З чого складається ДНК. Нуклеотид – це що таке? Склад, будова, число та послідовність нуклеотидів у ланцюзі ДНК Подвійну спіраль днк стабілізують іони
Нуклеїнові кислоти - високомолекулярні речовини, що складаються з мононуклеотидів, які з'єднані один з одним у полімерний ланцюжок за допомогою 3",5"- фосфодіефірних зв'язків і упаковані в клітинах певним чином.
Нуклеїнові кислоти – біополімери двох різновидів: рибонуклеїнова кислота (РНК) та дезоксирибонуклеїнова кислота (ДНК). Кожен біополімер складається з нуклеотидів, що розрізняються по вуглеводному залишку (рибозе, дезоксирибозе) та одному з азотистих основ (урацил, тимін). Відповідно до цих відмінностей нуклеїнові кислоти і отримали свою назву.
Структура дезоксирибонуклеїнової кислоти
Нуклеїнові кислоти мають первинну, вторинну та третинну структуру.
Первинна структура ДНК
Первинною структурою ДНК називають лінійний полінуклеотидний ланцюг, в якому мононуклеотиди з'єднані 3", 5"-фосфодіефірними зв'язками. Вихідним матеріалом при складанні ланцюга нуклеїнової кислоти в клітині є нуклеозид 5"-трифосфат, який в результаті видалення β і γ залишків фосфорної кислоти здатний приєднати 3" атом вуглецю іншого нуклеозиду. Таким чином, 3"-атом вуглецю однієї дезоксирибози ковалентно зв'язується з 5"-атомом вуглецю іншої дезоксирибози за допомогою одного залишку фосфорної кислоти та утворює лінійний полінуклеотидний ланцюг нуклеїнової кислоти. Звідси і назва: 3", 5"-фосфодіефірні зв'язки. Азотисті основи не беруть участі у поєднанні нуклеотидів одного ланцюга (рис. 1.).
Така сполука між залишком молекули фосфорної кислоти одного нуклеотиду і вуглеводом іншого призводить до утворення пентозо-фосфатного скелета молекули полінуклеотиду, на якому збоку один за одним приєднуються азотисті основи. Їх послідовність розташування ланцюгах молекул нуклеїнових кислот суворо специфічна для клітин різних організмів, тобто. носить видовий характер (правило Чаргафа).
Лінійний ланцюг ДНК, довжина якої залежить від числа нуклеотидів, що входять у ланцюг, має два кінці: один називається 3"-кінцем і містить вільний гідроксил, а інший - 5"-кінцем, містить залишок фосфорної кислоти. Ланцюг полярний і може мати напрям 5"->3" і 3"->5". Винятком є кільцеві ДНК.
Генетичний "текст" ДНК складено за допомогою кодових "слів" - триплетів нуклеотидів, які називаються кодонами. Ділянки ДНК, що містять інформацію про первинну структуру всіх типів РНК, називають структурними генами.
Полінуклеодитні ланцюжки ДНК досягають гігантських розмірів, тому в клітині вони упаковані певним чином.
Вивчаючи склад ДНК, Чаргафф (1949) встановив важливі закономірності щодо змісту окремих підстав ДНК. Вони допомогли розкрити вторинну структуру ДНК. Ці закономірності називають правилами Чаргафа. Правила Чаргафа
Ці правила свідчать, що з побудові ДНК має дотримуватися досить суворе відповідність (спарювання) не пуринових і пиримидиновых підстав взагалі, саме тиміну з аденіном і цитозину з гуаніном. На підставі цих правил у тому числі, в 1953 Уотсон і Крик запропонували модель вторинної структури ДНК, що отримала назву подвійної спіралі (рис.). |

Вторинна структура ДНК
Вторинна структура ДНК - це подвійна спіраль, модель якої була запропонована Д.Уотсоном та Ф.Кріком у 1953 році.
Передумови створення моделі ДНК
В результаті початкових аналізів склалося уявлення, що ДНК будь-якого походження містить усі чотири нуклеотиди в рівних молярних кількостях. Однак у 1940-х роках Е. Чаргафф та його співробітники в результаті аналізу ДНК, виділених з різноманітних організмів, ясно показали, що азотисті основи містяться в них у різних кількісних співвідношеннях. Чаргафф виявив, що, хоча ці співвідношення однакові для ДНК із усіх клітин одного й того ж виду організмів, ДНК від різних видів можуть помітно відрізнятися за змістом тих чи інших нуклеотидів. Це наводило на думку, що відмінності у співвідношенні азотистих основ, можливо, пов'язані з якимсь біологічним кодом. Хоча співвідношення окремих пуринових і піримідинових основ у різних зразках ДНК виявилося неоднаковим, при порівнянні результатів аналізів виявилася певна закономірність: у всіх зразках загальна кількість пуринів дорівнювала загальній кількості піримідинів (А + Г = Т + Ц), кількість аденіну - кількості тиміну (А = Т), а кількість гуаніну – кількості цитозину (Г = Ц). ДНК, виділена з клітин ссавців, була загалом багатша аденіном і тиміном і щодо бідніше гуаніном і цитозином, тоді як у бактерій ДНК була багатша гуаніном і цитозином і щодо бідніше аденіном і тиміном. Ці дані склали важливу частину фактичного матеріалу, на основі якого пізніше було побудовано модель структури ДНК Уотсона – Крику.
Ще однією важливою непрямою вказівкою на можливу структуру ДНК стали дані Л. Полінга про будову білкових молекул. Полінг показав, що можливо кілька різних стійких конфігурацій амінокислотного ланцюга в молекулі білка. Одна з поширених конфігурацій пептидного ланцюга - α-спіраль - є правильною гвинтоподібною структурою. За такої структури можливе утворення водневих зв'язків між амінокислотами, що знаходяться на суміжних витках ланцюга. Полінг описав α-спіральну конфігурацію поліпептидного ланцюга в 1950 році і висловив припущення, що молекули ДНК, ймовірно, мають спіральну структуру, закріплену водневими зв'язками.
Однак найцінніші відомості про будову молекули ДНК дали результати рентгеноструктурного аналізу. Рентгенівські промені, проходячи крізь кристал ДНК, зазнають дифракції, тобто відхиляються у певних напрямках. Ступінь та характер відхилення променів залежать від структури самих молекул. Дифракційна рентгенограма (рис. 3) дає досвідченому оку ряд непрямих вказівок щодо будови молекул досліджуваної речовини. Аналіз дифракційних рентгенограм ДНК привів до висновку, що азотисті основи (що мають плоску форму) укладені на зразок стопки тарілок. Рентгенограми дозволили виявити у структурі кристалічної ДНК три основних періоди: 0,34, 2 і 3,4 нм.

Модель ДНК Уотсона-Кріка
Виходячи з аналітичних даних Чаргаффа, рентгенограм, отриманих Уілкінсом та досліджень хіміків, що надали відомості про точні відстані між атомами в молекулі, про кути між зв'язками даного атома та про величину атомів, Уотсон і Крик почали будувати фізичні моделі окремих складових частин молекули ДНК у певному масштабі. і "підганяти" їх один до одного з таким розрахунком, щоб отримана система відповідала різним експериментальним даним [показати] .
Ще раніше було відомо, що в ланцюзі ДНК сусідні нуклеотиди з'єднані фосфодіефірними містками, що зв'язують 5"-вуглецевий дезоксирибози атом одного нуклеотиду з 3"-вуглецевим атомом дезоксирибози наступного нуклеотиду. Уотсон і Крик не сумнівалися, що період 0,34 нм відповідає відстані між послідовними нуклеотидами в ланцюзі ДНК. Далі можна було припускати, що період 2 нм відповідає товщині ланцюга. А для того, щоб пояснити, якій реальній структурі відповідає період 3,4 нм, Уотсон і Крик, як і раніше Полінг, припустили, що ланцюг закручений у вигляді спіралі (або, точніше, утворює гвинтову лінію, тому що спіраль у строгому сенсі цього слова виходить тоді, коли витки утворюють у просторі конічну, а чи не циліндричну поверхню). Тоді період 3,4 нм відповідатиме відстані між послідовними витками цієї спіралі. Така спіраль може бути дуже щільною або дещо розтягнутою, тобто витки її можуть бути пологими або крутими. Оскільки період 3,4 нм рівно в 10 разів більший за відстань між послідовними нуклеотидами (0,34 нм), ясно, що кожен повний виток спіралі містить 10 нуклеотидів. За цими даними Уотсон і Крик змогли обчислити щільність полінуклеотидного ланцюга, закрученого в спіраль діаметром 2 нм, з відстанню між витками, що дорівнює 3,4 нм. Виявилося, що у такого ланцюга щільність була б удвічі меншою за фактичну щільність ДНК, яка була вже відома. Довелося припустити, що молекула ДНК і двох ланцюгів - що це подвійна спіраль з нуклеотидів.
Наступним завданням було, звісно, з'ясування просторових відносин між обома ланцюгами, що утворюють подвійну спіраль. Випробувавши на своїй фізичній моделі ряд варіантів розташування ланцюгів, Вотсон і Крик знайшли, що всім наявним даним найкраще відповідає такий варіант, у якому дві полінуклеотидні спіралі йдуть у протилежних напрямках; при цьому ланцюги, що складаються із залишків цукру та фосфату, утворюють поверхню подвійної спіралі, а пурини та піримідини розташовуються всередині. Розташовані один проти одного підстави, що належать двом ланцюгам, попарно з'єднані водневими зв'язками; саме ці водневі зв'язки та утримують ланцюги разом, фіксуючи таким чином загальну конфігурацію молекули.
Подвійну спіраль ДНК можна уявити у вигляді гвинтоподібно закручених мотузкових сходів, так щоб перекладини її залишалися в горизонтальному положенні. Тоді дві поздовжні мотузки відповідатимуть ланцюгам із залишків цукру та фосфату, а перекладини – парам азотистих основ, з'єднаних водневими зв'язками.
В результаті подальшого вивчення можливих моделей Вотсон і Крик дійшли висновку, що кожна "перекладина" повинна складатися з одного пурину та одного піримідину; при періоді 2 нм (що відповідає діаметру подвійної спіралі) для двох пуринів не вистачило б місця, а два піримідини не могли б при цьому розташовуватись досить близько один до одного, щоб утворити належні водневі зв'язки. Поглиблене дослідження детальної моделі показало, що аденін і цитозин, складаючи відповідну за розмірами комбінацію, все ж таки не могли б розташовуватися таким чином, щоб між ними утворилися водневі зв'язки. Аналогічні повідомлення змусили виключити також комбінацію гуанін – тимін, тоді як поєднання аденін – тимін та гуанін – цитозин виявилися цілком прийнятними. Природа водневих зв'язків така, що аденін утворює пару з тиміном, а гуанін – з цитозином. Це уявлення про специфічне спаровування підстав дозволяло пояснити "правило Чаргаффа", згідно з яким у будь-якій молекулі ДНК кількість аденіну завжди дорівнює вмісту тиміну, а кількість гуаніну - кількості цитозину. Між аденіном і тиміном утворюються два водневі зв'язки, а між гуаніном та цитозином – три. Завдяки цій специфічності в утворенні водневих зв'язків проти кожного аденіну в одному ланцюгу в інший виявляється тимін; так само проти кожного гуаніну може бути тільки цитозин. Таким чином, ланцюги комплементарні один одному, т. Е. Послідовність нуклеотидів в одному ланцюгу однозначно визначає їх послідовність в іншій. Два ланцюги йдуть у протилежних напрямках, та його кінцеві фосфатні групи перебувають у протилежних кінцях подвійний спіралі.
В результаті своїх досліджень, в 1953 Уотсон і Крик запропонували модель будови молекули ДНК (рис. 3), яка залишається актуальною по теперішній час. Згідно моделі молекула ДНК складається з двох комплементарних полінуклеотидних ланцюгів. Кожен ланцюг ДНК є полінуклеотидом, що складається з декількох десятків тисяч нуклеотидів. У ній сусідні нуклеотиди утворюють регулярний пентозо-фосфатний кістяк за рахунок з'єднання залишку фосфорної кислоти та дезоксирибози міцним ковалентним зв'язком. Азотисті основи одного полінуклеотидного ланцюга при цьому розташовуються у строго визначеному порядку проти азотистих основ іншої. Чергування азотистих основ у полінуклеотидному ланцюзі нерегулярне.
Розташування азотистих підстав у ланцюзі ДНК є комплементарним (від грец. "комплемент" - доповнення), тобто. проти аденіну (А) завжди виявляється тимін (Т), а проти гуаніну (Г) – лише цитозин (Ц). Це тим, що і Т, і навіть Г і Ц суворо відповідають одне одному, тобто. доповнюють одне одному. Така відповідність визначається хімічною структурою основ, що дозволяє утворити водневі зв'язки в парі пурину і піримідину. Між А і Т виникають два зв'язки, між Г і Ц – три. Ці зв'язки забезпечують часткову стабілізацію молекули ДНК у просторі. Стійкість подвійної спіралі у своїй прямо пропорційна числу зв'язків G≡С, є стабільними проти зв'язками А=Т.
Відома послідовність розташування нуклеотидів одного ланцюга ДНК дозволяє за принципом комплементарності встановити нуклеотиди іншого ланцюга.
Крім того, встановлено, що азотисті основи, що мають ароматичну структуру, у водному розчині розташовуються один над одним, формуючи стопку монет. Такий процес формування стопок з органічних молекулназивається стекінг. Полінуклеотидні ланцюги молекули ДНК аналізованої моделі Уотсона-Кріка мають аналогічний фізико-хімічний стан, їх азотисті основи розташовуються у вигляді стопки монет, між площинами яких виникають ван-дер-ваальсові взаємодії (стекінг-взаємодія).
Водневі зв'язки між комплементарними основами (по горизонталі) та стекінг-взаємодія між площинами основ у полінуклеотидному ланцюгу за рахунок ван-дер-ваальсових сил (по вертикалі) забезпечує молекулі ДНК додаткову стабілізацію у просторі.
Сахарофосфатні кістяки обох ланцюгів звернені назовні, а підстави всередину, назустріч один одному. Напрямок ланцюгів в ДНК антипаралельно (одна з них має напрямок 5"->3", інша - 3"->5", тобто 3"-кінець одного ланцюга розташований навпроти 5"-кінця іншого.). Ланцюги утворюють праві спіралі із загальною віссю. Один виток спіралі становить 10 нуклеотидів, розмір витка 34 нм, висота кожного нуклеотиду 034 нм діаметр спіралі - 20 нм. В результаті обертання одного ланцюга навколо іншого утворюється велика борозна (діаметром близько 20 Å) і мала борозна (близько 12 Å) подвійної спіралі ДНК. Така форма подвійної спіралі Уотсона-Кріка надалі отримала назву В-форми. У клітинах ДНК зазвичай існує у формі, яка є найстабільнішою.
Функції ДНК
Запропонована модель пояснювала багато біологічних властивостей дезоксирибонуклеїнової кислоти, в тому числі зберігання генетичної інформації та різноманіття генів, що забезпечується великою різноманітністю послідовних поєднань 4-х нуклеотидів і фактом існування генетичного коду, здатність до самовідтворення та передачі генетичної інформації. у вигляді білків, а також будь-яких інших сполук, що утворюються за допомогою білків-ферментів.
Основні функції ДНК.
- ДНК є носієм генетичної інформації, що забезпечується фактом існування генетичного коду.
- Відтворення та передана генетичної інформації у поколіннях клітин та організмів. Ця функція забезпечується процесом реплікації.
- Реалізація генетичної інформації як білків, і навіть будь-яких інших сполук, що утворюються з допомогою білків-ферментів. Ця функція забезпечується процесами транскрипції та трансляції.

Форми організації дволанцюжкової ДНК
ДНК може утворювати кілька типів подвійних спіралей (рис.4). В даний час вже відомо шість форм (від А до Е та Z-форма).
Структурні форми ДНК, як встановила Розалінда Франклін, залежить від насичення водою молекули нуклеїнової кислоти. У дослідженнях волокон ДНК за допомогою рентгеноструктурного аналізу було показано, що рентгенограма радикальним чином залежить від того, за якої відносної вологості, за якого ступеня насичення водою цього волокна відбувається експеримент. Якщо волокно було досить насичене водою, виходила одна рентгенограма. При висушуванні виникала зовсім інша рентгенограма, що відрізняється від рентгенограми волокна високої вологості.
Молекула ДНК високої вологості отримала назву В-форми. За фізіологічних умов (низька концентрація солі, високий рівень гідратації) домінуючим структурним типом ДНК є В-форма (основна форма дволанцюжкової ДНК - модель Уотсона-Крика). Крок спіралі такої молекули дорівнює 34 нм. На виток припадає 10 комплементарних пар у вигляді скручених стосів "монет" - азотистих основ. Стопки утримуються водневими зв'язками між двома протилежними "монетами" стопок, і "обмотані" двома стрічками фосфодіефірного остова, закрученими у праву спіраль. Площини азотистих основ перпендикулярні до осі спіралі. Сусідні комплементарні пари повернені одна щодо одної на 36°. Діаметр спіралі 20Å, причому пуриновий нуклеотид займає 12Å, а піримідиновий – 8Å.
Молекула ДНК нижчої вологості отримала назву А-форми. А-форма утворюється в умовах менш високої гідратації і за більш високого вмісту іонів Na + або К + . Ця ширша правоспіральна конформація має 11 пар азотистих основ на виток. Площини азотистих основ мають сильніший нахил до осі спіралі, вони відхилені від нормалі до осі спіралі на 20 °. Звідси випливає наявність внутрішньої порожнечі діаметром 5Å. Відстань між сусідніми нуклеотидами становить 0,23 нм, довжина витка – 2,5 нм, діаметр спіралі – 2,3 нм.
Спочатку вважали, що А-форма ДНК менш важлива. Однак надалі з'ясувалося, що А-форма ДНК, як і В-форма, має величезне біологічне значення. А-форму має спіраль РНК-ДНК у комплексі матриця-затравка, а також спіраль РНК-РНК та шпилькові структури РНК (2'-гідроксильна група рибози не дозволяє молекулам РНК утворювати В-форму). А-форма ДНК виявлена у суперечках. Встановлено, що А-форма ДНК у 10 разів стійкіша до дії УФ-променів, ніж В-форма.
А-форму та В-форму називають канонічними формамиДНК.
Форми С-Етакож правоспіральні, їх освіту можна спостерігати лише у спеціальних експериментах, і, мабуть, вони не існують in vivo. С-форма ДНК має структуру, подібну до В-ДНК. Число пар основ на виток становить 9,33, довжина витка спіралі дорівнює 3,1 нм. Пари основ нахилені на кут 8 градусів щодо перпендикулярного положення до осі. Жолобки за розмірами близькі до жолобків В-ДНК. При цьому головний жолобок дещо дрібніший, а мінорний жолобок – глибший. У С-форму можуть переходити природні та синтетичні полінуклеотиди ДНК.
| Таблиця 1. Характеристика деяких типів структур ДНК | |||
| Тип спіралі | A | B | Z |
| Крок спіралі | 0,32 нм | 3,38 нм | 4,46 нм |
| Закрученість спіралі | Права | Права | Ліва |
| Число пар основ на виток | 11 | 10 | 12 |
| Відстань між площинами основ | 0,256 нм | 0,338 нм | 0,371 нм |
| Конформація глікозидного зв'язку | анти | анти | анти-С син-Г |
| Конформація фуранозного циклу | С3"-ендо | С2"-ендо | С3"-ендо-Г С2"-ендо-Ц |
| Ширина борозенки, малий/великий | 1,11/0,22 нм | 0,57/1,17 нм | 0,2/0,88 нм |
| Глибина борозенки, малий/великий | 0,26/1,30 нм | 0,82/0,85 нм | 1,38/0,37 нм |
| Діаметр спіралі | 2,3 нм | 2,0 нм | 1,8 нм |
Структурні елементи ДНК
(Неканонічні структури ДНК)
До структурних елементів ДНК можна віднести незвичайні структури, обмежені спеціальними послідовностями:
|

Z-форма ДНКбула відкрита 1979 року щодо гексануклеотида d(CG)3 - . Її відкрив професор Массачусетського технологічного інституту Олександр Річ із співробітниками. Z-форма стала одним з найважливіших структурних елементів ДНК у зв'язку з тим, що її утворення спостерігалося в ділянках ДНК, де пурини чергуються з піримідинами (наприклад, 5'-ГЦГЦГЦ-3'), або в повторах 5'-ЦГЦГЦГ-3' , що містять метильований цитозин Істотною умовою утворення та стабілізації Z-ДНК була присутність у ній пуринових нуклеотидів у син-конформації, що чергуються з піримідиновими основами в анти-конформації.
Природні молекули ДНК в основному існують у правій формі, якщо вони не містять послідовностей типу (ЦГ)n. Однак, якщо такі послідовності входять до складу ДНК, ці ділянки при зміні іонної сили розчину або катіонів, що нейтралізують негативний заряд на фосфодіефірному каркасі, можуть переходити в Z-форму, при цьому інші ділянки ДНК в ланцюзі залишаються в класичній В-формі. Можливість такого переходу вказує на те, що два ланцюги у подвійній спіралі ДНК знаходяться в динамічному стані і можуть розкручуватися один щодо одного, переходячи з правої форми в ліву та навпаки. Біологічні наслідки такої лабільності, що допускає конформаційні перетворення структури ДНК, поки що не цілком зрозумілі. Вважають, що ділянки Z-ДНК відіграють певну роль у регуляції експресії деяких генів та беруть участь у генетичній рекомбінації.
Z-форма ДНК - це лівозакручена подвійна спіраль, в якій фосфодіефірний кістяк розташований зигзагоподібно вздовж осі молекули. Звідси назва молекули (zigzag)-ДHK. Z-ДНК - найменш скручена (12 пар основ на виток) і найтонша з відомих у природі. Відстань між сусідніми нуклеотидами становить 0,38 нм, довжина витка – 4,56 нм, діаметр Z-ДНК – 1,8 нм. Крім того, зовнішній виглядцієї молекули ДНК відрізняється наявністю однієї борозенки.
Z-форма ДНК була виявлена в клітинах прокаріотів та еукаріотів. В даний час отримані антитіла, здатні відрізняти Z-форму від В-форми ДНК. Ці антитіла пов'язуються з певними ділянками гігантських хромосом клітин слинних залоз дрозофіли (Dr. melanogaster). За реакцією зв'язування легко стежити через незвичайну будову цих хромосом, у яких щільніші ділянки (диски) контрастують з менш щільними (міждисками). Ділянки Z-ДНК розташовані у міждисках. З цього випливає, що Z-форма реально існує у природних умовах, хоча розміри індивідуальних ділянок Z-форми поки що невідомі.

(перевертачі) - найбільш відомі і часто зустрічаються в ДНК послідовності основ. Паліндромом називають слово або фразу, яке читається зліва направо та навпаки однаково. Прикладами таких слів або фраз є: Шалаш, Казак, Потоп, А троянда впала на лапу Азора. У застосуванні до ділянок ДНК даний термін (паліндром) означає однакове чергування нуклеотидів уздовж ланцюга праворуч наліво та зліва направо (подібно до літер у слові "курінь" тощо).
Паліндром характеризується наявністю інвертованих повторів послідовностей підстав, що мають симетрію другого порядку щодо двох ланцюгів ДНК. Такі послідовності, з цілком зрозумілої причини, є самокомплементарними і мають схильність до утворення шпилькових чи хрестоподібних структур (мал.). Шпильки допомагають регуляторним білкам дізнаватися про місце списування генетичного тексту ДНК хромосом.
У тих випадках, коли інвертований повтор присутній в одному і тому ж ланцюгу ДНК така послідовність називається дзеркальним повтором. Дзеркальні повтори не володіють властивостями самокомплементарності і тому не здатні до формування шпилькових або хрестоподібних структур. Послідовності такого типу виявлені практично у всіх великих молекулах ДНК і можуть включати від декількох пар основ до декількох тисяч пар основ.
Присутність паліндромів у вигляді хрестоподібних структур в еукаріотичних клітинах не доведено, хоча деяка кількість хрестоподібних структур виявлена в умовах in vivo у клітинах E. coli. Наявність у складі РНК або одноланцюгових ДНК самокомплементарних послідовностей служить основною причиною згортання в розчинах нуклеїнового ланцюга певну просторову структуру, що відрізняється формуванням безлічі "шпильок".

Н-форма ДНК- це спіраль, яку утворюють три ланцюги ДНК - потрійна спіраль ДНК. Є комплексом уотсон-криківської подвійної спіралі з третьою одноланцюжковою ниткою ДНК, яка вкладається в її великий жолобок, з утворенням так званої хугстинівської пари.
Утворення подібного триплексу відбувається внаслідок складання подвійної спіралі ДНК таким чином, що половина її ділянки залишається у вигляді подвійної спіралі, а друга половина роз'єднується. При цьому одна з роз'єднаних спіралей утворює нову структуру з першою половиною подвійної спіралі - потрійну спіраль, а друга виявляється неструктурованою у вигляді однониткової ділянки. Особливістю цього структурного переходу є різка залежність від рН середовища, протони якого стабілізують нову структуру. В силу цієї особливості нова структура отримала назву Н-форми ДНК, утворення якої було виявлено у надспіралізованих плазмідах, що містять гомопурин-гомопіримідинові ділянки, що є дзеркальним повтором.
У подальших дослідженнях була встановлена можливість здійснення структурного переходу деяких гомопурин-гомопіримідинових двониткових полінуклеотидів з утворенням тринітієвої структури, що містить:
- одну гомопуринову та дві гомопіримідинові нитки ( Py-Pu-Py триплекс) [Хугстинівська взаємодія].
Складові блоки Py-Pu-Py триплексу - канонічні ізоморфні CGC+ та TAT тріади. Стабілізація триплексу потребує протонування тріади CGC+, тому ці триплекси залежать від рН розчину.
- одну гомопіримідинову та дві гомопуринові нитки ( Py-Pu-Pu триплекс) [Зворотне хугстинівська взаємодія].
Складові блоки Py-Pu-Pu триплексу - канонічні ізоморфні CGG та TAA тріад. Істотною властивістю Py-Pu-Pu триплексів є залежність їхньої стабільності від присутності двозарядних іонів, причому для стабілізації триплексів різної послідовності необхідні різні іони. Оскільки для утворення Py-Pu-Pu триплексів не потрібно протонування нуклеотидів, що входять до їх складу, такі триплекси можуть існувати при нейтральних pH.
Прим.: пряма і зворотна хугстинівська взаємодія пояснюється симетрією 1-метилтиміну: поворот на 180° призводить до того, що місце атома О4 займає атом О2, система водневих зв'язків зберігається.
Відомі два види потрійних спіралей:
- паралельні потрійні спіралі, в яких полярність третього ланцюга збігається з полярністю гомопуринового ланцюга Уотсон-Кріковського дуплекса
- антипаралельні потрійні спіралі, у яких полярності третього та гомопуринового ланцюгів протилежні.

G-квадруплекс- 4-х спіральна ДНК. Така структура утворюється у випадку, якщо є чотири гуаніни, які утворюють так званий G-квадруплекс - хоровод із чотирьох гуанінів.
Перші натяки на можливість утворення таких структур були отримані задовго до проривної роботи Уотсона та Крику – ще 1910 року. Тоді німецький хімік Івар Банг виявив, що один із компонентів ДНК – гуанозинова кислота – при високих концентраціях утворює гелі, у той час як інші складові частини ДНК такою властивістю не мають.
У 1962 році за допомогою рентгеноструктурного методу вдалося встановити структуру осередку цього гелю. Вона виявилася складена з чотирьох залишків гуаніну, що зв'язують один одного по колу і утворюють характерний квадрат. У центрі зв'язок підтримує іон металу (Na, K, Mg). Такі самі структури можуть утворюватися й у ДНК, якщо у ній багато гуаніну. Ці плоскі квадрати (G-квартети) складаються в стоси, і виходять досить стійкі, щільні структури (G-квадруплекси).
У чотириспіральні комплекси можуть сплітатися чотири окремі ланцюжки ДНК, але це є винятком. Найчастіше єдина нитка нуклеїнової кислоти просто зав'язується у вузол, утворюючи характерні потовщення (наприклад, на кінцях хромосом), або дволанцюжкова ДНК на якійсь багатій гуаніній ділянці утворює локальний квадруплекс.
Найбільш вивчено існування квадруплексів на кінцях хромосом – на теломерах та в онкопромоторах. Однак досі повне уявлення про локалізацію такої ДНК у людських хромосомах не відоме.
Всі ці незвичайні структури ДНК у лінійній формі нестабільні порівняно з В-формою ДНК. Однак ДНК часто існує в кільцевій формі топологічної напруги, коли вона має так звану надспіралізацію. У цих умовах легко утворюються неканонічні структури ДНК: Z-форми, "хрести" та "шпильки", H-форми, гуанінові квадрулекси та i-мотив.
- Суперспіралізована форма - відзначається при виділенні з ядра клітини без пошкодження пентозо-фосфатного кістяка. Має форму надскручених замкнених кілець. У надскрученому стані подвійна спіраль ДНК хоча б один раз "перекручена сама на себе", тобто містить хоча б один супервиток (набуває форми вісімки).
- Релаксований стан ДНК – спостерігається при одиночному розриві (розриві однієї нитки). При цьому супервитки зникають і ДНК набуває форми замкненого кільця.
- Лінійна форма ДНК – спостерігається при розриві двох ниток подвійної спіралі.
Третинна структура ДНК
Третинна структура ДНКутворюється в результаті додаткового скручування у просторі двоспіральної молекули – її суперспіралізації. Суперспіралізація молекули ДНК в еукаріотичних клітинах на відміну від прокаріотів здійснюється у формі комплексів з білками.
ДНК еукаріотів майже вся знаходиться в хромосомах ядер, лише невелика кількість її міститься в мітохондріях, а у рослин і в пластидах. Основна речовина хромосом еукаріотичних клітин (у тому числі і хромосом людини) - це хроматин, що складається з дволанцюгової ДНК, гістонових та негістонових білків.
Гістонові білки хроматину
Гістони – прості білки, складають до 50% хроматину. У всіх вивчених клітинах тварин і рослин виявлено п'ять основних класів гістонів: H1, H2A, H2B, H3, H4, що відрізняються за розмірами, амінокислотним складом і величиною заряду (завжди позитивний).
Гістон Н1 ссавців складається з одного поліпептидного ланцюга, що містить приблизно 215 амінокислот; розміри інших гістонів варіюють від 100 до 135 амінокислот. Всі вони спіралізовані та скручені в глобулу діаметром близько 2,5 нм, містять надзвичайно велику кількість позитивно заряджених амінокислот лізину та аргініну. Гістони можуть бути ацетильовані, метильовані, фосфорильовані, полі(АДФ)-рибозильовані, а гістони Н2А та Н2В – ковалентно пов'язані з убіквітіном. Яка роль таких модифікацій у становленні структури та виконання функцій гістонами до кінця поки що не з'ясовано. Передбачається, що в цьому полягає їх здатність взаємодіяти з ДНК та забезпечувати один із механізмів регуляції дії генів.
Гістони взаємодіють з ДНК в основному через іонні зв'язки (сольові містки), що утворюються між негативно зарядженими фосфатними групами ДНК та позитивно зарядженими лізиновими та аргініновими залишками гістонів.
Негістонові білки хроматину
Негістонові білки, на відміну від гістонів, дуже різноманітні. Виділено до 590 різних фракцій ДНК-зв'язуючих негістонових білків. Їх ще називають кислими білками, тому що в їх структурі переважають кислі амінокислоти (вони є поліаніонами). З різноманітністю негістонових білків пов'язують специфічну регуляцію активності хроматину. Наприклад, ферменти, необхідні для реплікації та експресії ДНК, можуть зв'язуватися з хроматином тимчасово. Інші білки, скажімо, що у різних процесах регуляції, пов'язуються з ДНК лише у специфічних тканинах чи певних стадіях диференціації. Кожен білок комплементарний певній послідовності ДНК нуклеотидів (сайт ДНК). До цієї групи відносять:
- сімейство сайт-специфічних білків типу "цинкові пальці". Кожен "цинковий палець" дізнається певний сайт, що складається з 5 нуклеотидних пар.
- сімейство сайт-специфічних білків - гомодімери. Фрагмент такого білка, який контактує з ДНК, має структуру "спіраль-поворот-спіраль".
- білки високої рухливості (HMG-білки – від англ, high mobility gel proteins) – група структурних та регуляторних білків, які постійно асоційовані з хроматином. Вони мають молекулярну масу менше 30 кД та характеризуються високим вмістом заряджених амінокислот. Завдяки невеликій молекулярній масі HMG-білки мають високу рухливість при електрофорезі в поліакриламідному гелі.
- ферменти реплікації, транскрипції та репарації.
За участю структурних, регуляторних білків та ферментів, що беруть участь у синтезі ДНК та РНК, нитка нуклеосом перетворюється на висококонденсований комплекс білків та ДНК. Утворена структура у 10 000 разів коротша за вихідну молекулу ДНК.

Хроматин
Хроматин - це комплекс білків з ядерною ДНК та неорганічними речовинами. Основна частина хроматину є неактивною. Вона містить щільно упаковану, конденсовану ДНК. Це гетерохроматин. Розрізняють конститутивний, генетично неактивний хроматин (сателітна ДНК), що складається з неекспресованих областей, і факультативний - неактивний у ряді поколінь, але за певних обставин здатний еспресувати.
Активний хроматин (еухроматину) неконденсований, тобто. упакований менш щільно. У різних клітинах його вміст становить від 2 до 11%. У клітинах головного мозку його найбільше – 10-11%, у клітинах печінки – 3-4 та нирок – 2-3%. Зазначається активна транскрипція еухроматину. При цьому його структурна організація дозволяє використовувати ту саму генетичну інформацію ДНК, властиву даному виду організму, по-різному в спеціалізованих клітинах.
В електронному мікроскопі зображення хроматину нагадує намисто: кулясті потовщення розміром близько 10 нм, розділені ниткоподібними перемичками. Ці кулясті потовщення названі нуклеосомами. Нуклеосома є структурною одиницею хроматину. Кожна нуклеосома містить надспіральний сегмент ДНК довжиною 146 пар нуклеотидів, намотаний з утворенням 1,75 лівих витків на нуклеосомний кор. Нуклеосомний кор - це гістоновий октамер, що складається з гістонів Н2А, Н2В, Н3 і Н4 по дві молекули кожного виду (рис. 9), який виглядає як диск діаметром 11 нм і товщиною 5,7 нм. П'ятий гістон, Н1, не входить до складу нуклеосомного кора і не бере участь у процесі намотування ДНК на гістоновий октамер. Він контактує з ДНК у тих місцях, де подвійна спіраль входить та виходить із нуклеосомного кора. Це міжкорові (лінкерні) ділянки ДНК, довжина яких залежить від типу клітин від 40 до 50 нуклеотидних пар. Внаслідок цього варіює і довжина фрагмента ДНК, що входить до складу нуклеосом (від 186 до 196 нуклеотидних пар).
До складу нуклеосом входить приблизно 90% ДНК, решта її припадає на лінкер. Вважається, що нуклеосоми - це фрагменти хроматину, що "мовчить", а лінкер - активного. Однак нуклеосоми можуть розгортатися та переходити в лінійну форму. Розгорнуті нуклеосоми є активним хроматином. Так наочно проявляється залежність функції структури. Можна вважати, що чим більше хроматину знаходиться у складі глобулярних нуклеосів, тим менш він активний. Вочевидь, у різних клітинах неоднакова частка хроматину, що лежить, пов'язана з кількістю таких нуклеосом.
На електронно-мікроскопічних фотографіях залежно від умов виділення та ступеня розтягування хроматин може виглядати не тільки як довга нитка з потовщеннями – "намистинками" нуклеосом, але і як більш коротка і щільніша фібрила (волокно) діаметром 30 нм, утворення якої спостерігається при взаємодії гістону Н1, пов'язаного з лінкерною ділянкою ДНК та гістону Н3, що призводить до додаткового скручування спіралі з шести нуклеосом на виток з утворенням соленоїда діаметром 30 нм. При цьому гістоновий білок може перешкоджати транскрипції низки генів і таким чином регулювати їхню активність.
В результаті описаних вище взаємодій ДНК з гістонами сегмент подвійної спіралі ДНК з 186 пар основ із середнім діаметром 2 нм і довжиною 57 нм перетворюється на спіраль діаметром 10 нм та довжиною 5 нм. При подальшому стиску цієї спіралі до волокна діаметром 30 нм ступінь конденсації збільшується ще в шість разів.
Зрештою упаковка дуплексу ДНК із п'ятьма гістонами призводить до 50-кратної конденсації ДНК. Однак навіть такий високий ступінь конденсації не може пояснити майже 50 000 - 100 000-кратне ущільнення ДНК метафазної хромосомі. На жаль, деталі подальшої упаковки хроматину аж до метафазної хромосоми поки не відомі, тому можна розглядати лише загальні особливості цього процесу.
Рівні компактизації ДНК у хромосомах
Кожна молекула ДНК упакована в окрему хромосому. У диплоїдних клітинах людини міститься 46 хромосом, які знаходяться в ядрі клітини. Загальна довжина ДНК всіх хромосом клітини становить 1,74 м, проте діаметр ядра, в яке упаковані хромосоми, в мільйони разів менше. Таке компактне укладання ДНК у хромосомах і хромосом в ядрі клітини забезпечується різноманітними, гістоновими та негістоновими білками, що взаємодіють у певній послідовності з ДНК (див вище). Компактизація ДНК у хромосомах дозволяє зменшити її лінійні розміри приблизно в 10 000 разів – умовно з 5 см до 5 мкм. Вирізняють кілька рівнів компактизації (рис. 10).


- подвійна спіраль ДНК - негативно заряджена молекула діаметром 2 нм та завдовжки кілька див.
- нуклеосомний рівень- хроматин виглядає в електронному мікроскопі як ланцюжок "намистин" - нуклеосом - "на нитки". Нуклеосома - це універсальна структурна одиниця, яка виявляється як в еухроматині, так і в гетерохроматині, інтерфазному ядрі та метафазних хромосомах.
Нуклеосомний рівень компактизації забезпечується спеціальними білками – гістонами. Вісім позитивно заряджених гістонових доменів утворюють кор (серцевину) нуклеосоми, на яку намотується негативно заряджена молекула ДНК. Це дає скорочення в 7 разів, при цьому діаметр збільшується з 2 до 11 нм.
- соленоїдний рівень
Соленоїдний рівень організації хромосом характеризується скручуванням нуклеосомної нитки та утворенням з неї товстіших фібрил 20-35 нм у діаметрі - соленоїдів або супербідів. Крок соленоїда дорівнює 11 нм, однією виток припадає близько 6-10 нуклеосом. Соленоїдна упаковка вважається найбільш ймовірною, ніж супербідна, згідно з якою фібрила хроматину діаметром 20-35 нм є ланцюгом гранул, або супербідів, кожна з яких складається з восьми нуклеосом. На соленоїдному рівні лінійний розмір ДНК скорочується у 6-10 разів, діаметр збільшується до 30 нм.
- петльовий рівень
Петльовий рівень забезпечується негістоновими сайт-специфічними ДНК-зв'язуючими білками, які розпізнають певні послідовності ДНК та зв'язуються з ними, утворюючи петлі приблизно по 30-300 тисяч пар основ. Петля забезпечує експресію генів, тобто. петля є не лише структурною, а й функціональною освітою. Укорочення цьому рівні відбувається у 20-30 раз. Діаметр збільшується до 300 нм. Петлеподібні структури типу "лампових щіток" в земноводних ооцитах можна бачити на цитологічних препаратах. Ці петлі, мабуть, суперспіралізовані і є домени ДНК, відповідні, ймовірно, одиницям транскрипції та реплікації хроматину. Специфічні білки фіксують підстави петель і, можливо, деякі їхні внутрішні ділянки. Петлеподібна доменна організація сприяє укладанню хроматину в метафазних хромосомах у спіральні структури більш високих порядків.
- доменний рівень
Доменний рівень організації хромосом вивчений недостатньо. На даному рівні відзначається утворення петлевих доменів - структур з ниток (фібрил) товщиною 25-30 нм, які містять 60% білка, 35% ДНК і 5% РНК, практично не видно у всіх фазах клітинного циклу за винятком мітозу і дещо безладно розподілені по клітинному ядру. Петлеподібні структури типу "лампових щіток" в земноводних ооцитах можна бачити на цитологічних препаратах.
Петлеві домени своєю основою прикріплюються до внутрішньоядерного білкового матрикса в так званих вбудованих місцях прикріплення, що часто позначаються як MAR/SAR-послідовності (MAR, від англ. matrix associated region; SAR, від англ. scaffold attachment regions) – фрагментах ДНК протяжністю в кілька пар основ, що характеризуються високим вмістом (>65%) А/Т пар нуклеотидів. Кожен домен, мабуть, має одну точку початку реплікації та функціонує як автономна надспіральна одиниця. Будь-який петельний домен містить безліч одиниць транскрипції, функціонування яких, ймовірно, координується - весь домен знаходиться або в активному або неактивному стані.
На доменному рівні в результаті послідовної упаковки хроматину відбувається зменшення лінійних розмірів ДНК приблизно в 200 разів (700 нм).
- хромосомний рівень
На хромосомному рівні відбувається конденсація профазної хромосоми в метафазну з ущільнення петельних доменів навколо осьового каркасу негістонових білків. Ця суперспіралізація супроводжується фосфорилуванням у клітині всіх молекул H1. В результаті метафазну хромосому можна зобразити у вигляді щільно покладених соленоїдних петель, згорнутих у тугу спіраль. Типова хромосома людини може містити до 2600 петель. Товщина такої структури сягає 1400 нм (дві хроматиди), а молекула ДНК у своїй коротшується в 104 раз, тобто. із 5 см розтягнутої ДНК до 5 мкм.
Функції хромосом
У взаємодії із позахромосомними механізмами хромосоми забезпечують
- зберігання спадкової інформації
- використання цієї інформації для створення та підтримки клітинної організації
- регуляцію зчитування спадкової інформації
- самоподвоєння генетичного матеріалу
- передачу генетичного матеріалу від материнської клітини дочірнім.
Існують дані, що з активуванні ділянки хроматину, тобто. при транскрипції, з нього оборотно видаляються спочатку гістон H1, а потім октет гістонів. Це викликає деконденсацію хроматину, послідовний перехід 30-нанометрової фібрили хроматину в 10-нанометрову нитку та її подальше розгортання на ділянки вільної ДНК, тобто. втрату нуклеосомної структури.
Всі ми знаємо, що образ людини, деякі звички і навіть захворювання передаються у спадок. Уся ця інформація про живу істоту закодована в генах. То як же ці горезвісні гени виглядають, як вони функціонують і де знаходяться?
Отже, носієм усіх генів будь-якої людини чи тварини є ДНК. Ця сполука була відкрита в 1869 році Йоганном Фрідріхом Мішером. Хімічно ДНК - це дезоксирибонуклеїнова кислота. Що це означає? Яким чином ця кислота містить генетичний код всього живого на нашій планеті?
Почнемо з того, що розглянемо, де міститься ДНК. У клітині людини є безліч органоїдів, які виконують різні функції. ДНК міститься в ядрі. Ядро – це маленька органела, яка оточена спеціальною мембраною, і у якій зберігається весь генетичний матеріал – ДНК.
Яка будова молекули ДНК?
Насамперед, розглянемо, що є ДНК. ДНК – це дуже довга молекула, що складається із структурних елементів – нуклеотидів. Є 4 види нуклеотидів – це аденін (А), тимін (Т), гуанін (Г) та цитозин (Ц). Ланцюжок нуклеотидів схематично виглядає так: ГГААТЦТААГ.… Ось така послідовність нуклеотидів і є ланцюжок ДНК.Вперше структура ДНК була розшифрована в 1953 Джеймсом Вотсоном і Френсісом Криком.
В одній молекулі ДНК є два ланцюжки нуклеотидів, які спірально закручені навколо один одного. Які ж ці нуклеотидні ланцюжки тримаються поруч і закручуються в спіраль? Цей феномен обумовлений властивістю комплементарності. Комплементарність означає, що один навпроти одного у двох ланцюжках можуть бути лише певні нуклеотиди (комплементарні). Так, напроти аденіну завжди стоїть тимін, а напроти гуаніну завжди лише цитозин. Таким чином, гуанін комплементарний з цитозином, а аденін - з тиміном. Такі пари нуклеотидів, що стоять навпроти один одного в різних ланцюжках, також називаються комплементарними.
 Схематично можна зобразити так:
Схематично можна зобразити так:
Г - Ц
Т - А
Т - А
Ц - Г
Ці комплементарні пари А – Т та Г – Ц утворюють хімічний зв'язокміж нуклеотидами пари, причому зв'язок між Г і Ц більш міцний ніж між А і Т. Зв'язок утворюється суворо між комплементарними основами, тобто утворення зв'язку між некомлементарними Г і А - неможливе.
«Упаковка» ДНК, як ланцюжок ДНК стає хромосомою?
Чому ж ці нуклеотидні ланцюжки ДНК ще й закручуються довкола один одного? Навіщо це потрібно? Справа в тому, що кількість нуклеотидів є величезною і потрібно дуже багато місця, щоб розмістити такі довгі ланцюжки. З цієї причини відбувається спіральне закручування двох ниток ДНК навколо одного. Це явищеносить назву спіралізації. В результаті спіралізації ланцюжка ДНК коротшають у 5-6 разів.Деякі молекули ДНК активно використовуються організмом, інші використовуються рідко. Такі молекули ДНК, що рідко використовуються, крім спіралізації піддається ще більш компактній «упаковці». Така компактна упаковка називається суперспіралізацією та вкорочує нитку ДНК у 25-30 разів!
Як відбувається упакування спіралей ДНК?
 Для суперспіралізації використовуються гістонові білки, які мають вигляд та структуру стрижня або котушки для ниток. На ці «котушки» – гістонові білки намотуються спіралізовані нитки ДНК. Таким чином, довга нитка стає дуже компактно упакованою та займає дуже мало місця.
Для суперспіралізації використовуються гістонові білки, які мають вигляд та структуру стрижня або котушки для ниток. На ці «котушки» – гістонові білки намотуються спіралізовані нитки ДНК. Таким чином, довга нитка стає дуже компактно упакованою та займає дуже мало місця. За необхідності використовувати ту чи іншу молекулу ДНК відбувається процес «розкручування», тобто нитка ДНК «змотується» з «котушки» - гістонового білка (якщо була на неї накручена) і розкручується зі спіралі у два паралельні ланцюги. А коли молекула ДНК знаходиться в такому розкрученому стані, з неї можна вважати необхідну генетичну інформацію. Причому зчитування генетичної інформації відбувається лише з розкручених ниток ДНК!
Сукупність суперспіралізованих хромосом називається гетерохроматин, а хромосом, доступних для зчитування інформації – еухроматин.
Що таке гени, який їх зв'язок із ДНК?
Тепер давайте розглянемо, що таке гени. Відомо, що є гени, що визначають групу крові, колір очей, волосся, шкіри та безліч інших властивостей нашого організму. Ген – це строго певна ділянка ДНК, що складається з певної кількості нуклеотидів, розташованих у строго певній комбінації. Розташування в певному ділянці ДНК означає, що конкретному гену відведено його місце, і поміняти це місце неможливо. Доречно провести таке порівняння: людина живе на певній вулиці, у певному будинку та квартирі, і самовільно людина не може переселитися до іншого будинку, квартири чи іншої вулиці. Певна кількість нуклеотидів у гені означає, що кожен ген має конкретну кількість нуклеотидів і їх не може стати більшою або меншою. Наприклад, ген, що кодує вироблення інсуліну, складається з 60 пар нуклеотидів; ген, що кодує вироблення гормону окситоцину - з 370 пар нуклеотидів. Сувора послідовність нуклеотидів є унікальною для кожного гена та строго визначеною. Наприклад, послідовність ААТТААТА - це фрагмент гена, що кодує вироблення інсуліну. Щоб отримати інсулін, використовується саме така послідовність, для отримання, наприклад, адреналіну, використовується інша комбінація нуклеотидів. Важливо розуміти, що лише певна комбінація нуклеотидів кодує певний продукт (адреналін, інсулін тощо). Така ось унікальна комбінація певної кількості нуклеотидів, що стоїть на «своєму місці» - це і є ген.
Крім генів у ланцюзі ДНК розташовані, так звані «некодуючі послідовності». Такі некодуючі послідовності нуклеотидів регулюють роботу генів, допомагають спіралізації хромосом, відзначають точку початку та кінця гена. Однак, на сьогоднішній день, роль більшості послідовностей, що не кодують, залишається нез'ясованою.
Що таке хромосома? Статеві хромосоми
Сукупність генів індивіда називається геномом. Звичайно, весь геном неможливо укласти в одну ДНК. Геном розбито на 46 пар молекул ДНК. Одна пара молекул ДНК називається хромосомою. Так ось саме цих хромосом у людини є 46 штук. Кожна хромосома несе суворо певний набір генів, наприклад, в 18 хромосомі закладені гени, що кодують колір очей і т.д. Хромосоми відрізняються один від одного за довжиною та формою. Найпоширеніші форми як Х чи Y, але є й інші. У людини є дві хромосоми однакової форми, які називаються парними (парами). У зв'язку з такими відмінностями всі парні хромосоми пронумеровані - їх є 23 пари. Це означає, що є пара хромосом №1, пара №2, №3 тощо. Кожен ген відповідальний певний ознака перебуває у однієї й тієї ж хромосомі. У сучасних посібниках для фахівців може вказуватися локалізація гена, наприклад, таким чином: 22 хромосоми, довге плече.У чому різниця хромосом?
 Як ще відрізняються між собою хромосоми? Що означає термін довге плече? Візьмемо хромосоми форми Х. Перетин ниток ДНК може відбуватися строго посередині (Х), а може відбуватися і не центрально. Коли таке перетин ниток ДНК відбувається не центрально, то щодо точки перехреста одні кінці довші, інші, відповідно, коротші. Такі довгі кінці прийнято називати довгим плечем хромосоми, а короткі відповідно коротким плечем. У хромосом форми Y більшість займають довгі плечі, а короткі зовсім невеликі (на схематичному зображенні вони навіть не вказуються).
Як ще відрізняються між собою хромосоми? Що означає термін довге плече? Візьмемо хромосоми форми Х. Перетин ниток ДНК може відбуватися строго посередині (Х), а може відбуватися і не центрально. Коли таке перетин ниток ДНК відбувається не центрально, то щодо точки перехреста одні кінці довші, інші, відповідно, коротші. Такі довгі кінці прийнято називати довгим плечем хромосоми, а короткі відповідно коротким плечем. У хромосом форми Y більшість займають довгі плечі, а короткі зовсім невеликі (на схематичному зображенні вони навіть не вказуються). Розмір хромосом коливається: найбільшими є хромосоми пар №1 і №3, найменшими хромосоми пар №17, №19.
Крім форм і розмірів хромосоми розрізняються за функціями, що виконуються. З 23 пар, 22 пари є соматичними та 1 пара – статеві. Що це означає? Соматичні хромосоми визначають усі зовнішні ознаки індивідуума, особливості його поведінкових реакцій, Спадковий психотип, тобто всі риси та особливості кожної конкретної людини. А пара статевих хромосом визначає стать людини: чоловік або жінка. Існує два різновиди статевих хромосом людини – це Х (ікс) та У (ігрек). Якщо вони поєднуються як ХХ (ікс – ікс) – це жінка, а якщо ХУ (ікс – ігрек) – перед нами чоловік.
Спадкові хвороби та пошкодження хромосом
Проте трапляються «поломки» геному, тоді люди виявляються генетичні захворювання. Наприклад, коли в 21 парі хромосом замість двох присутні три хромосоми, людина народжується із синдромом Дауна.Існує безліч дрібніших «поломок» генетичного матеріалу, які не ведуть до виникнення хвороби, а навпаки, надають добрих властивостей. Усі «поломки» генетичного матеріалу називаються мутаціями. Мутації, які ведуть хвороб або погіршення властивостей організму, вважають негативними, а мутації, які ведуть освіту нових корисних властивостей, вважають позитивними.
Однак, стосовно більшості хвороб, якими сьогодні страждають люди, передається у спадок не захворювання, а лише схильність. Наприклад, у батька дитини цукор засвоюється повільно. Це не означає, що дитина народиться з цукровим діабетом, але у дитини буде схильність. Це означає, якщо дитина зловживатиме солодощами та борошняними виробами, то у нього розвинеться цукровий діабет.
На сьогоднішній день розвивається так звана предикативнамедицини. В рамках даної медичної практики у людини виявляються схильності (на основі виявлення відповідних генів), а потім їй даються рекомендації - якої дієти дотримуватися, як правильно чергувати режим праці та відпочинку, щоб не захворіти.
Як прочитати інформацію, закодовану в ДНК?
А як можна прочитати інформацію, що міститься в ДНК? Як використовує її власний організм? Сама ДНК є якоюсь матрицею, але не просту, а закодовану. Щоб прочитати інформацію з матриці ДНК, вона спочатку переноситься на спеціальний переносник – РНК. РНК – це хімічно рибонуклеїнова кислота. Відрізняється від ДНК тим, що може проходити через мембрану ядра в клітину, а ДНК позбавлена такої здатності (вона може бути тільки в ядрі). Закодована інформація використовується в самій клітині. Отже, РНК це переносник кодованої інформації з ядра в клітину.Як відбувається синтез РНК, як з допомогою РНК синтезується білок?
 Нитки ДНК, з яких потрібно "рахувати" інформацію, розкручуються, до них підходить спеціальний фермент - "будівельник" і синтезує паралельно нитки ДНК комплементарний ланцюжок РНК. Молекула РНК також складається з 4 видів нуклеотидів – аденіну (А), урацилу (У), гуаніну (Г) та цитозину (Ц). У цьому комплементарними є такі пари: аденін – урацил, гуанін – цитозин. Як видно, на відміну від ДНК, РНК використовується урацил замість тиміну. Тобто фермент-«будівельник» працює наступним чином: якщо в нитці ДНК він бачить А, то до нитки РНК приєднує У, якщо Г - то приєднує Ц і т.д. Таким чином, із кожного активного гена при транскрипції формується шаблон – копія РНК, здатна проходити через мембрану ядра.
Нитки ДНК, з яких потрібно "рахувати" інформацію, розкручуються, до них підходить спеціальний фермент - "будівельник" і синтезує паралельно нитки ДНК комплементарний ланцюжок РНК. Молекула РНК також складається з 4 видів нуклеотидів – аденіну (А), урацилу (У), гуаніну (Г) та цитозину (Ц). У цьому комплементарними є такі пари: аденін – урацил, гуанін – цитозин. Як видно, на відміну від ДНК, РНК використовується урацил замість тиміну. Тобто фермент-«будівельник» працює наступним чином: якщо в нитці ДНК він бачить А, то до нитки РНК приєднує У, якщо Г - то приєднує Ц і т.д. Таким чином, із кожного активного гена при транскрипції формується шаблон – копія РНК, здатна проходити через мембрану ядра. Як відбувається синтез білка, закодованого певним геном?
Залишивши ядро, РНК потрапляє до цитоплазми. Вже в цитоплазмі РНК може бути як матриця вбудована в спеціальні ферментні системи (рибосоми), які можуть синтезувати, керуючись інформацією РНК відповідну послідовність амінокислот білка. Як відомо, молекула білка складається з амінокислот. Як же рибосомі вдається дізнатися, яку саме амінокислоту треба приєднати до білкового ланцюга, що росте? Робиться це з урахуванням триплетного коду. Триплетний код означає, що послідовність три нуклеотиду ланцюжка РНК ( триплет,наприклад, ГГУ) кодують одну амінокислоту (у даному випадку гліцин). Кожну амінокислоту кодує певний триплет. І так, рибосома «прочитує» триплет, визначає яку амінокислоту треба приєднати наступною мірою зчитування інформації в РНК. Коли ланцюжок амінокислот сформована, вона приймає певну просторову форму і стає білком, здатним здійснювати покладені на нього ферментні, будівельні, гормональні та інші функції.Білок будь-якого живого організму є продуктом гена. Саме білками визначаються всі різноманітні властивості, якості та зовнішні прояви генів.
Справа найбільша спіраль ДНК людини, збудована з людей на пляжі у Варні (Болгарія), що увійшла до книги рекордів Гіннесса 23 квітня 2016 року
Дезоксирибонуклеїнова кислота. Загальні відомості
ДНК (дезоксирибонуклеїнова кислота) - своєрідне креслення життя, складний код, в якому укладені дані про спадкову інформацію. Ця складна макромолекула здатна зберігати та передавати спадкову генетичну інформацію з покоління до покоління. ДНК визначає такі властивості будь-якого живого організму як спадковість та мінливість. Закодована у ній інформація задає всю програму розвитку будь-якого живого організму. Генетично закладені фактори визначають весь перебіг життя як людини, так і будь-якого ін. організму. Штучний чи природний вплив зовнішнього середовища здатні лише незначною мірою вплинути на загальну вираженість окремих генетичних ознак або позначитися на розвитку запрограмованих процесів.
Дезоксирибонуклеїнова кислота(ДНК) - макромолекула (одна з трьох основних, дві інші - РНК та білки), що забезпечує зберігання, передачу з покоління в покоління та реалізацію генетичної програми розвитку та функціонування живих організмів. ДНК містить інформацію про структуру різних видів РНК та білків.
У клітинах еукаріотів (тварин, рослин та грибів) ДНК знаходиться в ядрі клітини у складі хромосом, а також у деяких клітинних органоїдах (мітохондріях і пластидах). У клітинах прокаріотичних організмів (бактерій та архей) кільцева чи лінійна молекула ДНК, так званий нуклеоїд, прикріплена зсередини до клітинної мембрани. У них і нижчих еукаріотів (наприклад, дріжджів) зустрічаються також невеликі автономні, переважно кільцеві молекули ДНК, звані плазмідами.
З хімічної точки зору ДНК - це довга полімерна молекула, що складається з блоків, що повторюються, - нуклеотидів. Кожен нуклеотид складається з азотистої основи, цукру (дезоксирибози) та фосфатної групи. Зв'язки між нуклеотидами в ланцюзі утворюються рахунок дезоксирибози ( З) та фосфатної ( Ф) групи (фосфодіефірні зв'язки).

Рис. 2. Нуклертид складається з азотистої основи, цукру (дезоксирибози) та фосфатної групи.
У переважній більшості випадків (крім деяких вірусів, що містять одноланцюжкову ДНК), макромолекула ДНК складається з двох ланцюгів, орієнтованих азотистими основами один до одного. Ця дволанцюжкова молекула закручена по гвинтовій лінії.
У ДНК зустрічається чотири види азотистих основ (аденін, гуанін, тимін та цитозин). Азотисті основи одного з ланцюгів з'єднані з азотистими основами іншого ланцюга водневими зв'язками згідно з принципом комплементарності: аденін з'єднується тільки з тиміном ( А-Т), гуанін - тільки з цитозином ( Г-Ц). Саме ці пари і становлять «перекладини» гвинтової "драбини" ДНК (див.: мал. 2, 3 та 4).

Рис. 2. Азотисті основи
Послідовність нуклеотидів дозволяє «кодувати» інформацію про різних типахРНК, найважливішими з яких є інформаційні, або матричні (мРНК), рибосомальні (рРНК) та транспортні (тРНК). Всі ці типи РНК синтезуються на матриці ДНК рахунок копіювання послідовності ДНК в послідовність РНК, синтезованої у процесі транскрипції, і беруть участь у біосинтезі білків (процесі трансляції). Крім кодуючих послідовностей, ДНК клітин містить послідовності, що виконують регуляторні та структурні функції.

Рис. 3. Реплікація ДНК
Розташування базових комбінацій хімічних сполук ДНК та кількісні співвідношення між цими комбінаціями забезпечують кодування спадкової інформації.
Освіта нової ДНК (реплікація)
- Процес реплікації: розкручування подвійної спіралі ДНК – синтез комплементарних ланцюгів ДНК-полімеразою – утворення двох молекул ДНК із однієї.
- Подвійна спіраль розстібається на дві гілки, коли ферменти руйнують зв'язок між базовими парами хімічних сполук.
- Кожна гілка є елементом нової ДНК. Нові базові пари з'єднуються у тій самій послідовності, що й батьківської гілки.
По завершенні дуплікації утворюються дві самостійні спіралі, створені з хімічних сполук батьківської ДНК і мають однаковий генетичний код. Таким шляхом ДНК здатна переривати інформацію від клітини до клітини.
Більш детальна інформація:
БУДОВА НУКЛЕЇНОВИХ КИСЛОТ

Рис. 4 . Азотисті основи: аденін, гуанін, цитозин, тимін
Дезоксирибонуклеїнова кислота(ДНК) відноситься до нуклеїнових кислот. Нуклеїнові кислоти– це клас нерегулярних біополімерів, мономерами яких є нуклеотиди.
НУКЛЕОТИДИскладаються з азотистої основи, сполученого з п'ятивуглецевим вуглеводом (пентозою) - дезоксирибозою(у разі ДНК) або рибозою(у разі РНК), який з'єднується із залишком фосфорної кислоти (H 2 PO 3 -).
Азотисті основибувають двох типів: піримідинові основи - урацил (тільки в РНК), цитозин і тимін, пуринові основи - аденін та гуанін.

Рис. 5. Структура нуклеотидів (ліворуч), розташування нуклеотиду в ДНК (знизу) та типи азотистих основ (праворуч): піримідинові та пуринові

Атоми вуглецю в молекулі пентози нумеруються числами від 1 до 5. Фосфат з'єднується з третім та п'ятим атомами вуглецю. Так нуклеїнотиди з'єднуються у ланцюг нуклеїнової кислоти. Таким чином, ми можемо виділити 3' та 5'-кінці ланцюга ДНК:

Рис. 6. Виділення 3' і 5'-кінців ланцюга ДНК
Два ланцюги ДНК утворюють подвійну спіраль. Ці ланцюги у спіралі зорієнтовані у протилежних напрямках. У різних ланцюгах ДНК азотисті основи з'єднані між собою за допомогою водневих зв'язків. Аденін завжди з'єднується з тиміном, а цитозин – з гуаніном. Це називається правилом комплементарності(Див. принцип комплементарності).
Правило комплементарності:
| A-T G-C |
Наприклад, якщо нам дано ланцюг ДНК, що має послідовність
3'- ATGTCCTAGCTGCTCG - 5',
то другий їй ланцюг буде комплементарний і спрямований у протилежному напрямку - від 5'-кінця до 3'-кінця:
5'-TACAGGATCGACGAGC-3'.

Рис. 7. Спрямованість ланцюгів молекули ДНК та з'єднання азотистих основ за допомогою водневих зв'язків
РЕПЛІКАЦІЯ ДНК
Реплікація ДНК– це процес подвоєння молекули ДНК шляхом матричного синтезу. Найчастіше природної реплікації ДНКпраймеромдля синтезу ДНК є короткий фрагмент (Створюється наново). Такий рибонуклеотидний праймер створюється ферментом праймазою (ДНК-праймаза у прокаріотів, ДНК-полімераза у еукаріотів), і згодом замінюється дезоксирибонуклеотидами полімеразою, що виконує в нормі функції репарації (виправлення хімічних пошкоджень та розривів у молеклі ДНК).
Реплікація відбувається за напівконсервативним механізмом. Це означає, що подвійна спіраль ДНК розплітається і кожному з її ланцюгів за принципом комплементарності добудовується новий ланцюг. Дочірня молекула ДНК, таким чином, містить у собі один ланцюг від материнської молекули та одну знову синтезовану. Реплікація відбувається у напрямку від 3' до 5' кінця материнської ланцюга.

Рис. 8. Реплікація (подвоєння) молекул ДНК
ДНК-синтез- це не такий складний процес, як може здатися на перший погляд. Якщо подумати, то для початку потрібно розібратися, що таке синтез. Це процес об'єднання чогось в одне ціле. Утворення нової молекули ДНК відбувається у кілька етапів:
1) ДНК-топоізомераза, розташовуючись перед вилкою реплікації, розрізає ДНК для того, щоб полегшити її розплетення та розкручування.
2) ДНК-хеліказу за топоізомеразою впливає процес «розплетення» спіралі ДНК.
3) ДНК-зв'язуючі білки здійснюють зв'язування ниток ДНК, а також проводять їхню стабілізацію, не допускаючи їх прилипання один до одного.
4) ДНК-полімераза δ(дельта) , погоджено зі швидкістю руху реплікативної вилки, що здійснює синтезведучоюланцюгидочірньої ДНК у напрямку 5"→3" на матриціматеринської нитки ДНК у напрямку від її 3"-кінця до 5"-кінця (швидкість до 100 пар нуклеотидів за секунду). Цією події на даній материнськоїнитки ДНК обмежуються.

Рис. 9. Схематичне зображення процесу реплікації ДНК: (1) Відстаючий ланцюг (запізнювальна нитка), (2) Ведучий ланцюг (лідируюча нитка), (3) ДНК-полімераза α (Polα), (4) ДНК-лігаза, (5) РНК -праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полімераза δ (Polδ ), (9) Хеліказа, (10) Однониткові ДНК-зв'язуючі білки, (11) Топоізомераза.
Далі описаний синтез відстаючого ланцюга дочірньої ДНК (див. Схемуреплікативної вилки та функції ферментів реплікації)
Наочніше про реплікацію ДНК див.
5) Безпосередньо відразу після розплетення та стабілізації іншої нитки материнської молекули до неї приєднуєтьсяДНК-полімераза α(альфа)та у напрямку 5"→3" синтезує праймер (РНК-затравку) - послідовність РНК на матриці ДНК довжиною від 10 до 200 нуклеотидів. Після цього ферментвидаляється з нитки ДНК.
Замість ДНК-полімеразиα
до 3"-кінця праймера приєднуєтьсяДНК-полімеразаε
.
6)
ДНК-полімеразаε
(епсілон) як би продовжує подовжувати праймер, але як субстрат вбудовуєдезоксирибонуклеотиди(у кількості 150-200 нуклеотидів). В результаті утворюється цільна нитка з двох частин -РНК(тобто праймер) та ДНК.
ДНК-полімераза εпрацює доти, доки не зустріне праймер попередньогофрагмента Оказаки(Синтезований трохи раніше). Після цього цей фермент видаляється з ланцюга.
7) ДНК-полімераза β(бета) встає замістьДНК-полімерази εрухається в тому ж напрямку (5"→3") і видаляє рибонуклеотиди праймера, одночасно вбудовуючи дезоксирибонуклеотиди на їхнє місце. Фермент працює до видалення праймера, тобто. поки на його шляху не встане дезоксирибонуклеотид (ще раніше синтезованийДНК-полімеразою ε). Зв'язати результат своєї роботи і попереду ДНК фермент, що стоїть, не в змозі, тому він сходить з ланцюга.
У результаті матриці материнської нитки " лежить " фрагмент дочірньої ДНК. Він називаєтьсяфрагмент Оказаки.
8) ДНК-лігаза робить зшивку двох сусідніх фрагментів Оказаки , тобто. 5"-кінця відрізка, синтезованогоДНК-полімеразою ε ,та 3"-кінця ланцюга, вбудованогоДНК-полімеразоюβ .
БУДОВА РНК
Рибонуклеїнова кислота(РНК) — одна з трьох основних макромолекул (дві інші — ДНК та білки), які містяться у клітинах усіх живих організмів.
Так само, як ДНК, РНК складається з довгого ланцюга, в якому кожна ланка називається нуклеотидом. Кожен нуклеотид складається з азотистої основи, цукру рибози та фосфатної групи. Однак, на відміну від ДНК, РНК зазвичай має не два ланцюги, а один. Пентоза в РНК представлена рибозою, а не дезоксирибозою (у рибози є додаткова гідроксильна група на другому атомі вуглеводу). Нарешті, ДНК відрізняється від РНК за складом азотистих основ: замість тиміну ( Т) в РНК представлений урацил ( U) , який також комплементарний аденіну.
Послідовність нуклеотидів дозволяє РНК кодувати генетичну інформацію. Усі клітинні організми використовують РНК (мРНК) для програмування синтезу білків.
Клітинні РНК утворюються під час процесу, званого транскрипцією , тобто синтезу РНК на матриці ДНК, що здійснюється спеціальними ферментами - РНК-полімеразами.
Потім матричні РНК (мРНК) беруть участь у процесі, званому трансляцією, тобто. синтезу білка на матриці мРНК з участю рибосом Інші РНК після транскрипції піддаються хімічним модифікаціям, і після утворення вторинної та третинної структур виконують функції, що залежать від типу РНК.

Рис. 10. Відмінність ДНК від РНК по азотистому підставі: замість тиміну (Т) в РНК представлений урацил (U), який також комплементарний аденіну.
ТРАНСКРИПЦІЯ
Це процес синтезу РНК на матриці ДНК. ДНК розкручується одному з ділянок. На одному з ланцюгів міститься інформація, яку необхідно скопіювати на молекулу РНК - цей ланцюг називається кодуючим. Другий ланцюг ДНК, що комплементарний кодує, називається матричною. У процесі транскрипції на матричному ланцюзі у напрямку 3' - 5' (ланцюгом ДНК) синтезується комплементарна їй ланцюг РНК. Таким чином, створюється РНК-копія кодуючого ланцюга.
![]()
Рис. 11. Схематичне зображення транскрипції
Наприклад, якщо нам дана послідовність кодуючого ланцюга
3'- ATGTCCTAGCTGCTCG - 5',
то, за правилом комплементарності, матричний ланцюг нестиме послідовність
5'- TACAGGATCGACGAGC- 3',
а синтезована з неї РНК – послідовність
ТРАНСЛЯЦІЯ
Розглянемо механізм синтезу білкана матриці РНК, а також генетичний код та його властивості. Також для наочності за наведеним нижче посиланням рекомендуємо подивитися невелике відео про процеси транскрипції та трансляції, що відбуваються в живій клітині:

Рис. 12. Процес синтезу білка: ДНК кодує РНК, РНК кодує білок
ГЕНЕТИЧНИЙ КІД
Генетичний код- спосіб кодування амінокислотної послідовності білків за допомогою нуклеотидної послідовності. Кожна амінокислота кодується послідовністю трьох нуклеотидів - кодоном або триплетом.
Генетичний код, загальний для більшості про- та еукаріотів. У таблиці наведено всі 64 кодони та зазначені відповідні амінокислоти. Порядок підстав - від 5" до 3" кінця мРНК.
Таблиця 1. Стандартний генетичний код
|
1-е ня |
2-а основа |
3-тє ня |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(Ser/S) |
U A U |
(Tyr/Y) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
U C A |
U A A |
Стоп-кодон** |
U G A |
Стоп-кодон** |
A |
||
|
U U G |
U C G |
U A G |
Стоп-кодон** |
U G G |
(Trp/W) |
G |
|||
|
C |
C U U |
C C U |
(Pro/P) |
C A U |
(His/H) |
C G U |
(Arg/R) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
C GA |
A |
||||
|
C U G |
C C G |
C A G |
C G G |
G |
|||||
|
A |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(Ser/S) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
A |
||||
|
A U G |
(Met/M) |
A C G |
A A G |
A G G |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
G U A |
G C A |
G A A |
(Glu/E) |
G G A |
A |
||||
|
G U G |
G C G |
G A G |
G G G |
G |
|||||
Серед триплетів є 4 спеціальні послідовності, що виконують функції «розділових знаків»:
- *Триплет AUG, також кодуючий метіонін, називається старт-кодоном. З цього кодону починається синтез молекули білка. Таким чином, під час синтезу білка першою амінокислотою в послідовності завжди буде метіонін.
- **Триплети UAA, UAGі UGAназиваються стоп-кодонамиі не кодують жодної амінокислоти. На цих послідовностях синтез білка припиняється.
Властивості генетичного коду
1. Триплетність. Кожна амінокислота кодується послідовністю трьох нуклеотидів - триплетом або кодоном.
2. Безперервність. Між триплетами немає додаткових нуклеотидів, інформація зчитується безперервно.
3. Неперекриваність. Один нуклеотид не може входити одночасно у два триплети.
4. Однозначність. Один кодон може кодувати лише одну амінокислоту.

5. Виродженість. Одна амінокислота може кодуватися кількома різними кодонами.

6. Універсальність. Генетичний код однаковий всім живих організмів.
приклад. Нам дана послідовність кодуючого ланцюга:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Матричний ланцюг матиме послідовність:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Тепер «синтезуємо» з цього ланцюга інформаційну РНК:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Синтез білка йде в напрямку 5' → 3', отже, нам потрібно перевернути послідовність, щоб прочитати генетичний код:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
Тепер знайдемо старт-кодон AUG:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
Розділимо послідовність на триплети:
звучить так: інформація з ДНК передається на РНК (транскрипція), з РНК - на білок (трансляція). ДНК може подвоюватися шляхом реплікації, і також можливий процес зворотної транскрипції, коли по матриці РНК синтезується ДНК, але такий процес в основному характерний для вірусів.

Рис. 13. Центральна догма молекулярної біології
ГЕНОМ: ГЕНИ та ХРОМОСОМИ
(загальні поняття)
Геном – сукупність всіх генів організму; його повний хромосомний набір.
Термін "геном" був запропонований Г. Вінклером у 1920 р. для опису сукупності генів, укладених у гаплоїдному наборі хромосом організмів одного біологічного виду. Початковий зміст цього терміна вказував на те, що поняття геному на відміну генотипу є генетичною характеристикою виду в цілому, а не окремої особи. З розвитком молекулярної генетики значення цього терміну змінилося. Відомо, що ДНК, яка є носієм генетичної інформації у більшості організмів і, отже, становить основу геному, включає не тільки гени в сучасному сенсі цього слова. Більша частинаДНК еукаріотичних клітин представлена некодуючими ("надмірними") послідовностями нуклеотидів, які не містять у собі інформації про білки та нуклеїнові кислоти. Таким чином, основну частину геному будь-якого організму становить вся ДНК його гаплоїдного набору хромосом.
Гени – це ділянки молекул ДНК, що кодують поліпептиди та молекули РНК.
За останнє сторіччя наше уявлення про гени суттєво змінилося. Раніше геном називали ділянку хромосоми, що кодує або визначає одну ознаку або фенотипічне(видима) властивість, наприклад колір очей.
У 1940 р. Джордж Бідл та Едвард Тейтем запропонували молекулярне визначення гена. Вчені обробляли суперечки гриба Neurospora crassaрентгенівським випромінюванням та іншими агентами, що викликають зміни у послідовності ДНК ( мутації), та виявили мутантні штами гриба, що втратили деякі специфічні ферменти, що в деяких випадках призводило до порушення цілого метаболічного шляху. Бідл і Тейтем дійшли висновку, що ген – це ділянка генетичного матеріалу, яка визначає чи кодує один фермент. Так виникла гіпотеза "один ген - один фермент". Пізніше ця концепція була розширена до визначення "один ген - один поліпептид"Оскільки багато генів кодують білки, що не є ферментами, а поліпептид може виявитися субодиницею складного білкового комплексу.
На рис. 14 показана схема того, як триплети нуклеотидів у ДНК визначають поліпептид - амінокислотну послідовність білка за посередництвом мРНК. Один із ланцюгів ДНК відіграє роль матриці для синтезу мРНК, нуклеотидні триплети (кодони) якої комплементарні триплетам ДНК. У деяких бактерій і багатьох еукаріотів послідовності, що кодують, перериваються некодуючими ділянками (так званими інтронами).
Сучасне біохімічне визначення гена ще конкретніше. Генами називаються всі ділянки ДНК, що кодують первинну послідовність кінцевих продуктів, до яких відносяться поліпептиди або РНК, що мають структурну або каталітичну функцію.
Поряд з генами ДНК містить інші послідовності, що виконують виключно регуляторну функцію. Регуляторні послідовностіможуть означати початок або кінець генів, впливати на транскрипцію або вказувати місце ініціації реплікації або рекомбінації. Деякі гени можуть експресуватися різними шляхами, при цьому одна і та ж ділянка ДНК служить матрицею для утворення різних продуктів.
Ми можемо приблизно розрахувати мінімальний розмір гена, що кодує середній білок Кожна амінокислота поліпептидного ланцюга кодується послідовністю з трьох нуклеотидів; послідовності цих триплетів (кодонів) відповідають ланцюжку амінокислот у поліпептиді, який кодується цим геном. Поліпептидний ланцюг із 350 амінокислотних залишків (ланцюг середньої довжини) відповідає послідовності з 1050 п.н. ( пар нуклеотидів). Однак багато генів еукаріотів і деякі гени прокаріотів перериваються сегментами ДНК, не несучими інформаціїпро білку, і тому виявляються значно довшими, ніж показує простий розрахунок.
Скільки генів в одній хромосомі?
 Рис. 15. Вид хромосом у прокарітичній (ліворуч) та еукаріотичній клеках. Гістони (Histones) — великий клас ядерних білків, виконують дві основні функції: вони беруть участь у упаковці ниток ДНК у ядрі та епігенетичної регуляції таких ядерних процесів, як транскрипція, реплікація і репарація.
Рис. 15. Вид хромосом у прокарітичній (ліворуч) та еукаріотичній клеках. Гістони (Histones) — великий клас ядерних білків, виконують дві основні функції: вони беруть участь у упаковці ниток ДНК у ядрі та епігенетичної регуляції таких ядерних процесів, як транскрипція, реплікація і репарація.
ДНК прокаріотів влаштована більш просто: їхні клітини не мають ядра, тому ДНК знаходиться безпосередньо в цитоплазмі у формі нуклеоїду.
 Як відомо, бактеріальні клітини мають хромосому у вигляді нитки ДНК, покладеної в компактну структуру – нуклеоїд. Хромосома прокаріоту Escherichia coli, чий геном повністю розшифрований, являє собою кільцеву молекулу ДНК (насправді це не правильне коло, а скоріше петля без початку і кінця), що складається з 4639675 п.н. У цій послідовності міститься приблизно 4300 генів білків та ще 157 генів стабільних молекул РНК. В геном людиниприблизно 3,1 млрд пар нуклеотидів, що відповідають майже 29 000 генам, розташованим на 24 різних хромосомах.
Як відомо, бактеріальні клітини мають хромосому у вигляді нитки ДНК, покладеної в компактну структуру – нуклеоїд. Хромосома прокаріоту Escherichia coli, чий геном повністю розшифрований, являє собою кільцеву молекулу ДНК (насправді це не правильне коло, а скоріше петля без початку і кінця), що складається з 4639675 п.н. У цій послідовності міститься приблизно 4300 генів білків та ще 157 генів стабільних молекул РНК. В геном людиниприблизно 3,1 млрд пар нуклеотидів, що відповідають майже 29 000 генам, розташованим на 24 різних хромосомах.
Прокаріоти (бактерії).
 Бактерія E. coliмає одну дволанцюжкову кільцеву молекулу ДНК. Вона складається з 4639675 п.н. і досягає в довжину приблизно 1,7 мм, що перевищує довжину самої клітини E. coliприблизно 850 раз. Крім великої кільцевої хромосоми у складі нуклеоїда багато бактерій містять одну або кілька маленьких кільцевих молекул ДНК, що вільно розташовуються в цитозолі. Такі позахромосомні елементи називають плазмідами(Рис. 16).
Бактерія E. coliмає одну дволанцюжкову кільцеву молекулу ДНК. Вона складається з 4639675 п.н. і досягає в довжину приблизно 1,7 мм, що перевищує довжину самої клітини E. coliприблизно 850 раз. Крім великої кільцевої хромосоми у складі нуклеоїда багато бактерій містять одну або кілька маленьких кільцевих молекул ДНК, що вільно розташовуються в цитозолі. Такі позахромосомні елементи називають плазмідами(Рис. 16).
Більшість плазмід складається всього з декількох тисяч пар нуклеотидів, деякі містять понад 10 000 п. н. Вони несуть генетичну інформацію та реплікуються з утворенням дочірніх плазмід, які потрапляють у дочірні клітини у процесі поділу батьківської клітини. Плазміди виявлені не тільки в бактеріях, але також у дріжджах та інших грибах. У багатьох випадках плазміди не дають жодних переваг клітинам-господарям, і їхнє єдине завдання — незалежне відтворення. Однак деякі плазміди несуть корисні для господаря гени. Наприклад, гени, що містяться в плазмідах, можуть надавати клітинам бактерій стійкість до антибактеріальних агентів. Плазміди, що несуть ген β-лактамази, забезпечують стійкість до β-лактамних антибіотиків, таких як пеніцилін та амоксицилін. Плазміди можуть переходити від клітин, стійких до антибіотиків, інших клітин того ж або іншого виду бактерій, в результаті чого ці клітини також стають резистентними. Інтенсивне застосування антибіотиків є потужним селективним фактором, що сприяє поширенню плазмід, що кодують стійкість до антибіотиків (а також транспозонів, що кодують аналогічні гени) серед хвороботворних бактерій, і призводить до появи бактеріальних штамів зі стійкістю до кількох антибіотиків. Лікарі починають розуміти небезпеку широкого використання антибіотиків і призначають їх лише у разі гострої потреби. З аналогічних причин обмежується широке використання антибіотиків на лікування сільськогосподарських тварин.
Див. також: Равін Н.В., Шестаков С.В. Геном прокаріотів // Вавиловський журнал генетики та селекції, 2013. Т. 17. № 4/2. С. 972-984.
Еукаріоти.
Таблиця 2. ДНК, гени та хромосоми деяких організмів
|
Загальна ДНК п.зв. |
Число хромосом* |
Орієнтовна кількість генів |
|
|
Escherichia coli(бактерія) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(дріжджі) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(Рослина) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(плодова мушка) |
120 367 260 |
20 000 |
|
|
Oryza sativa(Рис) |
480 000 000 |
57 000 |
|
|
Mus musculus(миша) |
2 634 266 500 |
27 000 |
|
|
Homo sapiens(людина) |
3 070 128 600 |
29 000 |
Примітка.Інформація постійно оновлюється; для отримання більш свіжої інформації зверніться до сайтів, присвячених окремим геномним проектам
* Для всіх еукаріотів, крім дріжджів, наводиться диплоїдний набір хромосом. Диплоїднийнабір хромосом (від грец. diploos-подвійний і eidos-вид) - подвійний набір хромосом(2n), кожна з яких має собі гомологічну.
**Гаплоїдний набір. Дикі штами дріжджів зазвичай мають вісім (октаплоїдний) або більше наборів таких хромосом.
***Для самок із двома Х хромосомами. У самців є Х хромосома, але немає Y, тобто всього 11 хромосом.
У клітці дріжджів, одних із найменших еукаріотів, у 2,6 рази більше ДНК, ніж у клітці E. coli(Табл. 2). Клітини плодової мушки Drosophila, класичного об'єкта генетичних досліджень, містять у 35 разів більше ДНК, а клітини людини – приблизно у 700 разів більше ДНК, ніж клітини E. coli.Багато рослин та амфібії містять ще більше ДНК. Генетичний матеріал клітин еукаріотів організований у вигляді хромосом. Диплоїдний набір хромосом (2 n) залежить від виду організму (табл. 2).
Наприклад, у соматичній клітині людини 46 хромосом ( Рис. 17). Кожна хромосома еукаріотичної клітини, як показано на рис. 17, амістить одну дуже велику двоспіральну молекулу ДНК. Двадцять чотири хромосоми людини (22 парні хромосоми і дві статеві хромосоми X та Y) розрізняються за довжиною більш ніж у 25 разів. Кожна хромосома еукаріотів містить певний набір генів.

Рис. 17. Хромосоми еукаріотів.а— пара пов'язаних та конденсованих сестринських хроматид із хромосоми людини. У такій формі еукаріотичні хромосоми перебувають після реплікації і в метафазі в процесі мітозу. б- Повний набір хромосом з лейкоциту одного з авторів книги. У кожній нормальній соматичній клітині людини міститься 46 хромосом.

Розмір та функція ДНК як матриці для зберігання та передачі спадкового матеріалу пояснюють наявність спеціальних структурних елементів в організації цієї молекули. У вищих організмів ДНК розподілено між хромосомами.
Сукупність ДНК (хромосом) організму називається геномом. Хромосоми знаходяться у клітинному ядрі та формують структуру, яка називається хроматином. Хроматин є комплексом ДНК і основних білків (гістонів) у співвідношенні 1:1. Довжину ДНК зазвичай вимірюють кількістю пар комплементарних нуклеотидів (п.н.). Наприклад, 3-я хромосомастоліття є молекулою ДНК розміром 160 млн п.н.. Виділена лінеаризована ДНК розміром 3*10 6 п.н. має довжину приблизно 1 мм, отже, лінеаризована молекула 3-ї хромосоми людини була б 5 мм у довжину, а ДНК всіх 23 хромосом (~3*10 9 п.н., MR = 1,8*10 12) гаплоїдної клітини - яйцеклітини або сперматозоїда - у лінеаризованому вигляді становила б 1 м. За винятком статевих клітин, всі клітини організму людини (їх близько 1013) містять подвійний набір хромосом. При клітинному поділі всі 46 молекул ДНК реплікуються і організуються в 46 хромосом.
Якщо з'єднати між собою молекули ДНК людського геному (22 хромосоми та хромосоми X та Y або Х та Х), вийде послідовність довжиною близько одного метра. Прим.: У всіх ссавців та інших організмів з гетерогаметною чоловічою статтю, у самок дві X-хромосоми (XX), а у самців - одна X-хромосома та одна Y-хромосома (XY).
Більшість клітин людини тому загальна довжина ДНК таких клітин близько 2м. У дорослої людини приблизно 1014 клітин, таким чином, загальна довжина всіх молекул ДНК становить 2.1011 км. Для порівняння, коло Землі - 4 10 4 км, а відстань від Землі до Сонця - 1,5 10 8 км. Ось як напрочуд компактно упакована ДНК у наших клітинах!
У клітинах еукаріотів є й інші органели, що містять ДНК, - це мітохондрії та хлоропласти. Висувалася безліч гіпотез щодо походження ДНК мітохондрій та хлоропластів. Загальновизнана сьогодні точка зору полягає в тому, що вони є рудиментами хромосом древніх бактерій, які проникли в цитоплазму господарських клітин і стали попередниками цих органел. Мітохондріальна ДНК кодує мітохондріальні тРНК та рРНК, а також кілька мітохондріальних білків. Понад 95% мітохондріальних білків кодується ядерною ДНК.
БУДОВА ГЕНІВ
Розглянемо будову гена у прокаріотів та еукаріотів, їх подібності та відмінності. Незважаючи на те, що ген - це ділянка ДНК, що кодує всього один білок або РНК, крім безпосередньо кодує частини, він також включає регуляторні та інші структурні елементи, що мають різну будову у прокаріотів і еукаріотів.
Кодуюча послідовність- основна структурно-функціональна одиниця гена, саме в ній знаходяться триплети нуклеотидів, що кодуютьамінокислотну послідовність. Вона починається зі старт-кодону та закінчується стоп-кодоном.
До та після кодуючої послідовності знаходяться нетрансльовані 5'- та 3'-послідовності. Вони виконують регуляторні та допоміжні функції, наприклад, забезпечують посадку рибосоми на іРНК.
Нетрансльовані і кодуюча послідовності складають одиницю транскрипції - ділянку ДНК, що транскрибується, тобто ділянка ДНК, з якої відбувається синтез і-РНК.
Термінатор- Нетранскрибується ділянка ДНК в кінці гена, на якому зупиняється синтез РНК.
На початку гена знаходиться регуляторна область, Що включає в себе промоторі оператор.
Промотор- послідовність, з якою пов'язується полімераза у процесі ініціації транскрипції. Оператор- це область, з якою можуть зв'язуватись спеціальні білки - репресори, які можуть зменшувати активність синтезу РНК з цього гена - інакше кажучи, зменшувати його експресію.
Будова генів у прокаріотів
Загальний план будови генів у прокаріотів і еукаріотів не відрізняється - і ті, й інші містять регуляторну область з промотором і оператором, одиницю транскрипції з послідовностями, що кодують і нетрансльовані, і термінатор. Проте організація генів у прокаріотів та еукаріотів відрізняється.

Рис. 18. Схема будови гена у прокаріотів (бактерій) -зображення збільшується
На початку та в кінці оперону є єдині регуляторні області для кількох структурних генів. З ділянки, що транскрибується, оперона зчитується одна молекула і-РНК, яка містить кілька кодуючих послідовностей, в кожній з яких є свій старт-і стоп-кодон. З кожної з таких ділянок зінтезується один білок. Таким чином, з однієї молекули і РНК синтезується кілька молекул білка.
Для прокаріотів характерне поєднання кількох генів в єдину функціональну одиницю. оперон. Роботу оперону можуть регулювати інші гени, які можуть бути помітно віддалені від самого оперону. регулятори. Білок, що транслюється з цього гена називається репресор. Він зв'язується з оператором оперону, регулюючи експресію відразу всіх генів, що в ньому містяться.
Для прокаріотів також характерне явище сполучення транскрипції та трансляції.
![]()
Рис. 19 Явище сполучення транскрипції та трансляції у прокаріотів - зображення збільшується
Таке сполучення не зустрічається у еукаріотів через наявність у них ядерної оболонки, що відокремлює цитоплазму, де відбувається трансляція, від генетичного матеріалу, на якому відбувається транскрипція. У прокаріотів під час синтезу РНК на матриці ДНК із молекулою РНК, що синтезується, може відразу зв'язуватися рибосома. Таким чином, трансляція починається ще до завершення транскрипції. Понад те, з однією молекулою РНК може одночасно зв'язуватися кілька рибосом, синтезуючи відразу кілька молекул одного білка.
Будова генів у еукаріотів
Гени та хромосоми еукаріотів дуже складно організовані
У бактерій багатьох видів лише одна хромосома, і майже у всіх випадках у кожній хромосомі присутня по одній копії кожного гена. Лише деякі гени, наприклад гени рРНК, містяться у кількох копіях. Гени та регуляторні послідовності становлять майже весь геном прокаріотів. Понад те, майже кожен ген суворо відповідає амінокислотної послідовності (чи послідовності РНК), що він кодує (рис. 14).
Структурна та функціональна організація генів еукаріотів набагато складніше. Дослідження хромосом еукаріотів, а пізніше секвенування повних послідовностей геномів еукаріотів принесло багато сюрпризів. Багато, якщо не більшість, генів еукаріотів мають цікаву особливість: їх нуклеотидні послідовності містять одну або кілька ділянок ДНК, в яких не кодується амінокислотна послідовність поліпептидного продукту. Такі нетрансльовані вставки порушують пряму відповідність між нуклеотидною послідовністю гена і амінокислотною послідовністю поліпептиду, що кодується. Ці нетрансльовані сегменти у складі генів називають інтронами, або вбудованими послідовностями, а кодуючі сегменти екзонами. У прокаріотів лише деякі гени містять інтрони.
Отже, у еукаріотів практично не зустрічається об'єднання генів в оперони, і кодуюча послідовність гена еукаріотів найчастіше розділена на ділянки, що транслюються. - екзони, та нетрансльовані ділянки - інтрони.
Найчастіше функція інтронів не встановлена. Загалом, лише близько 1,5% ДНК людини є «кодуючими», тобто несуть інформацію про білки або РНК. Проте з урахуванням великих інтронів виходить, що ДНК людини на 30% складається із генів. Оскільки гени становлять відносно невелику частку геномі людини, значна частина ДНК залишається неврахованою.

Рис. 16. Схема будова гена у еукаріотів - зображення збільшується
З кожного гена спочатку синтезується незріла, або пре-РНК, яка містить як інтрони, так і екзони.
Після цього проходить процес сплайсингу, в результаті якого інтронні ділянки вирізаються, і утворюється зріла іРНК, з якої може бути синтезований білок.

Рис. 20. Процес альтернативного сплайсингу - зображення збільшується
Така організація генів дозволяє, наприклад, здійснити, коли з одного гена можуть бути синтезовані різні форми білка, за рахунок того, що в процесі сплайсингу екзони можуть зшиватися в різних послідовностях.

Рис. 21. Відмінності у будові генів прокаріотів та еукаріотів - зображення збільшується
МУТАЦІЇ ТА МУТАГЕНЕЗ
Мутацієюназивається стійка зміна генотипу, тобто зміна нуклеотидної послідовності.
Процес, що призводить до виникнення мутацій називається мутагенезом, а організм, Усеклітини якого несуть ту саму мутацію. мутантом.
Мутаційна теоріябула вперше сформульована Гуго де Фрізом у 1903 році. Сучасний її варіант включає наступні положення:
1. Мутації виникають раптово, стрибкоподібно.
2. Мутації передаються з покоління до покоління.
3. Мутації можуть бути корисними, шкідливими чи нейтральними, домінантними чи рецесивними.
4. Можливість виявлення мутацій залежить від кількості досліджених особин.
5. Подібні мутації можуть бути повторно.
6. Мутації не спрямовані.
Мутації можуть бути під впливом різних чинників. Розрізняють мутації, що виникли під дією мутагенних впливів: фізичних (наприклад, ультрафіолету або радіації), хімічних (наприклад, колхіцину або активних форм кисню) та біологічних (наприклад, вірусів). Також мутації можуть бути спричинені помилками реплікації.
Залежно та умовами появи мутації поділяють на спонтанні- тобто мутації, що виникли в нормальних умовах, і індуковані— тобто мутації, що виникли за особливих умов.
Мутації можуть виникати не тільки в ядерній ДНК, але і, наприклад, ДНК мітохондрій або пластид. Відповідно, ми можемо виділяти ядерніі цитоплазматичнімутації.
Внаслідок виникнення мутацій часто можуть з'являтися нові алелі. Якщо мутантний аллель пригнічує дію нормальної, мутація називається домінантною. Якщо нормальний аллель пригнічує мутантний, така мутація називається рецесивний. Більшість мутацій, що призводять до нових алелів є рецесивними.
За ефектом виділяють мутації адаптивні, що призводять до підвищення пристосованості організму до середовища, нейтральні, що не впливають на виживання, шкідливі, що знижують пристосованість організмів до умов середовища та летальні, що призводять до смерті організму на ранніх стадіях розвитку
За наслідками виділяються мутації, що призводять до втрати функції білка, мутації, що призводять до виникненню у білка нової функції, а також мутації, які змінюють дозу гена, і, відповідно, дозу синтезованого білка з нього.
Мутація може виникнути до будь-якої клітини організму. Якщо мутація виникає у статевій клітині, вона називається гермінативною(Гермінальною, або генеративною). Такі мутації не виявляються в організму, у якого вони з'явилися, але призводять до появи мутантів у потомстві і передаються у спадок, тому вони важливі для генетики та еволюції. Якщо мутація виникає у будь-якій іншій клітині, вона називається соматичної. Така мутація може тією чи іншою мірою проявлятися у того організму, у якого вона виникла, наприклад, призводити до утворення ракових пухлин. Однак така мутація не передається у спадок і не впливає на нащадків.
Мутації можуть зачіпати різні за розміром ділянки геному. Виділяють генні, хромосомніі геномнімутації.
Генні мутації
Мутації, що виникають у масштабі меншому, ніж один ген, називаються генними, або точковими (точковими). Такі мутації призводять до зміни одного та декількох нуклеотидів у послідовності. Серед генних мутацій виділяютьзаміни, що призводять до заміни одного нуклеотиду на інший,делеції, що призводять до випадання одного з нуклеотидів,інсерції, що призводять до додавання зайвого нуклеотиду в послідовність

Рис. 23. Генні (точкові) мутації
За механізмом на білок, генні мутації ділять на:синонімічні, які (в результаті виродженості генетичного коду) не призводять до зміни амінокислотного складу білкового продукту,міссенс-мутації, які призводять до заміни однієї амінокислоти на іншу і можуть впливати на структуру білка, що синтезується, хоча часто вони виявляються незначними,нонсенс-мутації, що призводять до заміни кодуючого кодону на стоп-кодон,мутації, що призводять до порушення сплайсингу:

Рис. 24. Схеми мутацій
Також за механізмом на білок виділяють мутації, що призводять до зсуву рамки зчитування, наприклад, інсерції та делеції. Такі мутації, як і нонсенс-мутації, хоч і виникають в одній точці гена, часто впливають на всю структуру білка, що може призвести до повної зміни його структури. коли ділянка хромосоми повертається на 180 градусів, Рис. 28. Транслокація
коли ділянка хромосоми повертається на 180 градусів, Рис. 28. Транслокація
Рис. 29. Хромосома до та після дуплікації
Геномні мутації
Зрештою, геномні мутаціїзачіпають весь геном повністю, тобто змінюється кількість хромосом. Виділяють поліплоїдії - збільшення плоїдності клітини і анеуплоїдії, тобто зміна кількості хромосом, наприклад, трисомії (наявність в однієї з хромосом додаткового гомолога) і моносомії (відсутність у хромосоми гомолога).
Відео на тему ДНК
РЕПЛІКАЦІЯ ДНК, КОДИРУВАННЯ РНК, СИНТЕЗ БІЛКУ
(Якщо відео не відображається воно доступне за
За своєю хімічною будовою ДНК ( дезоксирибонуклеїнова кислота) є біополімером, мономерами якого є нуклеотиди. Тобто ДНК – це полінуклеотид. Причому молекула ДНК зазвичай складається з двох ланцюгів, закручених один щодо одного по гвинтовій лінії (часто кажуть «спірально закручених») та з'єднаних між собою водневими зв'язками.
Ланцюжки можуть бути закручені як у ліву, так і в праву (найчастіше) бік.
У деяких вірусів ДНК складається з одного кола.
Кожен нуклеотид ДНК складається з 1) азотистої основи, 2) дезоксирибози, 3) залишку фосфорної кислоти.
Подвійна правозакручена спіраль ДНК
До складу ДНК входять такі: аденін, гуанін, тиміні цитозин. Аденін і гуанін відносяться до пуринам, а тимін та цитозин - до піримідинам. Іноді до складу ДНК входить урацил, який зазвичай характерний для РНК де заміщає тимін.
Азотисті основи одного ланцюга молекули ДНК з'єднуються з азотистими основами іншого строго за принципом комплементарності: аденін тільки з тиміном (утворюють між собою два водневі зв'язки), а гуанін лише з цитозином (три зв'язки).
Азотиста основа в самому нуклеотиді пов'язана з першим атомом вуглецю циклічної форми дезоксирибозияка є пентозою (вуглеводом з п'ятьма атомами вуглецю). Зв'язок є ковалентним, глікозидним (C-N). На відміну від рибози, у дезоксирибози відсутня одна з гідроксильних груп. Кільце дезоксирибози формують чотири атоми вуглецю та один атом кисню. П'ятий атом вуглецю знаходиться поза кільцем і з'єднаний через атом кисню із залишком фосфорної кислоти. Також через атом кисню третього атома вуглецю приєднується залишок фосфорної кислоти сусіднього нуклеотиду.
Таким чином, в одному ланцюгу ДНК сусідні нуклеотиди пов'язані між собою ковалентними зв'язкамиміж дезоксирибозою та фосфорною кислотою (фосфодіефірний зв'язок). Утворюється фосфат-дезоксирибозний кістяк. Перпендикулярно йому, назустріч іншому ланцюжку ДНК, спрямовані азотисті основи, які з'єднуються з основами другого ланцюжка водневими зв'язками.
Будова ДНК така, що кістяки з'єднаних водневими зв'язками ланцюжків спрямовані в різні боки (кажуть «різноспрямовані», «антипаралельні»). З того боку, де одна закінчується фосфорною кислотою, сполученою з п'ятим атомом вуглецю дезоксирибози, інша закінчується «вільним» третім атомом вуглецю. Тобто кістяк одного ланцюжка перевернуть як би з ніг на голову щодо іншого. Таким чином, у будові ланцюжків ДНК розрізняють 5"-кінці та 3"-кінці.
При реплікації (подвоєння) ДНК синтез нових ланцюжків завжди йде від їх 5-го кінця до третього, тому що нові нуклеотиди можуть приєднуватися тільки до третього вільного кінця.
Зрештою (опосередковано через РНК) кожні три нуклеотиди в ланцюгу ДНК, що йдуть поспіль, кодують одну амінокислоту білка.
Відкриття будови молекули ДНК відбулося в 1953 завдяки роботам Ф. Крику і Д. Уотсона (чому також сприяли ранні роботи інших вчених). Хоча як хімічна речовинаДНК було відомо ще у ХІХ столітті. У 40-х роках XX століття стало зрозумілим, що саме ДНК є носієм генетичної інформації.
Подвійна спіраль вважається вторинною структурою молекули ДНК. У клітинах еукаріотів переважна кількість ДНК знаходиться в хромосомах, де пов'язана з білками та іншими речовинами, а також піддається більш щільній упаковці.
Дезоксирибонуклеїнова кислота або ДНК є носієм генетичної інформації. Головним чином ДНК у клітинах зосереджена в ядрі. Це є основним компонентом хромосом. У еукаріотів ДНК також виявляється ще в мітохондріях і пластидах. ДНК складається з мононуклеотидів, ковалентно пов'язаних між собою, являючи собою довгий нерозгалужений полімер. Мононуклеотиди, що входять до складу ДНК, складаються з дезоксирибози, однієї з 4-х азотистих основ (аденін, гуанін, цитозин та тимін), та залишку фосфорної кислоти. Кількість цих мононуклеотидів дуже велика. Наприклад, у клітинах прокаріотів, що містять одну єдину хромосому, ДНК являє собою одну макромолекул з молекулярною масою більше 2 x 10 9 .
Мононуклеотиди одного ланцюга ДНК з'єднуються послідовно один з одним завдяки утворенню ковалентних фосфодіефірних зв'язків між ОН-групою дезоксирибози одного мононуклеотиду та залишком фосфорної кислоти іншого. З одного боку від остова одного ланцюга ДНК, що утворився, розташовуються азотисті основи. Їх можна порівняти, з чотирма різними бусинками одягнених однією нитку, т.к. вони нанизані на сахарофосфатную ланцюг.
Виникає питання, як цей довгий полінуклеотидний ланцюг може кодувати програму розвитку клітини або навіть цілого організму? Відповідь це питання можна отримати, зрозумівши, як утворюється просторова структура ДНК. Структура цієї молекули була розшифрована та описана Дж. Уотсоном та Ф. Криком у 1953 році.
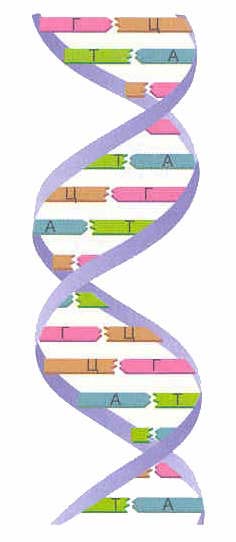
Молекули ДНК є дві нитки, які розташовуються паралельно одна одній і формують правозакручену спіраль
. Ширина цієї спіралі становить близько 2 нм, зате її довжина може сягати сотень тисяч нанометрів. Уотсоном і Криком запропонували модель ДНК, згідно з якою всі підстави ДНК розташовані всередині спіралі, зовні знаходиться цукрофосфатний кістяк. Таким чином, основи одного ланцюга максимально зближені з основами іншого,
тому між ними формуються водневі зв'язки. Структура спіралі ДНК така, що полінуклеотидні ланцюги, що входять до її складу, можуть бути розділені лише після її розкручування.
Завдяки максимальній зближеності двох ланцюгів ДНК у її складі міститься однакова кількість азотистих основ одного типу (аденін та гуанін) та азотистих основ іншого типу (тимін та цитозин), тобто справедлива формула: А+Г=Т+Ц. Це пояснюється розмірами азотистих основ, а саме, довжина структур, що утворюються завдяки виникненню водневого зв'язку між парами аденін-тимін та гуанін-цитозин, приблизно становить 1,1 нм. Сумарні розміри цих пар відповідають розмірам внутрішньої частини спіралі ДНК. Для формування спіралі пара Ц-Тбула б надто мала, а пара А-Г, Навпаки, занадто велика. Тобто, азотна основа першого ланцюга ДНК, визначає основу, яка розташовується в тому ж самому місці іншого ланцюга ДНК. Сувора відповідність нуклеотидів, розташованих у молекулі ДНК у парних ланцюжках паралельно один одному, назвали комплементарністю (Додатковістю). Точне відтворення або реплікація генетична інформація можлива саме завдяки цій особливості молекули ДНК.
У ДНК біологічна інформація записана таким чином, що вона може точно копіюватися і передаватися клітинам-нащадкам. До розподілу клітини у ній відбувається реплікація (самоподвоєння ) ДНК. Оскільки кожен ланцюг містить послідовність нуклеотидів, комплементарну послідовності цепіпартнера, то насправді вони мають однакову генетичну інформацію. Якщо розділити ланцюги і використовувати кожну з них як шаблон (матриця) для побудови другого ланцюга, то вийде два нових ідентичних ланцюга ДНК. Саме в такий спосіб і відбувається подвоєння ДНК у клітині.


