarning har biridanbitta protein uzilib qoladi. Shunday qilib, bir i-RNK molekulasidan bir nechta oqsil molekulalari sintezlanadi.
Prokaryotlar uchun bir nechta genlarni bitta funktsional birlikka birlashtirish xarakterlidir - operon... Operonning ishi operonning o'zidan sezilarli darajada uzoq bo'lishi mumkin bo'lgan boshqa genlar tomonidan boshqarilishi mumkin - regulyatorlar... Ushbu gendan tarjima qilingan oqsil deyiladi repressor... U operon operatori bilan bog'lanib, undagi barcha genlarning ifodasini bir vaqtning o'zida tartibga soladi.
Bu hodisa prokariotlarga ham xosdir transkripsiya va tarjimani juftlashtirish.
![]()
Guruch. 19 Prokaryotlarda transkripsiya va translatsiyaning konjugatsiya hodisasi - tasvir kattalashtiriladi
Transkripsiya sodir bo'ladigan genetik materialdan tarjima sodir bo'ladigan sitoplazmani ajratib turuvchi yadro qobig'i mavjudligi sababli eukariotlarda bunday konjugatsiya sodir bo'lmaydi. Prokariotlarda DNK shablonida RNK sintezi jarayonida ribosoma sintez qilingan RNK molekulasi bilan darhol bog'lanishi mumkin. Shunday qilib, tarjima transkripsiya tugashidan oldin ham boshlanadi. Bundan tashqari, bir nechta ribosomalar bir vaqtning o'zida bir proteinning bir nechta molekulalarini sintez qilib, bir RNK molekulasiga bog'lanishi mumkin.
Eukariotlarda gen tuzilishi
Eukariotlarning genlari va xromosomalari juda murakkab tuzilgan
Ko'pgina bakteriyalar turlari faqat bitta xromosomaga ega va deyarli barcha hollarda har bir xromosomada har bir genning bitta nusxasi mavjud. Faqat bir nechta genlar, masalan, rRNK genlari bir nechta nusxada mavjud. Genlar va tartibga soluvchi ketma-ketliklar prokariotlarning deyarli butun genomini tashkil qiladi. Bundan tashqari, deyarli har bir gen o'zi kodlaydigan aminokislotalar ketma-ketligiga (yoki RNK ketma-ketligiga) qat'iy mos keladi (14-rasm).
Eukaryotik genlarning strukturaviy va funksional tashkil etilishi ancha murakkab. Eukaryotik xromosomalarni o'rganish va keyinchalik eukaryotik genomlarning to'liq ketma-ketligini aniqlash ko'plab kutilmagan hodisalar keltirdi. Ko'pchilik bo'lmasa ham, eukaryotik genlar qiziqarli xususiyatga ega: ularning nukleotidlar ketma-ketligi polipeptid mahsulotining aminokislotalar ketma-ketligi kodlanmagan bir yoki bir nechta DNK hududlarini o'z ichiga oladi. Bunday tarjima qilinmagan qo'shimchalar genning nukleotidlar ketma-ketligi va kodlangan polipeptidning aminokislotalar ketma-ketligi o'rtasidagi to'g'ridan-to'g'ri yozishmalarni buzadi. Genlarning bu tarjima qilinmagan segmentlari deyiladi intronlar, yoki o'rnatilgan ketma-ketliklar va kodlash segmentlari ekzonlar... Prokariotlarda faqat bir nechta genlarda intronlar mavjud.
Shunday qilib, eukaryotlarda genlarning operonlarga birikmasi deyarli yo'q va eukaryotik genning kodlash ketma-ketligi ko'pincha tarjima qilingan hududlarga bo'linadi. - ekzonlar, va tarjima qilinmagan bo'limlar - intronlar.
Ko'pgina hollarda intronlarning funktsiyasi aniqlanmagan. Umuman olganda, inson DNKsining atigi 1,5% "kodlaydi", ya'ni ular oqsillar yoki RNK haqida ma'lumot olib boradi. Biroq, katta intronlar hisobga olinsa, inson DNKsining 30% genlardan iborat ekanligi ma'lum bo'ladi. Genlar inson genomining nisbatan kichik qismini tashkil qilganligi sababli, DNKning muhim qismi hisobga olinmagan.

Guruch. 16. Eukariotlarda gen tuzilishi sxemasi - tasvir kattalashtiriladi
Har bir gendan avvalo intronlar va ekzonlarni o'z ichiga olgan etuk bo'lmagan yoki pre-RNK sintezlanadi.
Shundan so'ng, splayslash jarayoni sodir bo'ladi, buning natijasida intron hududlari kesiladi va etuk mRNK hosil bo'ladi, undan oqsil sintezlanishi mumkin.

Guruch. 20. Muqobil qo'shish jarayoni - tasvir kattalashtiriladi
Genlarning bunday tashkil etilishi, masalan, bir gendan oqsilning turli shakllari qachon sintezlanishi mumkinligini tushunishga imkon beradi, chunki qo'shilish jarayonida ekzonlarni turli xil ketma-ketlikda tikish mumkin.

Guruch. 21. Prokariotlar va eukariotlar genlari tuzilishidagi farqlar - tasvir kattalashtiriladi
MUTASYONLAR VA MUTAGENEZ
Mutatsiya genotipning doimiy o'zgarishi, ya'ni nukleotidlar ketma-ketligining o'zgarishi deyiladi.
Mutatsiyalarning paydo bo'lishiga olib keladigan jarayon deyiladi mutagenez va organizm, hammasi kimning hujayralari bir xil mutatsiyaga ega - mutant.
Mutatsiya nazariyasi Birinchi marta 1903 yilda Gyugo de Vries tomonidan ishlab chiqilgan. Uning zamonaviy versiyasi quyidagi qoidalarni o'z ichiga oladi:
1. Mutatsiyalar to'satdan, sakrash va chegaralarda paydo bo'ladi.
2. Mutatsiyalar avloddan avlodga o‘tadi.
3. Mutatsiyalar foydali, zararli yoki neytral, dominant yoki retsessiv bo'lishi mumkin.
4. Mutatsiyalarni aniqlash ehtimoli tekshirilgan shaxslar soniga bog'liq.
5. Shunga o'xshash mutatsiyalar qayta-qayta sodir bo'lishi mumkin.
6. Mutatsiyalar maqsadli emas.
Mutatsiyalar turli omillar tufayli yuzaga kelishi mumkin. Ta'sir ostida paydo bo'lgan mutatsiyalarni farqlang mutagen ta'sirlar: jismoniy (masalan, ultrabinafsha yoki radiatsiya), kimyoviy (masalan, kolxitsin yoki reaktiv kislorod turlari) va biologik (masalan, viruslar). Shuningdek, mutatsiyalar sabab bo'lishi mumkin replikatsiya xatolari.
Ko'rinish shartlariga qarab, mutatsiyalar quyidagilarga bo'linadi o'z-o'zidan- ya'ni normal sharoitda paydo bo'lgan mutatsiyalar va qo'zg'atilgan- ya'ni maxsus sharoitlarda paydo bo'lgan mutatsiyalar.
Mutatsiyalar nafaqat yadro DNKsida, balki, masalan, mitoxondriya yoki plastidlar DNKsida ham sodir bo'lishi mumkin. Shunga ko'ra, biz farqlashimiz mumkin yadroviy va sitoplazmatik mutatsiyalar.
Mutatsiyalar natijasida ko'pincha yangi allellar paydo bo'lishi mumkin. Agar mutant allel normalning ta'sirini bostirsa, mutatsiya deyiladi hukmron... Oddiy allel mutantni bostirsa, bunday mutatsiya deyiladi retsessiv... Yangi allellarning paydo bo'lishiga olib keladigan mutatsiyalarning aksariyati retsessivdir.
Ta'siri bo'yicha mutatsiyalar ajralib turadi moslashuvchan tananing atrof-muhitga moslashuvining kuchayishiga olib keladi, neytral omon qolishga ta'sir qilmaydi, zararli organizmlarning atrof-muhit sharoitlariga moslashuvini kamaytiradigan va halokatli rivojlanishning dastlabki bosqichlarida organizmning o'limiga olib keladi.
Natijalarga ko'ra mutatsiyalar ajralib turadi, bu esa olib keladi protein funktsiyasini yo'qotish ga olib keladigan mutatsiyalar paydo bo'lishi oqsil yangi funktsiyaga ega, shuningdek mutatsiyalar genning dozasini o'zgartiring, va shunga ko'ra, undan sintez qilingan oqsilning dozasi.
Mutatsiya tanadagi har qanday hujayrada sodir bo'lishi mumkin. Agar jinsiy hujayrada mutatsiya sodir bo'lsa, u deyiladi germinal(germinal yoki generativ). Bunday mutatsiyalar ular paydo bo'lgan organizmda paydo bo'lmaydi, balki naslda mutantlarning paydo bo'lishiga olib keladi va meros bo'lib qoladi, shuning uchun ular genetika va evolyutsiya uchun muhimdir. Agar mutatsiya boshqa hujayrada sodir bo'lsa, u deyiladi somatik... Bunday mutatsiya, u yoki bu darajada, u paydo bo'lgan organizmda o'zini namoyon qilishi mumkin, masalan, saraton o'smalarining shakllanishiga olib keladi. Biroq, bu mutatsiya meros bo'lib o'tmaydi va naslga ta'sir qilmaydi.
Mutatsiyalar genomning turli o'lchamdagi hududlariga ta'sir qilishi mumkin. Ajratish gen, xromosomali va genomik mutatsiyalar.
Gen mutatsiyalari
Bir gendan kamroq miqyosda yuzaga keladigan mutatsiyalar deyiladi genetik, yoki nuqta (nuqta)... Bunday mutatsiyalar ketma-ketlikda bir yoki bir nechta nukleotidlarning o'zgarishiga olib keladi. Gen mutatsiyalari orasida boralmashtirishlar bir nukleotidni boshqasi bilan almashtirishga olib keladi;o'chirishlar nukleotidlardan birining yo'qolishiga olib keladi,qo'shimchalar ketma-ketlikka qo'shimcha nukleotid qo'shilishiga olib keladi.

Guruch. 23. Gen (nuqta) mutatsiyalari
Proteinga ta'sir qilish mexanizmiga ko'ra, gen mutatsiyalari quyidagilarga bo'linadi:sinonim, bu (genetik kodning degeneratsiyasi natijasida) protein mahsulotining aminokislotalar tarkibining o'zgarishiga olib kelmaydi,noto'g'ri mutatsiyalar, bu bir aminokislotaning boshqasi bilan almashtirilishiga olib keladi va sintez qilingan oqsilning tuzilishiga ta'sir qilishi mumkin, garchi ular ko'pincha ahamiyatsiz bo'lib chiqadi,bema'ni mutatsiyalar kodlash kodonining to'xtash kodon bilan almashtirilishiga olib keladi,ga olib keladigan mutatsiyalar qo'shilish buzilishi:

Guruch. 24. Mutatsiyalar sxemalari
Shuningdek, oqsilga ta'sir qilish mexanizmiga ko'ra, mutatsiyalar ajratiladi, bu esa olib keladi ramka siljishi o'qishlar masalan, qo'shish va o'chirish. Bunday mutatsiyalar, bema'ni mutatsiyalar kabi, garchi ular genning bir nuqtasida sodir bo'lsa-da, ko'pincha oqsilning butun tuzilishiga ta'sir qiladi, bu uning tuzilishini to'liq o'zgartirishga olib kelishi mumkin. xromosoma bo'limi 180 gradusga aylantirilganda, Guruch. 28. Translokatsiya
xromosoma bo'limi 180 gradusga aylantirilganda, Guruch. 28. Translokatsiya
Guruch. 29. Xromosoma duplikatsiyadan oldin va keyin
Genomik mutatsiyalar
Nihoyat, genomik mutatsiyalar butun genomga ta'sir qiladi, ya'ni xromosomalar soni o'zgaradi. Poliploidiyani ajrating - hujayra ploidiyasining ko'payishi va anevloidiya, ya'ni xromosomalar sonining o'zgarishi, masalan, trisomiya (xromosomalardan birida qo'shimcha homologning mavjudligi) va monosomiya (xromosomalarda homologning yo'qligi). xromosoma).
DNK videolar
DNK REPLIKATSIYASI, RNK KODLASH, PROTEINLAR SINTEZI
(Agar video ko'rsatilmasa, u tomonidan mavjud
Kimyoviy tuzilishiga ko'ra DNK ( Deoksiribonuklein kislotasi) bir biopolimer, ularning monomerlari nukleotidlar... Ya'ni, DNK polinukleotid... Bundan tashqari, DNK molekulasi odatda ikki zanjirdan iborat bo'lib, ular bir-biriga nisbatan spiral chiziq bo'ylab o'ralgan (ko'pincha "spiral o'ralgan" deb ataladi) va vodorod aloqalari bilan bog'langan.
Zanjirlarni ham chapga, ham o'ngga (ko'pincha) tomonga burish mumkin.
Ba'zi viruslar bitta DNK zanjiriga ega.
Har bir DNK nukleotidi 1) azotli asos, 2) dezoksiriboza, 3) fosfor kislotasi qoldig'idan iborat.
Ikki o'ng qo'l DNK spiral
DNK tarkibiga quyidagilar kiradi: adenin, guanin, timin va sitozin... Adenin va guanin tegishli purinam, va timin va sitozin - to pirimidin... Ba'zida DNK tarkibida urasil mavjud bo'lib, u odatda RNKga xos bo'lib, u erda timin o'rnini bosadi.
DNK molekulasining bir zanjirining azotli asoslari ikkinchisining azotli asoslari bilan to'ldiruvchilik printsipiga muvofiq bog'langan: adenin faqat timin bilan (bir-biri bilan ikkita vodorod aloqasi hosil qiladi), guanin esa faqat sitozin bilan (uchta bog') .
Nukleotidning o'zida azotli asos siklik shakldagi birinchi uglerod atomiga bog'langan. deoksiriboza bu pentoza (besh uglerod atomiga ega uglevod). Bog' kovalent, glikozid (C-N). Ribozadan farqli o'laroq, dezoksiribozada gidroksil guruhlaridan biri yo'q. Dezoksiriboza halqasi to'rtta uglerod atomi va bitta kislorod atomidan hosil bo'ladi. Beshinchi uglerod atomi halqadan tashqarida joylashgan va fosfor kislotasining qolgan qismi bilan kislorod atomi orqali bog'langan. Shuningdek, uchinchi uglerod atomidagi kislorod atomi orqali qo'shni nukleotidning fosfor kislotasi qoldig'i biriktiriladi.
Shunday qilib, bitta DNK zanjirida qo'shni nukleotidlar bir-biriga bog'langan. kovalent aloqalar deoksiriboza va fosfor kislotasi (fosfodiester bog'i) o'rtasida. Fosfat-dezoksiriboza magistral hosil bo'ladi. Unga perpendikulyar ravishda, boshqa DNK zanjiriga qarab, ikkinchi zanjirning asoslari bilan vodorod bog'lari bilan bog'langan azotli asoslar yo'naltiriladi.
DNKning tuzilishi shundayki, vodorod bilan bog'langan zanjirlarning magistrallari turli yo'nalishlarga yo'naltirilgan (ular "ko'p yo'nalishli", "antiparallel" deb aytadilar). Bir tomoni dezoksiribozaning beshinchi uglerod atomi bilan bog'langan fosfor kislotasi bilan tugaydigan tomonda, ikkinchisi "erkin" uchinchi uglerod atomi bilan tugaydi. Ya'ni, bir zanjirning skeleti ikkinchisiga nisbatan teskari buriladi. Shunday qilib, DNK zanjirlarining tuzilishida 5 ta "uch va 3" uchi ajralib turadi.
DNKni replikatsiya qilishda (ikki marta ko'paytirishda) yangi iplarning sintezi har doim ularning 5-uchidan uchinchisiga o'tadi, chunki yangi nukleotidlar faqat erkin uchinchi uchiga yopishishi mumkin.
Oxir oqibat (bilvosita RNK orqali) DNK zanjiridagi har uchta ketma-ket nukleotid oqsilning bitta aminokislotasini kodlaydi.
DNK molekulasining tuzilishini kashf qilish 1953 yilda F. Krik va D. Uotsonning (boshqa olimlarning dastlabki ishlari ham yordam bergan) ishlari tufayli sodir bo'ldi. Qanday bo'lsa ham Kimyoviy modda DNK 19-asrdayoq ma'lum bo'lgan. XX asrning 40-yillarida genetik ma'lumotlarning tashuvchisi DNK ekanligi ma'lum bo'ldi.
Ikkilamchi spiral DNK molekulasining ikkilamchi tuzilishi hisoblanadi. Eukaryotik hujayralarda DNKning katta miqdori xromosomalarda joylashgan bo'lib, u erda oqsillar va boshqa moddalar bilan bog'lanadi, shuningdek, zichroq o'rashdan o'tadi.
Deoksiribonuklein kislotasi yoki DNK irsiy axborot tashuvchisi hisoblanadi. Hujayralardagi DNKning katta qismi yadroda to'plangan. Bu xromosomalarning asosiy tarkibiy qismidir. Eukariotlarda DNK mitoxondriya va plastidlarda ham uchraydi. DNK bir-biri bilan kovalent bog'langan mononukleotidlardan iborat bo'lib, uzun, tarmoqlanmagan polimerni ifodalaydi. DNK ni tashkil etuvchi mononukleotidlar 4 ta azotli asosdan biri (adenin, guanin, sitozin va timin) va fosfor kislotasi qoldig'i bo'lgan dezoksiribozadan iborat. Ushbu mononukleotidlarning soni juda ko'p. Masalan, bitta xromosomani o'z ichiga olgan prokaryotik hujayralarda DNK molekulyar og'irligi 2 x 10 9 dan ortiq bo'lgan bitta makromolekuladir.
Bir DNK zanjirining mononukleotidlari hosil bo'lishi sababli bir-biri bilan ketma-ket bog'langan kovalent fosfodiester aloqalari Bir mononukleotidning deoksiribozasining OH guruhi va boshqasining fosfor kislotasi qoldig'i o'rtasida. Bitta DNK zanjirining hosil bo'lgan umurtqa pog'onasining bir tomonida azotli asoslar joylashgan. Ularni bir ipga kiyilgan to'rt xil boncuk bilan solishtirish mumkin, chunki ular shakar-fosfat zanjiriga bog'langanga o'xshaydi.
Savol tug'iladi, bu uzun polinukleotid zanjiri hujayra yoki hatto butun organizmni rivojlantirish dasturini qanday kodlashi mumkin? Bu savolga javobni DNKning fazoviy tuzilishi qanday shakllanganligini tushunish orqali olish mumkin. Ushbu molekulaning tuzilishi 1953 yilda J. Uotson va F. Krik tomonidan deşifrlangan va tavsiflangan.
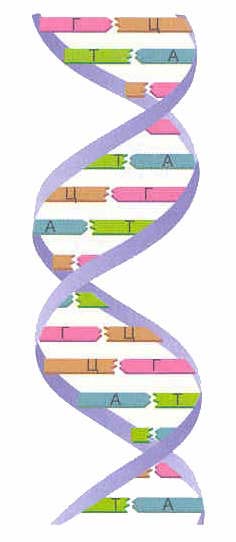
DNK molekulalari bir-biriga parallel bo'lgan va hosil bo'lgan ikkita zanjirdir o'ng qo'l spiral
... Ushbu spiralning kengligi taxminan 2 nm, lekin uning uzunligi yuz minglab nanometrlarga yetishi mumkin. Uotson va Krik DNK modelini taklif qilishdi, unga ko'ra barcha DNK asoslari spiral ichida, tashqarida esa shakar-fosfat magistral bilan joylashgan. Shunday qilib, bir zanjirning asoslari ikkinchisining asoslariga imkon qadar yaqin,
shuning uchun ular orasida vodorod aloqalari hosil bo'ladi. DNK spiralining tuzilishi shundayki, uni tashkil etuvchi polinukleotid zanjirlari faqat ochilgandan keyingina ajratilishi mumkin.
Ikki DNK zanjirining maksimal yaqinligi tufayli uning tarkibida bir xil miqdordagi azotli asoslar (adenin va guanin) va boshqa turdagi azotli asoslar (timin va sitozin) mavjud, ya'ni formula to'g'ri keladi: A + G = T + C... Bu azotli asoslarning kattaligi bilan bog'liq, ya'ni adenin-timin va guanin-sitozin juftlari o'rtasida vodorod bog'ining paydo bo'lishi natijasida hosil bo'lgan tuzilmalarning uzunligi taxminan 1,1 nm. Ushbu juftlarning umumiy o'lchamlari DNK spiralining ichki qismining o'lchamlariga mos keladi. Spiral hosil qilish uchun juft C-T juda kichik bo'ladi va A-G juftligi aksincha, u juda katta. Ya'ni, birinchi DNK zanjirining azotli asosi boshqa DNK zanjirida xuddi shu joyda joylashgan asosni belgilaydi. DNK molekulasida bir-biriga parallel juft zanjirlarda joylashgan nukleotidlarning qat'iy mos kelishi deyiladi. bir-birini to'ldirish (qo'shimchalik). To'g'ri ko'paytirish yoki replikatsiya genetik ma'lumot aynan DNK molekulasining shu xususiyati tufayli mumkin.
DNKda biologik ma'lumotlar shunday qayd qilinadiki, u aniq nusxa ko'chiriladi va avlod hujayralariga uzatilishi mumkin. Unda hujayra bo'linishi sodir bo'lishidan oldin replikatsiya (o'z-o'zidan ikki baravar ko'paytirish ) DNK. Har bir ipda sheriklar zanjiri ketma-ketligini to'ldiruvchi nukleotidlar ketma-ketligi mavjud bo'lganligi sababli, ular aslida bir xil genetik ma'lumotni olib yuradilar. Agar siz iplarni ajratsangiz va ularning har birini ikkinchi ipni qurish uchun shablon (shablon) sifatida ishlatsangiz, ikkita yangi bir xil DNK zanjirini olasiz. Hujayrada DNKning duplikatsiyasi shu tarzda sodir bo'ladi.


