DNA neyden yapılmıştır. Nükleotid - bu nedir? DNA zincirindeki nükleotitlerin bileşimi, yapısı, sayısı ve dizisi DNA çift sarmalı, iyonlar tarafından stabilize edilir
Nükleik asitler, 3",5" - fosfodiester bağları kullanılarak bir polimer zincirinde birbirine bağlanan ve hücrelerde belirli bir şekilde paketlenen mononükleotitlerden oluşan makromoleküler maddelerdir.
Nükleik asitler iki çeşit biyopolimerdir: ribonükleik asit (RNA) ve deoksiribonükleik asit (DNA). Her biyopolimer, karbonhidrat kalıntısı (riboz, deoksiriboz) ve azotlu bazlardan biri (urasil, timin) bakımından farklılık gösteren nükleotidlerden oluşur. Buna göre, nükleik asitler isimlerini aldı.
Deoksiribonükleik asidin yapısı
Nükleik asitler birincil, ikincil ve üçüncül yapılara sahiptir.
DNA'nın birincil yapısı
DNA'nın birincil yapısı, mononükleotitlerin 3", 5" fosfodiester bağlarıyla bağlandığı doğrusal bir polinükleotit zinciridir. Bir hücrede bir nükleik asit zincirinin birleştirilmesi için başlangıç malzemesi, fosforik asidin β ve y kalıntılarının çıkarılmasının bir sonucu olarak, başka bir nükleosidin 3'-karbon atomunu bağlayabilen nükleosit 5'-trifosfattır. . Böylece, bir deoksiribozun 3" karbon atomu, bir fosforik asit kalıntısı yoluyla başka bir deoksiribozun 5" karbon atomuna kovalent olarak bağlanır ve lineer bir nükleik asit polinükleotit zinciri oluşturur. Bu nedenle adı: 3", 5"-fosfodiester bağları. Azotlu bazlar, bir zincirin nükleotitlerinin bağlantısında yer almaz (Şekil 1.).
Bir nükleotidin fosforik asit kalıntısı ile diğerinin karbonhidratı arasındaki böyle bir bağlantı, polinükleotit molekülünün bir pentoz-fosfat omurgasının oluşumuna yol açar, bunun üzerine azotlu bazlar arka arkaya yandan eklenir. Nükleik asit moleküllerinin zincirlerindeki dizileri, kesinlikle farklı organizmaların hücrelerine, yani. belirli bir karakteri vardır (Chargaff kuralı).
Uzunluğu zincire dahil olan nükleotit sayısına bağlı olan lineer bir DNA zincirinin iki ucu vardır: biri 3 "uç olarak adlandırılır ve bir serbest hidroksil içerir ve diğeri 5" ucu bir fosforik asit içerir. kalıntı. Devre polar olup 5"->3" ve 3"->5" olabilir. Bir istisna dairesel DNA'dır.
DNA'nın genetik "metni" kod "kelimelerinden" oluşur - kodon adı verilen nükleotidlerin üçlüleri. Tüm RNA türlerinin birincil yapısı hakkında bilgi içeren DNA bölümlerine yapısal genler denir.
Polinükleoditik DNA zincirleri devasa boyutlara ulaşır, bu nedenle hücre içinde belirli bir şekilde paketlenirler.
DNA'nın bileşimini inceleyen Chargaff (1949), bireysel DNA bazlarının içeriği ile ilgili önemli düzenlilikler belirledi. DNA'nın ikincil yapısını ortaya çıkarmaya yardımcı oldular. Bu kalıplara Chargaff kuralları denir. Chargaff kuralları
Bu kurallar, DNA oluştururken, genel olarak pürin ve pirimidin bazları için değil, özellikle adenin ile timin ve guanin ile sitozin için oldukça katı bir yazışma (eşleşme) gözlemlenmesi gerektiğini söyler. Bu kurallara dayanarak, 1953'te Watson ve Crick, DNA'nın çift sarmal adı verilen ikincil yapısının bir modelini önerdiler (Şekil). |

DNA'nın ikincil yapısı
DNA'nın ikincil yapısı, modeli 1953'te D. Watson ve F. Crick tarafından önerilen çift sarmaldır.
DNA modeli oluşturmak için ön koşullar
İlk analizlerin bir sonucu olarak, fikir, herhangi bir kökenden gelen DNA'nın, dört nükleotidin tümünü eşit molar miktarlarda içerdiğiydi. Ancak 1940'lı yıllarda E. Chargaff ve meslektaşları, çeşitli organizmalardan izole edilen DNA'nın analizleri sonucunda, içlerinde çeşitli kantitatif oranlarda azotlu bazların bulunduğunu açıkça göstermiştir. Chargaff, bu oranların aynı organizma türünün tüm hücrelerinden alınan DNA için aynı olmasına rağmen, farklı türlerden gelen DNA'nın belirli nükleotidlerin içeriğinde önemli ölçüde farklılık gösterebileceğini buldu. Bu, azotlu bazların oranındaki farklılıkların bazı biyolojik kodlarla ilgili olabileceğini düşündürdü. Farklı DNA örneklerinde tek tek pürin ve pirimidin bazlarının oranı aynı olmasa da, analizlerin sonuçları karşılaştırılırken belirli bir model ortaya çıktı: tüm örneklerde toplam pürin miktarı toplam pirimidin miktarına eşitti ( A + G = T + C), adenin miktarı timin miktarına (A = T) ve guanin miktarı - sitozin miktarına (G = C) eşittir. Memeli hücrelerinden izole edilen DNA, genellikle adenin ve timin açısından daha zengin ve guanin ve sitozin açısından nispeten daha zayıfken, bakterilerden elde edilen DNA, guanin ve sitozin açısından daha zengin ve adenin ve timin açısından nispeten daha fakirdi. Bu veriler, daha sonra Watson-Crick DNA yapı modelinin inşa edildiği temele dayanan gerçek materyalin önemli bir bölümünü oluşturdu.
DNA'nın olası yapısının bir başka önemli dolaylı göstergesi, L. Pauling'in protein moleküllerinin yapısına ilişkin verileriydi. Pauling, bir protein molekülünde amino asit zincirinin birkaç farklı kararlı konfigürasyonunun mümkün olduğunu gösterdi. Peptit zincirinin ortak konfigürasyonlarından biri - a-sarmal - düzenli bir sarmal yapıdır. Böyle bir yapı ile zincirin bitişik dönüşlerinde bulunan amino asitler arasında hidrojen bağlarının oluşması mümkündür. Pauling, 1950'de polipeptit zincirinin a-sarmal konfigürasyonunu tanımladı ve DNA moleküllerinin muhtemelen hidrojen bağları ile sabitlenmiş sarmal bir yapıya sahip olduğunu öne sürdü.
Ancak DNA molekülünün yapısı hakkında en değerli bilgi, X-ışını kırınım analizi sonuçlarıyla sağlandı. Bir DNA kristalinden geçen X ışınları kırınıma uğrar, yani belirli yönlerde sapar. Işınların sapma derecesi ve doğası, moleküllerin kendi yapılarına bağlıdır. X-ışını kırınım modeli (Şekil 3) deneyimli göze incelenen maddenin moleküllerinin yapısı ile ilgili bir dizi dolaylı gösterge verir. DNA X-ışını kırınım modellerinin analizi, azotlu bazların (düz bir şekle sahip) bir plaka yığını gibi istiflendiği sonucuna yol açtı. X-ışını desenleri, kristal DNA'nın yapısındaki üç ana periyodu tanımlamayı mümkün kıldı: 0.34, 2 ve 3.4 nm.

Watson-Crick DNA Modeli
Watson ve Crick, Chargaff'ın analitik verilerinden, Wilkins'in x-ışınlarından ve bir moleküldeki atomlar arasındaki kesin mesafeler, belirli bir atomun bağları arasındaki açılar ve atomların boyutu hakkında kesin bilgi sağlayan kimyagerlerden başlayarak, belirli bir ölçekte DNA molekülünün tek tek bileşenlerinin fiziksel modellerini oluşturmak ve elde edilen sistem çeşitli deneysel verilere karşılık gelecek şekilde bunları birbirine "ayarlamak" [göstermek] .
Daha önceleri, bir DNA zincirindeki bitişik nükleotidlerin, bir nükleotidin deoksiribozun 5'-karbon atomunu bir sonraki nükleotidin deoksiribozunun 3'-karbon atomuna bağlayan fosfodiester köprüleriyle bağlı olduğu biliniyordu. Watson ve Crick'in 0.34 nm'lik bir periyodun bir DNA zincirindeki ardışık nükleotitler arasındaki mesafeye karşılık geldiğinden hiç şüphesi yoktu. Ayrıca, 2 nm'lik periyodun zincirin kalınlığına karşılık geldiği varsayılabilir. Ve 3.4 nm'lik periyodun hangi gerçek yapıya tekabül ettiğini açıklamak için, Watson ve Crick ve daha önce Pauling, zincirin bir spiral şeklinde büküldüğünü (veya daha doğrusu bir sarmal oluşturduğunu varsaydılar, çünkü kelimenin tam anlamıyla spiral, dönüşler uzayda silindirik bir yüzeyden ziyade konik bir yüzey oluşturduğunda elde edilir). Daha sonra 3.4 nm'lik periyot, bu spiralin ardışık dönüşleri arasındaki mesafeye karşılık gelecektir. Böyle bir spiral çok yoğun veya biraz gergin olabilir, yani dönüşleri düz veya dik olabilir. 3.4 nm'lik periyot, ardışık nükleotitler (0.34 nm) arasındaki mesafenin tam olarak 10 katı olduğundan, sarmalın her tam dönüşünün 10 nükleotit içerdiği açıktır. Bu verilerden Watson ve Crick, dönüşler arasındaki mesafe 3.4 nm'ye eşit olan, 2 nm çapında bir sarmal şeklinde bükülmüş bir polinükleotit zincirinin yoğunluğunu hesaplayabildiler. Böyle bir ipliğin, zaten bilinen DNA'nın gerçek yoğunluğunun yarısı kadar bir yoğunluğa sahip olacağı ortaya çıktı. DNA molekülünün iki zincirden oluştuğunu, bunun bir çift nükleotid sarmalı olduğunu varsaymak zorunda kaldım.
Bir sonraki görev, elbette, çift sarmalı oluşturan iki iplik arasındaki uzamsal ilişkiyi aydınlatmaktı. Watson ve Crick, fiziksel modellerinde bir dizi dizi düzenlemesini denedikten sonra, mevcut tüm veriler için en uygun olanın, iki polinükleotid sarmalının zıt yönlerde ilerlediği bir model olduğunu buldular; bu durumda, şeker ve fosfat kalıntılarından oluşan zincirler bir çift sarmalın yüzeyini oluşturur ve içinde pürinler ve pirimidinler bulunur. Karşılıklı iki zincire ait olan bazlar, hidrojen bağları ile çiftler halinde bağlanır; zincirleri bir arada tutan, böylece molekülün genel konfigürasyonunu sabitleyen bu hidrojen bağlarıdır.
DNA çift sarmalı, basamakları yatay kalan sarmal bir halat merdiveni olarak düşünülebilir. Daha sonra iki uzunlamasına ip, şeker ve fosfat kalıntılarının zincirlerine karşılık gelecek ve çapraz çubuklar, hidrojen bağlarıyla bağlanmış azotlu baz çiftlerine karşılık gelecek.
Olası modeller üzerinde daha fazla çalışmanın bir sonucu olarak, Watson ve Crick, her "enine çubuğun" bir pürin ve bir pirimidin içermesi gerektiği sonucuna vardılar; 2 nm'lik bir periyotta (çift sarmalın çapına tekabül eder), iki pürin için yeterli alan olmazdı ve iki pirimidin, uygun hidrojen bağları oluşturmak için birbirine yeterince yakın olamazdı. Ayrıntılı modelin derinlemesine incelenmesi, doğru boyutta bir kombinasyon oluşturan adenin ve sitozinin, aralarında hidrojen bağları oluşturacak şekilde düzenlenemediğini gösterdi. Benzer raporlar, guanin-timin kombinasyonunu da dışlanmaya zorlarken, adenin-timin ve guanin-sitozin kombinasyonlarının oldukça kabul edilebilir olduğu bulundu. Hidrojen bağlarının doğası öyledir ki, adenin timin ile ve guanin sitozin ile çiftleşir. Bu spesifik baz eşleşmesi kavramı, herhangi bir DNA molekülünde adenin miktarının her zaman timin içeriğine eşit olduğu ve guanin miktarının her zaman sitozin miktarına eşit olduğu "Chargaff kuralı"nı açıklamayı mümkün kılmıştır. . Adenin ile timin arasında iki, guanin ile sitozin arasında üç hidrojen bağı oluşur. Bir zincirdeki her bir adenin karşısında hidrojen bağı oluşumundaki bu özellik nedeniyle, diğerinde timin bulunur; aynı şekilde, her guanine karşı sadece sitozin yerleştirilebilir. Böylece zincirler birbirini tamamlayıcıdır, yani bir zincirdeki nükleotit dizisi, diğerindeki diziyi benzersiz şekilde belirler. İki zincir zıt yönlerde ilerler ve bunların fosfat uç grupları çift sarmalın zıt uçlarındadır.
Araştırmalarının bir sonucu olarak, 1953'te Watson ve Crick, DNA molekülünün yapısı için günümüze uygun kalan bir model önerdi (Şekil 3). Modele göre, bir DNA molekülü iki tamamlayıcı polinükleotit zincirinden oluşur. Her DNA zinciri, on binlerce nükleotitten oluşan bir polinükleotittir. İçinde, komşu nükleotitler, güçlü bir kovalent bağ ile bir fosforik asit kalıntısı ve deoksiribozun kombinasyonu nedeniyle düzenli bir pentoz-fosfat omurgası oluşturur. Bir polinükleotit zincirinin azotlu bazları, diğerinin azotlu bazlarına karşı kesin olarak tanımlanmış bir sırada düzenlenir. Polinükleotid zincirindeki azotlu bazların değişimi düzensizdir.
DNA zincirindeki azotlu bazların düzenlenmesi tamamlayıcıdır (Yunanca "tamamlayıcı" - eklemeden), yani. adenin (A)'ya karşı her zaman timindir (T) ve guanine (G) karşı - sadece sitozin (C). Bu, A ve T'nin yanı sıra G ve C'nin kesinlikle birbirine karşılık geldiği gerçeğiyle açıklanır, yani. birbirini tamamlar. Bu yazışma, bir çift pürin ve pirimidin içinde hidrojen bağlarının oluşumuna izin veren bazların kimyasal yapısı tarafından verilir. A ve T arasında iki, G ve C arasında üç bağ vardır. Bu bağlar, DNA molekülünün uzayda kısmi stabilizasyonunu sağlar. Çift sarmalın kararlılığı, A=T bağlarından daha kararlı olan G≡C bağlarının sayısı ile doğru orantılıdır.
DNA'nın bir dizisindeki bilinen nükleotid dizisi, tamamlayıcılık ilkesiyle başka bir zincirin nükleotidlerinin oluşturulmasını mümkün kılar.
Ayrıca aromatik yapıya sahip azotlu bazların, sulu çözelti deyim yerindeyse bir madeni para yığını oluşturacak şekilde üst üste dizilirler. Bu yığın oluşturma işlemi organik moleküller istifleme denir. Söz konusu Watson-Crick modelinin DNA molekülünün polinükleotit zincirleri, benzer bir fizikokimyasal duruma sahiptir, azotlu bazları, van der Waals etkileşimlerinin (istifleme etkileşimlerinin) meydana geldiği düzlemler arasında bir madeni para yığını şeklinde düzenlenir.
Tamamlayıcı bazlar arasındaki (yatay olarak) hidrojen bağları ve van der Waals kuvvetleri nedeniyle (dikey olarak) bir polinükleotit zincirindeki baz düzlemleri arasındaki istifleme etkileşimi, DNA molekülüne uzayda ek stabilizasyon sağlar.
Her iki zincirin şeker-fosfat omurgaları dışa dönük ve bazlar içe, birbirine doğru. DNA'daki ipliklerin yönü antiparaleldir (bir tanesi 5"->3" yönüne sahiptir, diğeri - 3"->5" yönüne sahiptir, yani bir ipliğin 3"-ucu 5"-ucunun karşısında yer alır. diğerinin.). Zincirler, ortak bir eksene sahip dik sarmallar oluşturur. Sarmalın bir dönüşü 10 nükleotittir, dönüşün boyutu 3.4 nm, her nükleotidin yüksekliği 0.34 nm, sarmalın çapı 2.0 nm'dir. Bir ipliğin diğeri etrafında dönmesinin bir sonucu olarak, DNA çift sarmalında bir büyük oluk (yaklaşık 20 Å çapında) ve bir küçük oluk (yaklaşık 12 Å) oluşur. Watson-Crick çift sarmalının bu formu daha sonra B formu olarak adlandırıldı. Hücrelerde, DNA genellikle en kararlı olan B formunda bulunur.
DNA'nın İşlevleri
Önerilen model, genetik bilginin depolanması ve 4 nükleotidin çok çeşitli ardışık kombinasyonları tarafından sağlanan genlerin çeşitliliği dahil olmak üzere deoksiribonükleik asidin biyolojik özelliklerinin çoğunu ve bir genetik kodun varlığı gerçeğini açıkladı. replikasyon süreci tarafından sağlanan genetik bilgiyi kendi kendine çoğaltır ve iletir ve genetik bilginin proteinler şeklinde uygulanmasının yanı sıra enzim proteinlerinin yardımıyla oluşturulan diğer bileşikler.
DNA'nın temel işlevleri.
- DNA, genetik kodun varlığı gerçeğiyle sağlanan genetik bilginin taşıyıcısıdır.
- Hücre ve organizmaların nesillerinde üreme ve aktarılan genetik bilgi. Bu işlev, çoğaltma işlemi tarafından sağlanır.
- Genetik bilginin proteinler şeklinde uygulanmasının yanı sıra enzim proteinlerinin yardımıyla oluşturulan diğer bileşikler. Bu işlev, transkripsiyon ve çeviri süreçleri tarafından sağlanır.

Çift sarmallı DNA'nın organizasyon biçimleri
DNA birkaç tipte çift sarmal oluşturabilir (Şekil 4). Halihazırda altı form zaten bilinmektedir (A'dan E'ye ve Z formuna).
Rosalind Franklin tarafından kurulan DNA'nın yapısal formları, nükleik asit molekülünün suyla doygunluğuna bağlıdır. X-ışını kırınım analizini kullanan DNA lifleri üzerinde yapılan çalışmalarda, X-ışını kırınım modelinin radikal olarak bu lifin hangi bağıl neme, bu lifin ne kadar su doygunluğuna sahip olduğuna bağlı olduğu gösterilmiştir. Elyaf yeterince su ile doyurulmuşsa, bir radyografi elde edildi. Kurutulduğunda, yüksek nemli bir fiberin X-ışını modelinden çok farklı, tamamen farklı bir X-ışını deseni ortaya çıktı.
Yüksek nemli DNA molekülüne B-şekli denir. Fizyolojik koşullar altında (düşük tuz konsantrasyonu, yüksek derecede hidrasyon), baskın yapısal DNA tipi B-formudur (çift sarmallı DNA'nın ana formu Watson-Crick modelidir). Böyle bir molekülün sarmal aralığı 3.4 nm'dir. Azotlu bazlar - bükülmüş "madeni para" yığınları şeklinde dönüş başına 10 tamamlayıcı çift vardır. Yığınlar, yığınların iki zıt "madeni parası" arasındaki hidrojen bağları ile bir arada tutulur ve fosfodiester omurgasının iki şeridi ile sağ-elli bir sarmal şeklinde bükülerek "sarılır". Azotlu bazların düzlemleri sarmalın eksenine diktir. Komşu tamamlayıcı çiftler birbirlerine göre 36° döndürülür. Sarmal çapı 20Â'dır, purin nükleotidi 12Â'yı ve pirimidin nükleotidi 8Â'yı işgal eder.
Düşük nemli DNA molekülüne A-formu denir. A-formu, daha az yüksek hidrasyon koşulları altında ve daha yüksek Na + veya K + iyonları içeriğinde oluşturulur. Bu daha geniş sağ elini kullanan konformasyon dönüş başına 11 baz çiftine sahiptir. Azotlu bazların düzlemleri, sarmalın eksenine daha güçlü bir eğime sahiptir, normalden sarmalın eksenine 20 ° saparlar. Bu, 5 Å çapında bir iç boşluğun varlığına işaret eder. Bitişik nükleotitler arasındaki mesafe 0.23 nm, bobinin uzunluğu 2.5 nm ve sarmalın çapı 2.3 nm'dir.
Başlangıçta, DNA'nın A formunun daha az önemli olduğu düşünülüyordu. Bununla birlikte, daha sonra DNA'nın A formunun yanı sıra B formunun biyolojik olarak büyük önemi olduğu ortaya çıktı. Şablon-tohum kompleksindeki RNA-DNA sarmalı, A-formunun yanı sıra RNA-RNA sarmalı ve RNA saç tokası yapılarına sahiptir (2'-hidroksil riboz grubu, RNA moleküllerinin B-formunu oluşturmasına izin vermez) . DNA'nın A formu sporlarda bulunur. DNA'nın A formunun UV ışınlarına B formuna göre 10 kat daha dayanıklı olduğu tespit edilmiştir.
A formu ve B formu denir kanonik formlar DNA.
Formlar C-E ayrıca sağ elini kullananlar, oluşumları yalnızca özel deneylerde gözlemlenebilir ve görünüşe göre in vivo olarak mevcut değiller. DNA'nın C-formu, B-DNA'ya benzer bir yapıya sahiptir. Tur başına baz çifti sayısı 9.33 ve sarmalın uzunluğu 3.1 nm'dir. Baz çiftleri, eksene dik konuma göre 8 derecelik bir açıyla eğimlidir. Oluklar, B-DNA'nın oluklarına yakın boyuttadır. Bu durumda, ana oluk biraz daha küçüktür ve küçük oluk daha derindir. Doğal ve sentetik DNA polinükleotitleri C-formuna geçebilir.
| Tablo 1. Bazı DNA yapılarının özellikleri | |||
| Spiral tipi | A | B | Z |
| Spiral adım | 0,32 nm | 3.38 nm | 4,46 nm |
| Spiral büküm | Doğru | Doğru | Sol |
| Tur başına baz çifti sayısı | 11 | 10 | 12 |
| Temel düzlemler arasındaki mesafe | 0,256 nm | 0,338 nm | 0.371 nm |
| Glikozidik bağ yapısı | anti | anti | anti-C syn-G |
| Furanoz halka yapısı | C3 "-endo | C2 "-endo | C3 "-endo-G C2 "-endo-C |
| Oluk genişliği, küçük/büyük | 1.11/0.22 nm | 0,57/1,17 nm | 0.2/0.88 nm |
| Oluk derinliği, küçük/büyük | 0.26/1.30 nm | 0.82/0.85 nm | 1.38/0.37 nm |
| Spiral çapı | 2,3 nm | 2.0 nm | 1.8 nm |
DNA'nın yapısal elemanları
(kanonik olmayan DNA yapıları)
DNA'nın yapısal elemanları, bazı özel dizilerle sınırlı olağandışı yapıları içerir:
|

DNA'nın Z-formu 1979'da hekzanükleotid d(CG)3 - üzerinde çalışırken keşfedildi. MIT profesörü Alexander Rich ve ekibi tarafından açılmıştır. Z-formu, oluşumunun, pürinlerin pirimidinlerle dönüşümlü olduğu DNA bölgelerinde (örneğin, 5'-HCHCHC-3') veya 5' tekrarlarında gözlemlenmesi nedeniyle DNA'nın en önemli yapısal elemanlarından biri haline gelmiştir. -CHCHCH-3' metillenmiş sitozin içerir. Z-DNA'nın oluşumu ve stabilizasyonu için temel bir koşul, içinde, anti-konformasyonda pirimidin bazları ile değişen, sen-konformasyonda purin nükleotitlerinin mevcudiyetiydi.
Doğal DNA molekülleri, (CG)n gibi diziler içermedikçe çoğunlukla doğru B formunda bulunur. Ancak, bu tür diziler DNA'nın bir parçasıysa, o zaman bu bölgeler, çözeltinin iyonik kuvveti veya fosfodiester omurgasındaki negatif yükü nötralize eden katyonlar, Z-formuna dönüşürken, zincirdeki diğer DNA bölgeleri kalır. klasik B-formunda. Böyle bir geçişin olasılığı, DNA çift sarmalındaki iki ipliğin dinamik bir durumda olduğunu ve sağ formdan sola geçerek birbirine göre gevşeyebileceğini gösterir. DNA yapısının konformasyonel dönüşümlerine izin veren bu kararsızlığın biyolojik sonuçları henüz tam olarak anlaşılamamıştır. Z-DNA bölgelerinin belirli genlerin ekspresyonunun düzenlenmesinde rol oynadığına ve genetik rekombinasyonda yer aldığına inanılmaktadır.
DNA'nın Z-formu, fosfodiester omurgasının molekül ekseni boyunca zikzak olduğu, solak bir çift sarmaldır. Bu nedenle molekülün adı (zikzak) -DNA. Z-DNA, doğada bilinen en az bükülmüş (dönüş başına 12 baz çifti) ve en ince olanıdır. Bitişik nükleotitler arasındaki mesafe 0.38 nm, bobin uzunluğu 4.56 nm ve Z-DNA çapı 1.8 nm'dir. Ayrıca, görünüm Bu DNA molekülü, tek bir oluğun varlığı ile ayırt edilir.
DNA'nın Z-formu prokaryotik ve ökaryotik hücrelerde bulunmuştur. Bugüne kadar, DNA'nın Z-formu ile B-formunu ayırt edebilen antikorlar elde edilmiştir. Bu antikorlar, Drosophila (Dr. melanogaster) tükürük bezi hücrelerinin dev kromozomlarının spesifik bölgelerine bağlanır. Daha yoğun bölgelerin (diskler) daha az yoğun bölgelerle (diskler arası) kontrast oluşturduğu bu kromozomların olağandışı yapısı nedeniyle bağlanma reaksiyonunu takip etmek kolaydır. Z-DNA bölgeleri interdisklerde bulunur. Bundan, Z-formunun ayrı bölümlerinin boyutları henüz bilinmemekle birlikte, Z-formunun doğal koşullarda gerçekten var olduğu sonucu çıkar.

(değiştiriciler) - DNA'daki en ünlü ve sıklıkla meydana gelen baz dizileri. Palindrom, aynı şekilde soldan sağa ve tersi şekilde okunan bir kelime veya ifadedir. Bu tür kelime veya deyimlere örnek olarak: KÖPEK, KUZAK, SEL VE AZOR'UN PARÇALARINA DÜŞEN BİR GÜL. DNA bölümlerine uygulandığında, bu terim (palindrom), zincir boyunca sağdan sola ve soldan sağa aynı nükleotid değişimi anlamına gelir ("kulübe" kelimesindeki harfler gibi, vb.).
Bir palindrom, iki DNA zincirine göre ikinci dereceden bir simetriye sahip olan baz dizilerinin ters çevrilmiş tekrarlarının varlığı ile karakterize edilir. Bu tür diziler, bariz nedenlerden dolayı, kendi kendini tamamlayıcı niteliktedir ve saç tokası veya haç biçimli yapılar oluşturmaya eğilimlidir (Şek.). Firketeler, düzenleyici proteinlerin kromozom DNA'sının genetik metninin kopyalandığı yeri tanımasına yardımcı olur.
Aynı DNA zincirinde ters çevrilmiş bir tekrarın mevcut olduğu durumlarda, böyle bir diziye ayna tekrarı denir. Ayna tekrarları kendi kendini tamamlayıcı özelliklere sahip değildir ve bu nedenle saç tokası veya haç biçimli yapılar oluşturamazlar. Bu tip diziler neredeyse tüm büyük DNA moleküllerinde bulunur ve sadece birkaç baz çiftinden birkaç bin baz çiftine kadar değişebilir.
Ökaryotik hücrelerde haç biçimli yapılar biçiminde palindromların varlığı kanıtlanmamıştır, ancak E. coli hücrelerinde in vivo olarak çok sayıda haç biçiminde yapılar bulunmuştur. RNA'da veya tek iplikli DNA'da kendini tamamlayıcı dizilerin varlığı, nükleik zincirin çözeltilerde, birçok "saç tokası" oluşumu ile karakterize edilen belirli bir uzaysal yapıya katlanmasının ana nedenidir.

DNA'nın H-formu- bu, üç DNA zincirinden oluşan bir sarmaldır - DNA'nın üçlü sarmalı. Bu, Hoogsteen çifti olarak adlandırılan oluşumu ile büyük oluğuna uyan üçüncü tek iplikli DNA ipliği ile Watson-Crick çift sarmalının bir kompleksidir.
Böyle bir tripleksin oluşumu, DNA çift sarmalının, bölümünün yarısı çift sarmal şeklinde kalacak ve ikinci yarısı kopuk olacak şekilde eklenmesi sonucu meydana gelir. Bu durumda, bağlantısız spirallerden biri, çift sarmalın ilk yarısı - üçlü sarmal ile yeni bir yapı oluşturur ve ikincisi, tek filamentli bir bölüm şeklinde yapılandırılmamış olarak ortaya çıkar. Bu yapısal geçişin bir özelliği, protonları yeni yapıyı stabilize eden ortamın pH'ına keskin bir bağımlılıktır. Bu özellik nedeniyle, oluşumu ayna tekrarı olan homopurin-homopirimidin bölgelerini içeren süper sarmal plazmitlerde bulunan yeni yapıya DNA'nın H-formu adı verildi.
Daha sonraki çalışmalarda, bazı homopurin-homopirimidin çift sarmallı polinükleotitlerin yapısal geçiş olasılığı, aşağıdakileri içeren üç sarmallı bir yapının oluşumuyla belirlenmiştir:
- bir homopurin ve iki homopirimidin zinciri ( Py-Pu-Py tripleks) [Hoogsteen etkileşimi].
Py-Pu-Py tripleksinin kurucu blokları, kanonik izomorfik CGC+ ve TAT triadlarıdır. Tripleksin stabilizasyonu CGC+ triadının protonlanmasını gerektirir, dolayısıyla bu tripleksler çözeltinin pH'ına bağlıdır.
- bir homopirimidin ve iki homopurin zinciri ( Py-Pu-Pu tripleks) [ters Hoogsteen etkileşimi].
Py-Pu-Pu tripleksinin kurucu blokları, kanonik izomorfik CGG ve TAA üçlüleridir. Py-Pu-Pu triplekslerinin temel bir özelliği, kararlılıklarının çift yüklü iyonların varlığına bağlı olmasıdır ve farklı dizilerdeki tripleksleri stabilize etmek için farklı iyonlara ihtiyaç vardır. Py-Pu-Pu triplekslerinin oluşumu, kendilerini oluşturan nükleotidlerin protonlanmasını gerektirmediğinden, bu tür tripleksler nötr pH'ta var olabilir.
Not: Doğrudan ve ters Hoogsteen etkileşimi, 1-metiltiminin simetrisi ile açıklanır: 180 ° döndürme, hidrojen bağları sistemi korunurken O4 atomunun yerinin O2 atomu tarafından işgal edilmesine yol açar.
İki tür üçlü sarmal vardır:
- üçüncü ipliğin polaritesinin Watson-Crick dupleksinin homopurin zincirinin polaritesi ile aynı olduğu paralel üçlü sarmallar
- üçüncü ve homopurin zincirlerinin kutuplarının zıt olduğu antiparalel üçlü sarmallar.

G-dörtlü- 4 iplikli DNA. Böyle bir yapı, sözde G-dörtlü - dört guaninin yuvarlak bir dansını oluşturan dört guanin varsa oluşur.
Bu tür yapıların oluşma olasılığının ilk ipuçları, Watson ve Crick'in çığır açan çalışmasından çok önce - 1910 kadar erken bir tarihte elde edildi. Daha sonra Alman kimyager Ivar Bang, DNA'nın bileşenlerinden birinin - guanosik asit - yüksek konsantrasyonlarda jeller oluşturduğunu, DNA'nın diğer bileşenlerinin ise bu özelliğe sahip olmadığını keşfetti.
1962'de X-ışını kırınım yöntemi kullanılarak bu jelin hücre yapısını oluşturmak mümkün oldu. Birbirini bir daire içinde birbirine bağlayan ve karakteristik bir kare oluşturan dört guanin kalıntısından oluştuğu ortaya çıktı. Merkezde bağ bir metal iyonu (Na, K, Mg) tarafından desteklenir. Aynı yapılar çok fazla guanin içeriyorsa DNA'da da oluşturulabilir. Bu düz kareler (G-dörtlüleri) oldukça kararlı, yoğun yapılar (G-dörtlüleri) oluşturmak için istiflenir.
Dört ayrı DNA dizisi, dört zincirli kompleksler halinde dokunabilir, ancak bu bir istisnadır. Daha sıklıkla, tek bir nükleik asit dizisi, karakteristik kalınlaşmalar (örneğin, kromozomların uçlarında) oluşturan bir düğüme basitçe bağlanır veya çift sarmallı DNA, bazı guanin bakımından zengin bölgelerde yerel bir dörtlü oluşturur.
En çok çalışılan, kromozomların uçlarında - telomerlerde ve onkopromoterlerde - dörtlülerin varlığıdır. Bununla birlikte, bu tür DNA'nın insan kromozomlarındaki lokalizasyonunun tam olarak anlaşılması hala bilinmemektedir.
Doğrusal formdaki tüm bu olağandışı DNA yapıları, DNA'nın B formuna kıyasla kararsızdır. Bununla birlikte, DNA, süper sarmal olarak bilinen şeye sahip olduğunda, genellikle bir halka topolojik gerilim biçiminde bulunur. Bu koşullar altında, kanonik olmayan DNA yapıları kolaylıkla oluşturulur: Z-formları, "haçlar" ve "firketeler", H-formları, guanin dörtlüleri ve i-motifi.
- Süper sarmal form - hücre çekirdeğinden pentoz-fosfat omurgasına zarar vermeden serbest bırakıldığında not edilir. Süper bükümlü kapalı halkalar şeklindedir. Süper bükülmüş durumda, DNA çift sarmalı en az bir kez "kendi üzerine bükülür", yani. en az bir süper bobin içerir (sekiz rakamı şeklini alır).
- Gevşemiş DNA durumu - tek bir kopma ile gözlemlenir (bir iplik kopması). Bu durumda süper bobinler kaybolur ve DNA kapalı bir halka şeklini alır.
- DNA'nın lineer formu, çift sarmalın iki ipliği kırıldığında gözlemlenir.
DNA'nın üçüncül yapısı
DNA'nın üçüncül yapısıçift sarmallı bir molekülün uzayda ek bükülmesinin bir sonucu olarak oluşur - onun süper sarılması. Ökaryotik hücrelerde DNA molekülünün prokaryotların aksine süper sarmalanması, proteinlerle kompleksler şeklinde gerçekleştirilir.
Hemen hemen tüm ökaryotik DNA, çekirdeğin kromozomlarında bulunur, mitokondride, bitkilerde ve plastidlerde sadece küçük bir miktar bulunur. Ökaryotik hücrelerin kromozomlarının (insan kromozomları dahil) ana maddesi, çift sarmallı DNA, histon ve histon olmayan proteinlerden oluşan kromatindir.
Kromatinin histon proteinleri
Histonlar, kromatinin %50'sini oluşturan basit proteinlerdir. Hayvanların ve bitkilerin incelenen tüm hücrelerinde, beş ana histon sınıfı bulundu: H1, H2A, H2B, H3, H4, büyüklük, amino asit bileşimi ve yük bakımından farklılık gösterir (her zaman pozitif).
Memeli histon H1, yaklaşık 215 amino asit içeren tek bir polipeptit zincirinden oluşur; diğer histonların boyutları 100 ila 135 amino asit arasında değişir. Hepsi spiralize edilmiş ve yaklaşık 2.5 nm çapında bir küre şeklinde bükülmüştür, alışılmadık derecede büyük miktarda pozitif yüklü amino asitler lizin ve arginin içerir. Histonlar asetillenmiş, metillenmiş, fosforile edilmiş, poli(ADP)-ribosilatlanmış olabilir ve H2A ve H2B histonları ubikuitine kovalent olarak bağlanabilir. Histonların yapı oluşumunda ve işlevlerin performansında bu tür değişikliklerin rolü henüz tam olarak açıklanmamıştır. Bunun, DNA ile etkileşime girme ve genlerin hareketini düzenleyen mekanizmalardan birini sağlama yetenekleri olduğu varsayılmaktadır.
Histonlar, esas olarak DNA'nın negatif yüklü fosfat grupları ile pozitif yüklü lizin ve histonların arginin kalıntıları arasında oluşan iyonik bağlar (tuz köprüleri) yoluyla DNA ile etkileşime girer.
Kromatinin histon olmayan proteinleri
Histon olmayan proteinler, histonlardan farklı olarak çok çeşitlidir. DNA'ya bağlanan histon olmayan proteinlerin 590'a kadar farklı fraksiyonu izole edilmiştir. Asidik proteinler olarak da adlandırılırlar, çünkü yapılarında asidik amino asitler baskındır (bunlar polianyonlardır). Kromatin aktivitesinin spesifik düzenlenmesi, çeşitli histon olmayan proteinlerle ilişkilidir. Örneğin, DNA replikasyonu ve ekspresyonu için gerekli olan enzimler, geçici olarak kromatine bağlanabilir. Diğer proteinler, örneğin çeşitli düzenleyici işlemlerde yer alanlar, DNA'ya yalnızca belirli dokularda veya farklılaşmanın belirli aşamalarında bağlanır. Her protein, belirli bir DNA nükleotid dizisine (DNA bölgesi) tamamlayıcıdır. Bu grup şunları içerir:
- bölgeye özgü çinko parmak proteinleri ailesi. Her "çinko parmak", 5 nükleotid çiftinden oluşan belirli bir bölgeyi tanır.
- siteye özgü proteinlerin bir ailesi - homodimerler. DNA ile temas halinde olan böyle bir proteinin bir parçası, bir "sarmal dönüşlü sarmal" yapıya sahiptir.
- yüksek hareketli proteinler (HMG proteinleri - İngilizce'den, yüksek hareketli jel proteinlerinden), kromatin ile sürekli olarak ilişkili bir grup yapısal ve düzenleyici proteindir. 30 kD'den daha düşük bir moleküler ağırlığa sahiptirler ve yüksek miktarda yüklü amino asit içeriği ile karakterize edilirler. Düşük moleküler ağırlıklarından dolayı HMG proteinleri poliakrilamid jel elektroforezi sırasında oldukça hareketlidir.
- replikasyon, transkripsiyon ve onarım enzimleri.
DNA ve RNA sentezinde yer alan yapısal, düzenleyici proteinler ve enzimlerin katılımıyla, nükleozom ipliği oldukça yoğun bir protein ve DNA kompleksine dönüştürülür. Ortaya çıkan yapı, orijinal DNA molekülünden 10.000 kat daha kısadır.

kromatin
Kromatin, nükleer DNA içeren bir protein kompleksidir ve inorganik maddeler. Kromatinin çoğu inaktiftir. Yoğun şekilde paketlenmiş, yoğunlaştırılmış DNA içerir. Bu heterokromatindir. İfade edilmeyen bölgelerden oluşan kurucu, genetik olarak aktif olmayan kromatin (uydu DNA) ve fakültatif - birkaç nesilde aktif olmayan, ancak belirli koşullar altında ifade edebilen vardır.
Aktif kromatin (ökromatin) yoğunlaşmaz, yani. daha az sıkı paketlenmiş. Farklı hücrelerde içeriği %2 ile %11 arasında değişmektedir. Beyin hücrelerinde en çok - %10-11, karaciğer hücrelerinde - 3-4 ve böbreklerde - %2-3. Ökromatinin aktif bir transkripsiyonu vardır. Aynı zamanda, yapısal organizasyonu, belirli bir organizma tipinde bulunan aynı DNA genetik bilgisini, özel hücrelerde farklı şekillerde kullanmayı mümkün kılar.
Bir elektron mikroskobunda, kromatin görüntüsü boncuklara benzer: ipliksi köprülerle ayrılmış, yaklaşık 10 nm boyutunda küresel kalınlaşmalar. Bu küresel kalınlaşmalara nükleozom denir. Nükleozom, kromatinin yapısal birimidir. Her nükleozom, nükleozom çekirdeği başına 1.75 sola dönüş oluşturmak üzere sarılmış 146 bp uzunluğunda süper sarmallı bir DNA segmenti içerir. Nükleozomal çekirdek, 11 nm çapında ve 5,7 nm kalınlığında bir disk gibi görünen, her türden iki molekül olan H2A, H2B, H3 ve H4 histonlarından oluşan bir histon oktameridir (Şekil 9). Beşinci histon H1, nükleozomal çekirdeğin bir parçası değildir ve DNA'nın histon oktamerinin etrafına sarılması sürecinde yer almaz. Çift sarmalın nükleozomal çekirdeğe girip çıktığı noktalarda DNA ile temas eder. Bunlar, uzunluğu hücre tipine bağlı olarak 40 ila 50 nükleotid çifti arasında değişen DNA'nın intercore (bağlayıcı) bölümleridir. Sonuç olarak, nükleozomların parçası olan DNA parçasının uzunluğu da değişir (186 ila 196 nükleotid çifti).
Nükleozom, DNA'nın yaklaşık %90'ını içerir, geri kalanı bağlayıcıdır. Bağlayıcı aktifken nükleozomların "sessiz" kromatin parçaları olduğuna inanılmaktadır. Bununla birlikte, nükleozomlar açılabilir ve lineer hale gelebilir. Katlanmamış nükleozomlar zaten aktif kromatindir. Bu, işlevin yapıya bağımlılığını açıkça göstermektedir. Küresel nükleozomların bileşiminde ne kadar fazla kromatin varsa, o kadar az aktif olduğu varsayılabilir. Açıkçası, farklı hücrelerde, dinlenme kromatinin eşit olmayan oranı, bu tür nükleozomların sayısı ile ilişkilidir.
Elektron mikroskobik fotoğraflarda, izolasyon koşullarına ve gerilme derecesine bağlı olarak, kromatin sadece kalınlaşmaları olan uzun bir iplik - nükleozomların "boncukları" olarak değil, aynı zamanda çapı olan daha kısa ve daha yoğun bir fibril (lif) olarak da görünebilir. Oluşumu, DNA'nın bağlayıcı bölgesi ve histon H3 ile ilişkili histon H1'in etkileşimi sırasında gözlenen 30 nm, 30 nm çapında bir solenoid oluşumu ile dönüş başına altı nükleozom sarmalının ek bükülmesine yol açar. . Bu durumda, histon proteini bir dizi genin transkripsiyonuna müdahale edebilir ve böylece onların aktivitelerini düzenleyebilir.
DNA'nın yukarıda açıklanan histonlarla etkileşimleri sonucunda, ortalama çapı 2 nm ve uzunluğu 57 nm olan 186 baz çiftinden oluşan DNA çift sarmalının bir parçası, 10 nm çapında ve uzunlukta bir sarmal haline gelir. 5 nm'lik Bu sarmalın 30 nm çapında bir fibere daha sonra sıkıştırılmasıyla, yoğunlaşma derecesi altı kat daha artar.
Sonuç olarak, DNA dupleksinin beş histonla paketlenmesi 50 kat DNA yoğunlaşması ile sonuçlanır. Bununla birlikte, bu kadar yüksek derecede bir yoğunlaşma bile, metafaz kromozomundaki neredeyse 50.000-100.000 kat DNA sıkışmasını açıklayamaz. Ne yazık ki, kromatinin metafaz kromozomuna kadar paketlenmesinin detayları henüz bilinmemektedir; bu nedenle, bu işlemin sadece genel özellikleri dikkate alınabilir.
Kromozomlardaki DNA sıkıştırma seviyeleri
Her DNA molekülü ayrı bir kromozomda paketlenir. Diploid insan hücreleri, hücre çekirdeğinde bulunan 46 kromozom içerir. Bir hücrenin tüm kromozomlarının DNA'sının toplam uzunluğu 1.74 m'dir, ancak kromozomların paketlendiği çekirdeğin çapı milyonlarca kez daha küçüktür. Hücre çekirdeğindeki kromozomlarda ve kromozomlarda DNA'nın bu şekilde kompakt bir şekilde paketlenmesi, DNA ile belirli bir sırayla etkileşime giren çeşitli histon ve histon olmayan proteinler tarafından sağlanır (yukarıya bakın). DNA'nın kromozomlarda sıkıştırılması, doğrusal boyutlarını yaklaşık 10.000 kat - şartlı olarak 5 cm'den 5 mikrona düşürmeyi mümkün kılar. Birkaç sıkıştırma seviyesi vardır (Şekil 10).


- DNA çift sarmalı, 2 nm çapında ve birkaç cm uzunluğunda negatif yüklü bir moleküldür.
- nükleozomal seviye- kromatin bir elektron mikroskobunda "boncuklar" - nükleozomlar - "iplik üzerinde" bir zincir olarak görünür. Nükleozom, hem ökromatinde hem de heterokromatinde, fazlar arası çekirdekte ve metafaz kromozomlarında bulunan evrensel bir yapısal birimdir.
Nükleozomal sıkıştırma seviyesi, özel proteinler - histonlar tarafından sağlanır. Sekiz pozitif yüklü histon alanı, negatif yüklü DNA molekülünün etrafına sarıldığı nükleozomun çekirdeğini (çekirdeğini) oluşturur. Bu, 7 kat kısalma sağlarken, çap 2 nm'den 11 nm'ye çıkar.
- solenoid seviyesi
Solenoid kromozom organizasyonu seviyesi, nükleozomal filamentin bükülmesi ve ondan 20-35 nm çapında daha kalın fibrillerin oluşumu - solenoidler veya süperbitler ile karakterize edilir. Solenoid aralığı 11 nm'dir ve dönüş başına yaklaşık 6-10 nükleozom vardır. 20-35 nm çapında bir kromatin fibrilinin, her biri sekiz nükleozomdan oluşan bir granül zinciri veya süperbitler olduğuna göre, solenoid paketlemenin süperbit paketlemeden daha olası olduğu kabul edilir. Solenoid seviyesinde, DNA'nın lineer boyutu 6-10 kat küçülür, çap 30 nm'ye çıkar.
- döngü seviyesi
Döngü seviyesi, yaklaşık 30-300 kb'lik halkalar oluşturan, spesifik DNA dizilerini tanıyan ve bunlara bağlanan histon olmayan bölgeye özgü DNA bağlayıcı proteinler tarafından sağlanır. Döngü, gen ifadesini sağlar, yani. döngü sadece yapısal değil, aynı zamanda işlevsel bir oluşumdur. Bu seviyede kısalma 20-30 kez gerçekleşir. Çap 300 nm'ye çıkar. Amfibi oositlerinde ilmek benzeri "lamba fırçası" yapıları sitolojik preparasyonlarda görülebilir. Bu halkalar süper sarmal gibi görünmektedir ve muhtemelen kromatin transkripsiyon ve replikasyon birimlerine karşılık gelen DNA alanlarını temsil etmektedir. Spesifik proteinler, ilmeklerin tabanlarını ve muhtemelen bazı iç bölgelerini sabitler. Döngü benzeri alan organizasyonu, metafaz kromozomlarındaki kromatinin daha yüksek dereceli sarmal yapılara katlanmasını kolaylaştırır.
- etki alanı düzeyi
Kromozom organizasyonunun etki alanı seviyesi yeterince çalışılmamıştır. Bu seviyede, ilmek alanlarının oluşumu not edilir - %60 protein, %35 DNA ve %5 RNA içeren 25-30 nm kalınlığındaki filamentlerin (fibriller) yapıları, hücre döngüsünün tüm aşamalarında pratik olarak görünmezdir. mitoz dışındadır ve hücre çekirdeği üzerinde bir şekilde rastgele dağılmıştır. Amfibi oositlerinde ilmek benzeri "lamba fırçası" yapıları sitolojik preparasyonlarda görülebilir.
Döngü alanları bazlarıyla birlikte yerleşik bağlanma bölgeleri olarak adlandırılan ve genellikle MAR / SAR dizileri olarak adlandırılan intranükleer protein matrisine eklenir (MAR, İngiliz matrisi ile ilişkili bölgeden; SAR, İngiliz iskele bağlanma bölgelerinden) - DNA fragmanları, yüksek bir A/T baz çifti içeriği (>%65) ile karakterize edilen birkaç yüz uzun baz çifti. Her etki alanı, tek bir çoğaltma kaynağına sahip görünüyor ve özerk bir süper sarmal birim olarak işlev görüyor. Herhangi bir döngü alanı, işleyişi muhtemelen koordine edilecek birçok transkripsiyon birimi içerir - tüm alan ya aktif ya da aktif olmayan durumdadır.
Alan düzeyinde, kromatinin sıralı paketlenmesinin bir sonucu olarak, DNA'nın doğrusal boyutları yaklaşık 200 kat (700 nm) azalır.
- kromozom seviyesi
Kromozom düzeyinde, profaz kromozomu, histon olmayan proteinlerin eksenel çerçevesi etrafında ilmek alanlarının sıkıştırılmasıyla bir metafazda yoğunlaşır. Bu aşırı sargıya, hücredeki tüm H1 moleküllerinin fosforilasyonu eşlik eder. Sonuç olarak, metafaz kromozomu, sıkı bir spirale sarılmış yoğun şekilde paketlenmiş solenoid halkalar olarak tasvir edilebilir. Tipik bir insan kromozomu 2600 ilmek içerebilir. Böyle bir yapının kalınlığı 1400 nm'ye (iki kromatit) ulaşırken, DNA molekülü 104 kat kısaltılır, yani. 5 cm gerilmiş DNA'dan 5 µm'ye.
Kromozomların işlevleri
Ekstrakromozomal mekanizmalarla etkileşim içinde kromozomlar,
- kalıtsal bilgilerin saklanması
- hücresel organizasyonu oluşturmak ve sürdürmek için bu bilgileri kullanmak
- kalıtsal bilgileri okuma düzenlemesi
- genetik materyalin kendi kendini kopyalaması
- genetik materyalin bir ana hücreden yavru hücrelere aktarılması.
Bir kromatin bölgesinin aktivasyonu üzerine, yani. transkripsiyon sırasında, histon H1, önce ondan geri dönüşümlü olarak ve ardından histon oktetinden çıkarılır. Bu, kromatinin yoğunlaşmasına, 30 nm'lik bir kromatin fibrilinin 10 nm'lik bir filamente ardışık geçişine ve bunun daha sonra serbest DNA bölgelerine açılmasına, yani. nükleozomal yapının kaybı.
Hepimiz bir kişinin görünümünün, bazı alışkanlıkların ve hatta hastalıkların kalıtsal olduğunu biliyoruz. Bir canlıya ilişkin tüm bu bilgiler genlerde kodlanmıştır. Peki bu kötü şöhretli genler neye benziyor, nasıl işlev görüyor ve nerede bulunuyorlar?
Yani herhangi bir insan veya hayvanın tüm genlerinin taşıyıcısı DNA'dır. Bu bileşik 1869'da Johann Friedrich Miescher tarafından keşfedilmiştir.Kimyasal olarak DNA, deoksiribonükleik asittir. Ne anlama geliyor? Bu asit gezegenimizdeki tüm yaşamın genetik kodunu nasıl taşıyor?
DNA'nın bulunduğu yere bakarak başlayalım. İnsan hücresinde çeşitli işlevleri yerine getiren birçok organel vardır. DNA çekirdekte bulunur. Çekirdek, tüm genetik materyali - DNA'yı depolayan özel bir zarla çevrili küçük bir organeldir.
DNA molekülünün yapısı nedir?
Önce DNA'nın ne olduğuna bakalım. DNA, yapısal elementlerden - nükleotidlerden oluşan çok uzun bir moleküldür. 4 tip nükleotid vardır - adenin (A), timin (T), guanin (G) ve sitozin (C). Nükleotid zinciri şematik olarak şöyle görünür: GGAATTSTAAG.... Bu nükleotid dizisi DNA zinciridir.DNA'nın yapısı ilk olarak 1953'te James Watson ve Francis Crick tarafından deşifre edildi.
Bir DNA molekülünde, birbiri etrafında sarmal olarak bükülmüş iki nükleotit zinciri vardır. Bu nükleotid zincirleri nasıl birbirine yapışır ve bir spiral halinde bükülür? Bu fenomen, tamamlayıcılık özelliğinden kaynaklanmaktadır. Tamamlayıcılık, yalnızca belirli nükleotidlerin (tamamlayıcı) iki zincirde birbirinin karşısında olabileceği anlamına gelir. Yani, karşı adenin her zaman timindir ve zıt guanin her zaman sadece sitozindir. Bu nedenle, guanin sitozin ile ve adenin timin ile tamamlayıcıdır.Farklı zincirlerde birbirine zıt olan bu tür nükleotit çiftlerine tamamlayıcı olarak da adlandırılır.
 Şematik olarak aşağıdaki gibi temsil edilebilir:
Şematik olarak aşağıdaki gibi temsil edilebilir:
G - C
T - A
T - A
C - G
Bu tamamlayıcı çiftler A - T ve G - C formu Kimyasal bağçiftin nükleotitleri arasında ve G ve C arasındaki bağ A ve T arasındakinden daha güçlüdür. Bağ, kesinlikle tamamlayıcı bazlar arasında oluşur, yani tamamlayıcı olmayan G ve A arasında bir bağ oluşumu imkansızdır.
DNA'nın "paketlenmesi", bir DNA dizisi nasıl bir kromozom haline gelir?
DNA'nın bu nükleotid zincirleri neden birbirinin etrafında dönüyor? Bu neden gerekli? Gerçek şu ki, nükleotidlerin sayısı çok büyük ve bu kadar uzun zincirleri barındırmak için çok fazla alana ihtiyacınız var. Bu nedenle, iki DNA zincirinin diğerinin etrafında spiral bir bükülmesi vardır. Bu olgu spiralizasyon denir. Spiralizasyon sonucunda DNA zincirleri 5-6 kat kısalır.Bazı DNA molekülleri vücut tarafından aktif olarak kullanılırken diğerleri nadiren kullanılır. Bu tür nadiren kullanılan DNA molekülleri, sarmallaşmaya ek olarak daha da kompakt "paketleme" geçirir. Böyle kompakt bir pakete süper sarma denir ve DNA zincirini 25-30 kat kısaltır!
DNA sarmalı nasıl paketlenir?
 Süper sarma için, bir çubuk veya iplik makarası görünümüne ve yapısına sahip olan histon proteinleri kullanılır. Spiralize DNA iplikçikleri bu "bobinler" - histon proteinleri üzerine sarılır. Bu şekilde, uzun filaman çok kompakt bir şekilde paketlenir ve çok az yer kaplar.
Süper sarma için, bir çubuk veya iplik makarası görünümüne ve yapısına sahip olan histon proteinleri kullanılır. Spiralize DNA iplikçikleri bu "bobinler" - histon proteinleri üzerine sarılır. Bu şekilde, uzun filaman çok kompakt bir şekilde paketlenir ve çok az yer kaplar. Bir veya başka bir DNA molekülünün kullanılması gerekiyorsa, "gevşeme" işlemi gerçekleşir, yani DNA ipliği "bobinden" - histon proteininden (üzerine sarılmışsa) "açılır" ve gevşer. sarmalı iki paralel zincire ayırın. Ve DNA molekülü böyle bükülmemiş bir durumda olduğunda, gerekli genetik bilgi ondan okunabilir. Üstelik, genetik bilginin okunması sadece bükülmemiş DNA ipliklerinden gerçekleşir!
Bir dizi süper sarmal kromozom denir heterokromatin ve bilgileri okumak için mevcut olan kromozomlar - ökromatin.
Gen nedir, DNA ile ilişkisi nedir?
Şimdi genlerin ne olduğuna bakalım. Vücudumuzun kan grubunu, göz rengini, saçını, derisini ve daha birçok özelliğini belirleyen genlerin olduğu bilinmektedir. Bir gen, kesin olarak tanımlanmış bir kombinasyon halinde düzenlenmiş belirli sayıda nükleotitten oluşan, kesin olarak tanımlanmış bir DNA bölümüdür. DNA'nın kesin olarak tanımlanmış bir bölümünde yer, belirli bir genin yerinin olduğu ve bu yeri değiştirmenin imkansız olduğu anlamına gelir. Böyle bir karşılaştırma yapmak uygundur: Bir kişi belirli bir sokakta, belirli bir evde ve apartman dairesinde yaşar ve bir kişi keyfi olarak başka bir eve, daireye veya başka bir sokağa taşınamaz. Bir gende belirli sayıda nükleotit olması, her genin belirli sayıda nükleotidi olduğu ve daha fazla veya daha az olamayacağı anlamına gelir. Örneğin, insülin üretimini kodlayan gen 60 baz çifti uzunluğundadır; oksitosin hormonunun üretimini kodlayan gen 370 bp'dir. Kesin bir nükleotid dizisi, her bir gen için benzersizdir ve kesin olarak tanımlanmıştır. Örneğin, AATTAATA dizisi, insülin üretimini kodlayan bir genin bir parçasıdır. İnsülin elde etmek için tam da böyle bir dizi kullanılır; örneğin adrenalin elde etmek için farklı bir nükleotit kombinasyonu kullanılır. Yalnızca belirli bir nükleotid kombinasyonunun belirli bir "ürünü" (adrenalin, insülin, vb.) kodladığını anlamak önemlidir. "Yerinde" duran belirli sayıda nükleotidin böyle benzersiz bir kombinasyonu - bu gen.
DNA zincirinde genlere ek olarak "kodlamayan diziler" de bulunur. Bu tür kodlamayan nükleotid dizileri, genlerin işleyişini düzenler, kromozom sarmallaşmasına yardımcı olur ve bir genin başlangıç ve bitiş noktalarını işaretler. Bununla birlikte, bugüne kadar, kodlamayan dizilerin çoğunun rolü belirsizliğini koruyor.
kromozom nedir? cinsiyet kromozomları
Bir bireyin genlerinin toplamına genom denir. Doğal olarak, tüm genom tek bir DNA'da paketlenemez. Genom, 46 çift DNA molekülüne bölünmüştür. Bir çift DNA molekülüne kromozom denir. Yani tam olarak bu kromozomlar bir kişinin 46 parçasına sahiptir. Her kromozom, kesin olarak tanımlanmış bir gen kümesi taşır, örneğin, 18. kromozom, göz rengini vb. kodlayan genleri içerir. Kromozomlar, uzunluk ve şekil bakımından birbirinden farklıdır. En yaygın biçimler X veya Y biçimindedir, ancak başkaları da vardır. Bir kişinin eşleştirilmiş (çiftler) olarak adlandırılan aynı şekle sahip iki kromozomu vardır. Bu tür farklılıklarla bağlantılı olarak, tüm eşleştirilmiş kromozomlar numaralandırılmıştır - 23 çift vardır. Bu, bir çift kromozom #1, çift #2, #3 vb. olduğu anlamına gelir. Belirli bir özellikten sorumlu olan her gen aynı kromozom üzerinde bulunur. Uzmanlar için modern kılavuzlarda, genin lokalizasyonu, örneğin aşağıdaki gibi belirtilebilir: kromozom 22, uzun kol.Kromozomlar arasındaki farklar nelerdir?
 Kromozomlar birbirinden başka nasıl farklıdır? Uzun kol kelimesi ne anlama gelir? X-şekilli kromozomları ele alalım.DNA ipliklerinin çaprazlanması tam olarak ortada (X) gerçekleşebilir veya merkezi olarak gerçekleşmeyebilir. DNA ipliklerinin böyle bir kesişimi merkezi olarak gerçekleşmediğinde, kesişme noktasına göre bazı uçlar daha uzun, diğerleri sırasıyla daha kısadır. Bu tür uzun uçlara genellikle kromozomun uzun kolu ve kısa uçlara sırasıyla kısa kol denir. Y şeklindeki kromozomlar çoğunlukla uzun kollar tarafından işgal edilir ve kısa olanlar çok küçüktür (şematik görüntüde bile gösterilmezler).
Kromozomlar birbirinden başka nasıl farklıdır? Uzun kol kelimesi ne anlama gelir? X-şekilli kromozomları ele alalım.DNA ipliklerinin çaprazlanması tam olarak ortada (X) gerçekleşebilir veya merkezi olarak gerçekleşmeyebilir. DNA ipliklerinin böyle bir kesişimi merkezi olarak gerçekleşmediğinde, kesişme noktasına göre bazı uçlar daha uzun, diğerleri sırasıyla daha kısadır. Bu tür uzun uçlara genellikle kromozomun uzun kolu ve kısa uçlara sırasıyla kısa kol denir. Y şeklindeki kromozomlar çoğunlukla uzun kollar tarafından işgal edilir ve kısa olanlar çok küçüktür (şematik görüntüde bile gösterilmezler). Kromozomların boyutu dalgalanır: en büyüğü 1 ve 3 numaralı çiftlerin kromozomları, en küçükleri 17 ve 19 numaralı çiftlerin kromozomlarıdır.
Şekil ve boyutlara ek olarak, kromozomlar işlevlerinde farklılık gösterir. 23 çiftin 22'si somatik, 1 çifti cinseldir. Bu ne anlama geliyor? Somatik kromozomlar, bir bireyin tüm dış belirtilerini, özelliklerini belirler. davranışsal tepkiler, kalıtsal psikotip, yani her bireyin tüm özellikleri ve özellikleri. Bir çift cinsiyet kromozomu, bir kişinin cinsiyetini belirler: erkek veya kadın. İki tür insan cinsiyet kromozomu vardır - X (X) ve Y (Y). XX (x - x) olarak birleştirilirlerse - bu bir kadın ve eğer XY (x - y) ise - önümüzde bir erkek var.
Kalıtsal hastalıklar ve kromozom hasarı
Bununla birlikte, genomun "bozulmaları" vardır, daha sonra insanlarda genetik hastalıklar tespit edilir. Örneğin 21 çift kromozomda iki yerine üç kromozom olduğunda Down sendromlu bir kişi doğar.Genetik materyalin, hastalığın başlangıcına yol açmayan, aksine, iyi özellikler veren birçok küçük "bozulması" vardır. Genetik materyalin tüm "bozulmalarına" mutasyon denir. Hastalığa veya organizmanın özelliklerinin bozulmasına neden olan mutasyonlar negatif, yeni faydalı özelliklerin oluşumuna yol açan mutasyonlar ise pozitif kabul edilir.
Ancak günümüzde insanların muzdarip olduğu hastalıkların çoğu ile ilgili olarak, kalıtsal bir hastalık değil, sadece bir yatkınlıktır. Örneğin bir çocuğun babasında şeker yavaş emilir. Bu, çocuğun diyabetli doğacağı anlamına gelmez, ancak çocuğun bir yatkınlığı olacaktır. Bu, bir çocuk tatlıları ve un ürünlerini kötüye kullanırsa, diyabet geliştireceği anlamına gelir.
Bugün sözde tahmin edici ilaç. Bu tıbbi uygulama çerçevesinde, bir kişide yatkınlıklar ortaya çıkar (karşılık gelen genlerin tanımlanmasına dayanarak) ve daha sonra ona öneriler verilir - hangi diyetin izleneceği, çalışma ve dinlenme rejimlerinin uygun şekilde nasıl değiştirileceği hasta olmak.
DNA'da kodlanmış bilgiler nasıl okunur?
Fakat DNA'da bulunan bilgiyi nasıl okuyabilirsiniz? Kendi vücudu bunu nasıl kullanıyor? DNA'nın kendisi bir tür matristir, ancak basit değil, kodlanmıştır. DNA matrisinden bilgi okumak için önce özel bir taşıyıcıya - RNA'ya aktarılır. RNA kimyasal olarak ribonükleik asittir. Nükleer zardan hücreye geçebilmesi bakımından DNA'dan farklıdır ve DNA bu yetenekten yoksundur (sadece çekirdekte olabilir). Kodlanmış bilgi hücrenin kendisinde kullanılır. Yani RNA, çekirdekten hücreye kodlanmış bilginin taşıyıcısıdır.RNA sentezi nasıl gerçekleşir, RNA yardımıyla protein nasıl sentezlenir?
 Bilginin "okunması" gereken DNA zincirleri çözülür, özel bir enzim, "oluşturucu" onlara yaklaşır ve DNA zincirine paralel olarak tamamlayıcı bir RNA zinciri sentezler. RNA molekülü ayrıca 4 tip nükleotitten oluşur - adenin (A), urasil (U), guanin (G) ve sitozin (C). Bu durumda, aşağıdaki çiftler tamamlayıcıdır: adenin - urasil, guanin - sitozin. Gördüğünüz gibi, DNA'dan farklı olarak RNA, timin yerine urasil kullanır. Yani “oluşturucu” enzim şu şekilde çalışır: DNA zincirinde A görürse RNA zincirine Y'yi, G ise C'yi bağlar vb. Böylece, transkripsiyon sırasında her aktif genden bir şablon oluşur - nükleer zardan geçebilen bir RNA kopyası.
Bilginin "okunması" gereken DNA zincirleri çözülür, özel bir enzim, "oluşturucu" onlara yaklaşır ve DNA zincirine paralel olarak tamamlayıcı bir RNA zinciri sentezler. RNA molekülü ayrıca 4 tip nükleotitten oluşur - adenin (A), urasil (U), guanin (G) ve sitozin (C). Bu durumda, aşağıdaki çiftler tamamlayıcıdır: adenin - urasil, guanin - sitozin. Gördüğünüz gibi, DNA'dan farklı olarak RNA, timin yerine urasil kullanır. Yani “oluşturucu” enzim şu şekilde çalışır: DNA zincirinde A görürse RNA zincirine Y'yi, G ise C'yi bağlar vb. Böylece, transkripsiyon sırasında her aktif genden bir şablon oluşur - nükleer zardan geçebilen bir RNA kopyası. Belirli bir gen tarafından kodlanan bir proteinin sentezi nasıldır?
Çekirdekten ayrıldıktan sonra RNA sitoplazmaya girer. Zaten sitoplazmada, RNA, bir matris olarak, proteinin karşılık gelen amino asit dizisini RNA bilgisi tarafından yönlendirilen sentezleyebilen özel enzim sistemleri (ribozomlar) içine yerleştirilmiş olabilir. Bildiğiniz gibi, bir protein molekülü amino asitlerden oluşur. Ribozom, büyüyen protein zincirine hangi amino asidin bağlanacağını nasıl bilmektedir? Bu, üçlü kod temelinde yapılır. Üçlü kod, RNA zincirinin üç nükleotid dizisinin ( üçlü,örneğin, GGU) bir amino asidi (bu durumda glisin) kodlar. Her amino asit, belirli bir üçlü tarafından kodlanır. Ve böylece, ribozom üçlüyü "okur", bilgi RNA'ya okunurken hangi amino asidin eklenmesi gerektiğini belirler. Bir amino asit zinciri oluştuğunda, belirli bir uzaysal biçim alır ve kendisine atanan enzimatik, yapı, hormonal ve diğer işlevleri yerine getirebilen bir protein haline gelir.Herhangi bir canlı organizma için protein bir gen ürünüdür. Genlerin tüm çeşitli özelliklerini, niteliklerini ve dış tezahürlerini belirleyen proteinlerdir.
Sağda, 23 Nisan 2016'da Guinness Rekorlar Kitabı'na giren Varna'daki (Bulgaristan) sahildeki insanlardan yapılmış en büyük insan DNA sarmalı.
Deoksiribonükleik asit. Genel bilgi
DNA (deoksiribonükleik asit), bir tür yaşam planı, kalıtsal bilgilerle ilgili verileri içeren karmaşık bir koddur. Bu karmaşık makromolekül, kalıtsal genetik bilgiyi nesilden nesile depolama ve aktarma yeteneğine sahiptir. DNA, herhangi bir canlı organizmanın kalıtım ve değişkenlik gibi özelliklerini belirler. İçinde kodlanan bilgiler, herhangi bir canlı organizmanın tüm gelişim programını belirler. Genetik olarak gömülü faktörler, hem bir insanın hem de diğer herhangi bir organizmanın tüm yaşam seyrini önceden belirler. Dış çevrenin yapay veya doğal etkisi, bireysel genetik özelliklerin genel ciddiyetini ancak çok az etkileyebilir veya programlanmış süreçlerin gelişimini etkileyebilir.
Deoksiribonükleik asit(DNA), canlı organizmaların gelişimi ve işleyişi için genetik programın depolanmasını, nesilden nesile aktarılmasını ve uygulanmasını sağlayan bir makromoleküldür (üç ana molekülden biri, diğer ikisi RNA ve proteinlerdir). DNA, çeşitli RNA ve protein türlerinin yapısı hakkında bilgi içerir.
Ökaryotik hücrelerde (hayvanlar, bitkiler ve mantarlar), DNA hücre çekirdeğinde kromozomların bir parçası olarak ve ayrıca bazı hücre organellerinde (mitokondri ve plastitler) bulunur. Prokaryotik organizmaların (bakteriler ve arkeler) hücrelerinde, hücre zarına içeriden nükleoid adı verilen dairesel veya doğrusal bir DNA molekülü eklenir. Onlar ve düşük ökaryotlar (örneğin maya) ayrıca plazmit adı verilen küçük özerk, çoğunlukla dairesel DNA moleküllerine sahiptir.
Kimyasal açıdan DNA, tekrarlayan bloklardan - nükleotidlerden oluşan uzun bir polimerik moleküldür. Her nükleotit, azotlu bir baz, bir şeker (deoksiriboz) ve bir fosfat grubundan oluşur. Bir zincirdeki nükleotitler arasındaki bağlar deoksiriboz tarafından oluşturulur ( İle) ve fosfat ( F) grupları (fosfodiester bağları).

Pirinç. 2. Nüklertit, azotlu bir baz, şeker (deoksiriboz) ve bir fosfat grubundan oluşur.
Vakaların ezici çoğunluğunda (tek sarmallı DNA içeren bazı virüsler hariç), DNA makromolekülü, azotlu bazlar tarafından birbirine yönlendirilmiş iki zincirden oluşur. Bu çift sarmallı molekül bir sarmal içinde bükülür.
DNA'da bulunan dört tip azotlu baz vardır (adenin, guanin, timin ve sitozin). Zincirlerden birinin azotlu bazları, tamamlayıcılık ilkesine göre hidrojen bağları ile diğer zincirin azotlu bazlarına bağlanır: adenin sadece timin ile birleşir ( A-T), guanin - sadece sitozin ile ( G-C). DNA'nın sarmal "merdiveninin" "basamaklarını" oluşturan bu çiftlerdir (bakınız: Şekil 2, 3 ve 4).

Pirinç. 2. Azotlu bazlar
Nükleotit dizisi, ilgili bilgileri "kodlamanıza" izin verir. çeşitli tipler En önemlileri bilgi veya şablon (mRNA), ribozomal (rRNA) ve taşıma (tRNA) olan RNA. Tüm bu RNA türleri, DNA dizisinin transkripsiyon sırasında sentezlenen RNA dizisine kopyalanmasıyla DNA şablonu üzerinde sentezlenir ve protein biyosentezinde (çeviri işlemi) yer alır. Kodlama dizilerine ek olarak, hücre DNA'sı düzenleyici ve yapısal işlevleri yerine getiren diziler içerir.

Pirinç. 3. DNA replikasyonu
DNA kimyasal bileşiklerinin temel kombinasyonlarının yeri ve bu kombinasyonlar arasındaki nicel oranlar, kalıtsal bilgilerin kodlanmasını sağlar.
Eğitim yeni DNA (replikasyon)
- Replikasyon süreci: DNA çift sarmalının çözülmesi - DNA polimeraz tarafından tamamlayıcı ipliklerin sentezi - birinden iki DNA molekülünün oluşumu.
- Enzimler kimyasal bileşiklerin baz çiftleri arasındaki bağı kırdığında çift sarmal iki kola "açılır".
- Her dal yeni bir DNA elementidir. Yeni baz çiftleri, ana daldakiyle aynı sırayla bağlanır.
Kopyalama tamamlandıktan sonra, ana DNA'nın kimyasal bileşiklerinden oluşturulan ve onunla aynı genetik koda sahip iki bağımsız sarmal oluşur. Bu şekilde, DNA hücreden hücreye bilgi parçalayabilir.
Daha detaylı bilgi:
NÜKLEİK ASİTLERİN YAPISI

Pirinç. 4. Azotlu bazlar: adenin, guanin, sitozin, timin
Deoksiribonükleik asit(DNA) nükleik asitleri ifade eder. Nükleik asitler monomerleri nükleotid olan düzensiz biyopolimerlerin bir sınıfıdır.
NÜKLEOTİTLER oluşmaktadır azotlu baz, beş karbonlu bir karbonhidrata (pentoz) bağlı - deoksiriboz(DNA durumunda) veya riboz(RNA durumunda), bir fosforik asit kalıntısı (H2P03 -) ile birleşir.
azotlu bazlarİki tip vardır: pirimidin bazları - urasil (sadece RNA'da), sitozin ve timin, pürin bazları - adenin ve guanin.

Pirinç. Şekil 5. Nükleotitlerin yapısı (solda), nükleotidin DNA'daki yeri (altta) ve azotlu baz türleri (sağda): pirimidin ve pürin

Bir pentoz molekülündeki karbon atomları 1'den 5'e kadar numaralandırılmıştır. Fosfat üçüncü ve beşinci karbon atomlarıyla birleşir. Nükleik asitler bir nükleik asit zinciri oluşturmak için bu şekilde birbirine bağlanır. Böylece DNA zincirinin 3' ve 5' uçlarını izole edebiliriz:

Pirinç. 6. DNA zincirinin 3' ve 5' uçlarının izolasyonu
İki iplikli DNA formu çift sarmal. Bir spiraldeki bu zincirler zıt yönlerde yönlendirilir. DNA'nın farklı zincirlerinde azotlu bazlar birbirine şu şekilde bağlanır: hidrojen bağları. Adenin her zaman timin ile birleşir ve sitozin her zaman guanin ile birleşir. denir tamamlayıcılık kuralı(santimetre. tamamlayıcılık ilkesi).
Tamamlayıcılık kuralı:
| A-T G-C |
Örneğin, diziye sahip bir DNA zinciri verilirse
3'-ATGTCCTAGCTGTCCG - 5',
o zaman ikinci zincir onu tamamlayıcı olacak ve ters yöne yönlendirilecek - 5' ucundan 3' ucuna:
5'- TACAGGATCGACGAGC- 3'.

Pirinç. 7. DNA molekülünün zincirlerinin yönü ve hidrojen bağları kullanarak azotlu bazların bağlanması
DNA KOPYALAMA
DNA kopyalama kalıp sentezi yoluyla bir DNA molekülünü ikiye katlama işlemidir. Çoğu durumda doğal DNA replikasyonuastarDNA sentezi için kısa pasaj (tekrar oluşturuldu). Böyle bir ribonükleotid primeri, enzim primaz (prokaryotlarda DNA primaz, ökaryotlarda DNA polimeraz) tarafından oluşturulur ve daha sonra, normal olarak onarım işlevlerini yerine getiren (DNA molekülündeki kimyasal hasarı ve kırılmaları düzelten) deoksiribonükleotit polimeraz ile değiştirilir.
Çoğaltma yarı muhafazakar bir şekilde gerçekleşir. Bu, DNA'nın çift sarmalının çözüldüğü ve zincirlerinin her birinde tamamlayıcılık ilkesine göre yeni bir zincirin tamamlandığı anlamına gelir. Böylece yavru DNA molekülü, ana molekülden bir iplik ve yeni sentezlenmiş bir iplik içerir. Çoğaltma, ana ipliğin 3' ila 5' yönünde gerçekleşir.

Pirinç. 8. DNA molekülünün replikasyonu (iki katına çıkması)
DNA sentezi- bu ilk bakışta göründüğü kadar karmaşık bir süreç değil. Bunu düşünüyorsanız, önce sentezin ne olduğunu bulmanız gerekir. Bir şeyi bir araya getirme sürecidir. Yeni bir DNA molekülünün oluşumu birkaç aşamada gerçekleşir:
1) Replikasyon çatalının önünde yer alan DNA topoizomeraz, DNA'nın açılmasını ve çözülmesini kolaylaştırmak için DNA'yı keser.
2) Topoizomerazı takip eden DNA sarmalı, DNA sarmalının "açılması" sürecini etkiler.
3) DNA bağlayıcı proteinler, DNA iplikçiklerinin bağlanmasını gerçekleştirir ve ayrıca stabilizasyonlarını gerçekleştirerek birbirlerine yapışmalarını önler.
4) DNA polimeraz δ(delta) , çoğaltma çatalının hareket hızıyla koordineli olarak sentezi gerçekleştirirliderzincirler yan kuruluş Matriks üzerinde 5" → 3" yönünde DNA anne 3" ucundan 5" ucuna doğru DNA iplikçikleri (saniyede 100 baz çiftine kadar hızlanır). Bu olaylar bunun üzerine anne DNA zincirleri sınırlıdır.

Pirinç. 9. DNA replikasyon sürecinin şematik gösterimi: (1) Gecikmeli iplik (gecikmeli iplik), (2) Öncü iplik (öncü iplik), (3) DNA polimeraz α (Pola), (4) DNA ligaz, (5) RNA -primer, (6) Primaz, (7) Okazaki fragmanı, (8) DNA polimeraz δ (Polδ), (9) Helikaz, (10) Tek sarmallı DNA bağlayıcı proteinler, (11) Topoizomeraz.
Gecikmeli kızı DNA zincirinin sentezi aşağıda açıklanmıştır (aşağıya bakınız). şema replikasyon çatalı ve replikasyon enzimlerinin işlevi)
DNA replikasyonu hakkında daha fazla bilgi için bkz.
5) Ana molekülün başka bir ipliğinin çözülmesinden ve stabilizasyonundan hemen sonra, birleşir.DNA polimeraz α(alfa)ve 5 "→3" yönünde bir primer (RNA primeri) sentezler - 10 ila 200 nükleotit uzunluğunda bir DNA şablonu üzerinde bir RNA dizisi. Bundan sonra enzimDNA zincirinden çıkarılır.
Yerine DNA polimerazα
astarın 3" ucuna takılı DNA polimerazε
.
6)
DNA polimerazε
(epsilon) sanki astarı uzatmaya devam ediyor ama bir alt tabaka gömüyormuş gibideoksiribonükleotitler(150-200 nükleotit miktarında). Sonuç olarak, iki parçadan sağlam bir iplik oluşur -RNA(yani astar) ve DNA.
DNA polimeraz εöncekinin astarıyla karşılaşana kadar çalışırparça Okazaki(biraz daha erken sentezlendi). Bu enzim daha sonra zincirden çıkarılır.
7) DNA polimeraz β(beta) yerine duruyorDNA polimerazlar ε,aynı yönde hareket eder (5" → 3") ve yerlerine deoksiribonükleotitleri yerleştirirken primer ribonükleotitleri çıkarır. Enzim, primerin tamamen çıkarılmasına kadar çalışır, yani. bir deoksiribonükleotide kadar (daha önce sentezlenmişDNA polimeraz ε). Enzim, yaptığı işin sonucu ile önündeki DNA'yı bağlayamadığı için zinciri terk eder.
Sonuç olarak, ana ipliğin matrisi üzerinde kızı DNA'nın bir parçası "yatar". denirOkazaki parçası.
8) DNA ligaz, iki bitişik parçalar Okazaki , yani 5 "-segmentin sonu, sentezlendiDNA polimeraz ε,ve 3" zincir ucu yerleşikDNA polimerazβ .
RNA'NIN YAPISI
Ribonükleik asit(RNA), tüm canlı organizmaların hücrelerinde bulunan üç ana makromolekülden (diğer ikisi DNA ve proteinlerdir) biridir.
Tıpkı DNA gibi, RNA da her bir bağlantının adlandırıldığı uzun bir zincirden oluşur. nükleotid. Her nükleotit, azotlu bir baz, bir riboz şekeri ve bir fosfat grubundan oluşur. Bununla birlikte, DNA'dan farklı olarak, RNA'nın genellikle iki yerine bir zinciri vardır. RNA'daki pentoz, deoksiriboz değil, riboz ile temsil edilir (riboz, ikinci karbonhidrat atomunda ek bir hidroksil grubuna sahiptir). Son olarak, DNA, azotlu bazların bileşiminde RNA'dan farklıdır: timin yerine ( T) urasil RNA'da bulunur ( sen) , aynı zamanda adenin için tamamlayıcıdır.
Nükleotit dizisi, RNA'nın genetik bilgiyi kodlamasına izin verir. Tüm hücresel organizmalar, protein sentezini programlamak için RNA (mRNA) kullanır.
Hücresel RNA'lar denilen bir süreçte oluşturulur. transkripsiyon yani, özel enzimler tarafından gerçekleştirilen bir DNA şablonu üzerinde RNA sentezi - RNA polimerazlar.
Messenger RNA'ları (mRNA'lar) daha sonra adı verilen bir süreçte yer alır. yayın yapmak, onlar. ribozomların katılımıyla mRNA şablonu üzerinde protein sentezi. Diğer RNA'lar, transkripsiyondan sonra kimyasal modifikasyonlara uğrarlar ve ikincil ve üçüncül yapıların oluşumundan sonra, RNA tipine bağlı işlevleri yerine getirirler.

Pirinç. 10. Azotlu baz açısından DNA ve RNA arasındaki fark: RNA, timin (T) yerine, aynı zamanda adenin için tamamlayıcı olan urasil (U) içerir.
TRANSKRİPSİYON
Bu, bir DNA şablonu üzerinde RNA sentezi sürecidir. DNA, sitelerden birinde gevşer. Zincirlerden biri, RNA molekülüne kopyalanması gereken bilgileri içerir - bu zincire kodlama denir. Kodlama zincirini tamamlayıcı olan ikinci DNA zincirine şablon iplik denir. 3'-5' yönünde (DNA zinciri boyunca) şablon zincir üzerinde transkripsiyon işleminde, ona tamamlayıcı bir RNA zinciri sentezlenir. Böylece kodlama zincirinin bir RNA kopyası oluşturulur.
![]()
Pirinç. 11. Transkripsiyonun şematik gösterimi
Örneğin, bize kodlama dizisinin dizisi verilirse
3'-ATGTCCTAGCTGTCCG - 5',
daha sonra tamamlayıcılık kuralına göre matris zinciri diziyi taşıyacaktır.
5'- TACAGGATCGACGAGC- 3',
ve ondan sentezlenen RNA dizidir
YAYIN YAPMAK
Mekanizmayı düşünün protein sentezi RNA matrisinin yanı sıra genetik kod ve özellikleri üzerinde. Ayrıca, açıklığa kavuşturmak için aşağıdaki bağlantıda, canlı bir hücrede meydana gelen transkripsiyon ve çeviri süreçleri hakkında kısa bir video izlemenizi öneririz:

Pirinç. 12. Protein sentezi süreci: RNA için DNA kodları, protein için RNA kodları
GENETİK KOD
Genetik Kod- bir nükleotid dizisi kullanarak proteinlerin amino asit dizisini kodlama yöntemi. Her amino asit, bir kodon veya bir üçlü olmak üzere üç nükleotit dizisi tarafından kodlanır.
Çoğu pro ve ökaryot için ortak olan genetik kod. Tablo 64 kodonun tümünü listeler ve karşılık gelen amino asitleri listeler. Baz sıra, mRNA'nın 5" ucundan 3" ucuna kadardır.
Tablo 1. Standart genetik kod
|
1 inci kız |
2. taban |
3 üncü kız |
|||||||
|
sen |
C |
A |
G |
||||||
|
sen |
U U U |
(F/F) |
U C U |
(Ser/S) |
U A U |
(Tir/Y) |
UGU |
(Cys/C) |
sen |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(Leu/L) |
UCCA |
U A A |
Kodonu durdur** |
UGA |
Kodonu durdur** |
A |
||
|
U U G |
UC G |
U A G |
Kodonu durdur** |
U G G |
(Trp/K) |
G |
|||
|
C |
C U U |
CCU |
(Pro/P) |
C A U |
(Onun/H) |
CGU |
(Arg/R) |
sen |
|
|
CUC |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
CGA |
A |
||||
|
CUG |
C C G |
CAG |
C G G |
G |
|||||
|
A |
bir U U |
(Ile/I) |
bir CU |
(Bk/T) |
bir |
(Asn/N) |
bir GÜ |
(Ser/S) |
sen |
|
AUC |
bir C C |
bir A C |
bir G C |
C |
|||||
|
bir U A |
bir |
bir |
(Lys/K) |
bir G A |
A |
||||
|
bir UG |
(Met/E) |
bir CG |
bir G |
bir GG |
G |
||||
|
G |
G U U |
(Val/V) |
G C U |
(Ala/A) |
G A U |
(Asp/D) |
G G U |
(Gli/G) |
sen |
|
GUC |
G C C |
G A C |
GG C |
C |
|||||
|
G U A |
GCA |
G A A |
(Zamk) |
GG A |
A |
||||
|
G U G |
G C G |
GAG |
G G G |
G |
|||||
Üçüzler arasında "noktalama işareti" görevi gören 4 özel dizilim vardır:
- *Üçlü Ağustos, ayrıca metionin kodlayan, denir kodonu başlat. Bu kodon, bir protein molekülünün sentezini başlatır. Böylece protein sentezi sırasında dizideki ilk amino asit her zaman metionin olacaktır.
- **Üçüzler UAA, UAG ve UGA isminde kodonları durdurmak ve herhangi bir amino asit için kodlama yapmayın. Bu dizilerde protein sentezi durur.
Genetik kodun özellikleri
1. Üçlülük. Her amino asit, bir üçlü veya kodon olmak üzere üç nükleotit dizisi tarafından kodlanır.
2. Süreklilik. Üçüzler arasında ek nükleotid yoktur, bilgi sürekli okunur.
3. Örtüşmeyen. Bir nükleotid aynı anda iki üçlünün parçası olamaz.
4. Benzersizlik. Bir kodon sadece bir amino asidi kodlayabilir.

5. Dejenerasyon. Bir amino asit, birkaç farklı kodon tarafından kodlanabilir.

6. Çok yönlülük. Genetik kod tüm canlı organizmalar için aynıdır.
Misal. Bize kodlama dizisinin sırası verildi:
3’- CCGATTGCACGTCGATCGTATA- 5’.
Matris zinciri şu sıraya sahip olacaktır:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
Şimdi bu zincirden bilgi RNA'sını “sentezliyoruz”:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
Protein sentezi 5' → 3' yönünde ilerler, bu nedenle genetik kodu "okumak" için diziyi çevirmemiz gerekir:
5’- AAUUGCUAGCUGCACGUUAGCC- 3’.
Şimdi AUG başlangıç kodonunu bulun:
5’- AU AĞUSTOS CUAGCUGCACGUUAGCC- 3’.
Diziyi üçe bölün:
Kulağa şöyle geliyor: DNA'dan gelen bilgiler RNA'ya (transkripsiyon), RNA'dan proteine (çeviri) aktarılır. DNA, replikasyon yoluyla da kopyalanabilir ve DNA bir RNA şablonundan sentezlendiğinde ters transkripsiyon işlemi de mümkündür, ancak böyle bir işlem esas olarak virüslerin özelliğidir.

Pirinç. 13. Merkezi Dogma moleküler Biyoloji
GENOM: GENLER VE KROMOZOMLAR
(Genel konseptler)
Genom - bir organizmanın tüm genlerinin toplamı; tam kromozom seti.
"Genom" terimi, 1920'de G. Winkler tarafından aynı biyolojik türden organizmaların haploid kromozom setinde bulunan genlerin toplamını tanımlamak için önerildi. Bu terimin orijinal anlamı, genotipin aksine genom kavramının bir bireyin değil, bir bütün olarak türün genetik bir özelliği olduğunu gösterdi. Moleküler genetiğin gelişmesiyle birlikte bu terimin anlamı değişti. Çoğu organizmada genetik bilginin taşıyıcısı olan ve dolayısıyla genomun temelini oluşturan DNA'nın, kelimenin modern anlamıyla sadece genleri içermediği bilinmektedir. ÇoğuÖkaryotik hücrelerin DNA'sı, proteinler ve nükleik asitler hakkında bilgi içermeyen, kodlamayan (“artık”) nükleotit dizileriyle temsil edilir. Bu nedenle, herhangi bir organizmanın genomunun ana kısmı, haploid kromozom setinin tüm DNA'sıdır.
Genler, polipeptitleri ve RNA moleküllerini kodlayan DNA moleküllerinin segmentleridir.
Geçen yüzyılda, gen anlayışımız önemli ölçüde değişti. Daha önce genom, bir kromozomun bir özelliği ya da bir özelliği kodlayan ya da belirleyen bir bölgesiydi. fenotipik(görünür) özellik, örneğin göz rengi.
1940'ta George Beadle ve Edward Tatham, bir genin moleküler bir tanımını önerdiler. Bilim adamları mantar sporlarını işledi nörospora crassa DNA dizisinde değişikliklere neden olan X-ışınları ve diğer ajanlar ( mutasyonlar) ve bazı spesifik enzimleri kaybeden, bazı durumlarda tüm metabolik yolun bozulmasına yol açan mantarın mutant suşları buldu. Beadle ve Tatham, bir genin, tek bir enzimi tanımlayan veya kodlayan genetik materyalin bir bölümü olduğu sonucuna vardı. Hipotez böyle "bir gen, bir enzim". Bu kavram daha sonra tanımına genişletildi "bir gen - bir polipeptit" Birçok gen, enzim olmayan proteinleri kodladığından ve bir polipeptit, karmaşık bir protein kompleksinin alt birimi olabilir.
Şek. Şekil 14, DNA'daki üçlü nükleotitlerin, mRNA'nın aracılık ettiği bir proteinin amino asit dizisi olan bir polipeptidi nasıl belirlediğini gösteren bir diyagramı göstermektedir. DNA zincirlerinden biri, nükleotid üçlüleri (kodonları) DNA üçlülerini tamamlayıcı olan mRNA'nın sentezi için bir şablon rolü oynar. Bazı bakterilerde ve birçok ökaryotta, kodlama dizileri, kodlamayan bölgeler tarafından kesintiye uğrar. intronlar).
Bir genin modern biyokimyasal tanımı hatta daha spesifik olarak. Genlerin tümü, yapısal veya katalitik bir işlevi olan polipeptitler veya RNA içeren son ürünlerin birincil dizisini kodlayan DNA bölümleridir.
DNA, genlerle birlikte, yalnızca düzenleyici bir işlevi yerine getiren başka diziler de içerir. düzenleyici diziler genlerin başlangıcını veya sonunu işaretleyebilir, transkripsiyonu etkileyebilir veya replikasyon veya rekombinasyonun başladığı yeri gösterebilir. Bazı genler, farklı ürünlerin oluşumu için bir şablon görevi gören aynı DNA parçası ile farklı şekillerde ifade edilebilir.
kabaca hesaplayabiliriz minimum gen boyutu ara proteini kodlar. Bir polipeptit zincirindeki her amino asit, üç nükleotitlik bir dizi tarafından kodlanır; bu üçlülerin (kodonlar) dizileri, verilen gen tarafından kodlanan polipeptitteki amino asit zincirine karşılık gelir. 350 amino asit kalıntısından oluşan bir polipeptit zinciri (orta uzunlukta zincir), 1050 bp'lik bir diziye karşılık gelir. ( bp). Bununla birlikte, birçok ökaryotik gen ve bazı prokaryotik genler, DNA segmentleri tarafından kesintiye uğratılır. bilgi taşıyıcıları protein hakkında ve bu nedenle basit bir hesaplamanın gösterdiğinden çok daha uzun olduğu ortaya çıktı.
Bir kromozomda kaç gen vardır?
 Pirinç. 15. Prokaryotik (solda) ve ökaryotik hücrelerde kromozomların görünümü. Histonlar, iki ana işlevi yerine getiren geniş bir nükleer protein sınıfıdır: çekirdekteki DNA ipliklerinin paketlenmesinde ve transkripsiyon, replikasyon ve onarım gibi nükleer süreçlerin epigenetik düzenlenmesinde yer alırlar.
Pirinç. 15. Prokaryotik (solda) ve ökaryotik hücrelerde kromozomların görünümü. Histonlar, iki ana işlevi yerine getiren geniş bir nükleer protein sınıfıdır: çekirdekteki DNA ipliklerinin paketlenmesinde ve transkripsiyon, replikasyon ve onarım gibi nükleer süreçlerin epigenetik düzenlenmesinde yer alırlar.
Prokaryotların DNA'sı daha basittir: hücrelerinin çekirdeği yoktur, bu nedenle DNA doğrudan sitoplazmada bir nükleoid şeklinde bulunur.
 Bildiğiniz gibi, bakteri hücrelerinin, kompakt bir yapıya - bir nükleoid - paketlenmiş bir DNA dizisi şeklinde bir kromozomu vardır. prokaryotik kromozom Escherichia koli genomu tamamen çözülmüş olan, dairesel bir DNA molekülüdür (aslında, sağ daire, daha ziyade başı veya sonu olmayan bir döngü), 4.639.675 b.p. Bu dizi, yaklaşık 4300 protein geni ve kararlı RNA molekülleri için başka 157 gen içerir. AT insan genomu 24 farklı kromozom üzerinde yer alan yaklaşık 29.000 gene karşılık gelen yaklaşık 3,1 milyar baz çifti.
Bildiğiniz gibi, bakteri hücrelerinin, kompakt bir yapıya - bir nükleoid - paketlenmiş bir DNA dizisi şeklinde bir kromozomu vardır. prokaryotik kromozom Escherichia koli genomu tamamen çözülmüş olan, dairesel bir DNA molekülüdür (aslında, sağ daire, daha ziyade başı veya sonu olmayan bir döngü), 4.639.675 b.p. Bu dizi, yaklaşık 4300 protein geni ve kararlı RNA molekülleri için başka 157 gen içerir. AT insan genomu 24 farklı kromozom üzerinde yer alan yaklaşık 29.000 gene karşılık gelen yaklaşık 3,1 milyar baz çifti.
Prokaryotlar (Bakteriler).
 bakteri E. koli bir çift sarmallı dairesel DNA molekülüne sahiptir. 4.639.675 b.p'den oluşur. ve hücrenin kendisinin uzunluğunu aşan yaklaşık 1,7 mm uzunluğa ulaşır E. koli yaklaşık 850 kez. Nükleoidin bir parçası olan büyük dairesel kromozoma ek olarak, birçok bakteri sitozolde serbestçe bulunan bir veya daha fazla küçük dairesel DNA molekülü içerir. Bu ekstrakromozomal elementlere denir. plazmitler(Şek. 16).
bakteri E. koli bir çift sarmallı dairesel DNA molekülüne sahiptir. 4.639.675 b.p'den oluşur. ve hücrenin kendisinin uzunluğunu aşan yaklaşık 1,7 mm uzunluğa ulaşır E. koli yaklaşık 850 kez. Nükleoidin bir parçası olan büyük dairesel kromozoma ek olarak, birçok bakteri sitozolde serbestçe bulunan bir veya daha fazla küçük dairesel DNA molekülü içerir. Bu ekstrakromozomal elementlere denir. plazmitler(Şek. 16).
Çoğu plazmit sadece birkaç bin baz çiftinden oluşur, bazıları 10.000'den fazla baz çifti içerir. Genetik bilgiyi taşırlar ve ana hücrenin bölünmesi sırasında yavru hücrelere giren yavru plazmitleri oluşturmak için çoğalırlar. Plazmitler sadece bakterilerde değil, maya ve diğer mantarlarda da bulunur. Çoğu durumda, plazmitler konakçı hücrelere hiçbir avantaj sağlamaz ve tek görevleri bağımsız olarak çoğalmaktır. Bununla birlikte, bazı plazmitler, konakçı için yararlı olan genleri taşır. Örneğin, plazmitlerde bulunan genler, bakteri hücrelerinde antibakteriyel maddelere direnç kazandırabilir. β-laktamaz genini taşıyan plazmitler, penisilin ve amoksisilin gibi β-laktam antibiyotiklere direnç kazandırır. Plazmitler, antibiyotiğe dirençli hücrelerden aynı veya farklı bakteri türlerinin diğer hücrelerine geçerek bu hücrelerin de dirençli hale gelmesine neden olabilir. Yoğun antibiyotik kullanımı, patojenik bakteriler arasında antibiyotik direncini kodlayan plazmitlerin (ve benzer genleri kodlayan transpozonların) yayılmasını destekleyen ve çeşitli antibiyotiğe dirençli bakteri suşlarının ortaya çıkmasına neden olan güçlü bir seçici faktördür. Doktorlar, yaygın antibiyotik kullanımının tehlikelerini anlamaya başlıyor ve bunları yalnızca kesinlikle gerekli olduğunda reçete ediyor. Benzer nedenlerle, çiftlik hayvanlarının tedavisinde antibiyotiklerin yaygın kullanımı sınırlıdır.
Ayrıca bakınız: Ravin N.V., Shestakov S.V. Prokaryotların genomu // Vavilov Journal of Genetics and Breeding, 2013. V. 17. No. 4/2. s. 972-984.
ökaryotlar.
Tablo 2. Bazı organizmaların DNA, gen ve kromozomları
|
paylaşılan DNA, b.s. |
Kromozom sayısı* |
Yaklaşık gen sayısı |
|
|
Escherichia koli(bakteri) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(Maya) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(nematod) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(bitki) |
119 186 200 |
33 000 |
|
|
Drosophila melanogaster(Meyve sineği) |
120 367 260 |
20 000 |
|
|
Oryza sativa(pilav) |
480 000 000 |
57 000 |
|
|
Muş kası(fare) |
2 634 266 500 |
27 000 |
|
|
homo sapiens(İnsan) |
3 070 128 600 |
29 000 |
Not. Bilgi sürekli güncellenir; Daha güncel bilgiler için bireysel genomik proje web sitelerine bakın.
* Maya hariç tüm ökaryotlar için diploid kromozom seti verilmiştir. diploit takım kromozomlar (Yunanca diploos - çift ve eidos - görünümden) - çift kromozom seti(2n), her biri kendi içinde bir homolojiye sahiptir.
** Haploid seti. Yabani maya türleri tipik olarak sekiz (oktaploid) veya daha fazla bu kromozom setine sahiptir.
***İki X kromozomlu dişiler için. Erkeklerin bir X kromozomu vardır, ancak Y yoktur, yani sadece 11 kromozom.
En küçük ökaryotlardan biri olan bir maya hücresi, bir hücreden 2,6 kat daha fazla DNA'ya sahiptir. E. koli(Tablo 2). meyve sineği hücreleri Meyve sineği Genetik araştırmaların klasik bir nesnesi olan, 35 kat daha fazla DNA içerir ve insan hücreleri, hücrelerden yaklaşık 700 kat daha fazla DNA içerir. E. koli. Birçok bitki ve amfibi daha da fazla DNA içerir. Ökaryotik hücrelerin genetik materyali, kromozomlar şeklinde düzenlenir. Diploid kromozom seti (2 n) organizmanın tipine bağlıdır (Tablo 2).
Örneğin, bir insan somatik hücresinde 46 kromozom vardır ( pilav. 17). Ökaryotik bir hücredeki her bir kromozom, Şekil 1'de gösterildiği gibi. 17, a, çok büyük bir çift sarmallı DNA molekülü içerir. Yirmi dört insan kromozomu (22 çift kromozom ve iki cinsiyet kromozomu X ve Y) uzunluk olarak 25 kattan fazla farklılık gösterir. Her ökaryotik kromozom, belirli bir gen kümesi içerir.

Pirinç. 17. ökaryotik kromozomlar.a- insan kromozomundan bir çift bağlı ve yoğunlaştırılmış kardeş kromatit. Bu formda ökaryotik kromozomlar replikasyondan sonra ve mitoz sırasında metafazda kalır. b- kitabın yazarlarından birinin lökositinden tam bir kromozom seti. Her normal insan somatik hücresi 46 kromozom içerir.

Kalıtsal materyali depolamak ve iletmek için bir matris olarak DNA'nın boyutu ve işlevi, bu molekülün organizasyonunda özel yapısal elemanların varlığını açıklar. Daha yüksek organizmalarda, DNA kromozomlar arasında dağıtılır.
Bir organizmanın DNA kümesine (kromozomlar) genom denir. Kromozomlar hücre çekirdeğinde bulunur ve kromatin adı verilen bir yapı oluşturur. Kromatin, 1: 1 oranında bir DNA ve bazik proteinler (histonlar) kompleksidir. DNA'nın uzunluğu genellikle tamamlayıcı nükleotid çiftlerinin (bp) sayısı ile ölçülür. Örneğin, 3. insan kromozomuyüzyıl 160 milyon bp büyüklüğünde bir DNA molekülüdür. yaklaşık 1 mm uzunluğa sahiptir, bu nedenle, 3. insan kromozomunun doğrusallaştırılmış bir molekülünün uzunluğu 5 mm olacaktır ve bir haploidin 23 kromozomunun tamamının (~ 3 * 109 bp, MR = 1.8 * 10 12) DNA'sı olacaktır. hücre - yumurta veya sperm hücresi - doğrusallaştırılmış bir biçimde 1 m olacaktır Germ hücreleri hariç, insan vücudunun tüm hücreleri (yaklaşık 1013 tanesi vardır) çift kromozom seti içerir. Hücre bölünmesi sırasında, 46 DNA molekülünün tamamı replike olur ve 46 kromozom halinde yeniden düzenlenir.
İnsan genomunun DNA moleküllerini (22 kromozom ve X ve Y veya X ve X kromozomları) bağlarsanız, yaklaşık bir metre uzunluğunda bir dizi elde edersiniz. Not: Tüm memelilerde ve diğer heterogametik erkek organizmalarda dişilerde iki X kromozomu (XX) ve erkeklerde bir X kromozomu ve bir Y kromozomu (XY) bulunur.
Çoğu insan hücresi, yani bu tür hücrelerin toplam DNA uzunluğu yaklaşık 2 m'dir. Yetişkin bir insanın yaklaşık 10 14 hücresi vardır, bu nedenle tüm DNA moleküllerinin toplam uzunluğu 2・10 11 km'dir. Karşılaştırma için, Dünya'nın çevresi 4・10 4 km ve Dünya'dan Güneş'e olan mesafe 1.5・10 8 km'dir. Hücrelerimizde inanılmaz derecede kompakt bir şekilde paketlenmiş DNA budur!
Ökaryotik hücrelerde DNA içeren başka organeller vardır - bunlar mitokondri ve kloroplastlardır. Mitokondriyal ve kloroplast DNA'sının kökeni hakkında birçok hipotez ortaya atılmıştır. Günümüzde genel kabul gören bakış açısı, bunların konakçı hücrelerin sitoplazmasına nüfuz eden ve bu organellerin öncüsü haline gelen eski bakterilerin kromozomlarının temelleri oldukları yönündedir. Mitokondriyal DNA, mitokondriyal tRNA ve rRNA'nın yanı sıra birkaç mitokondriyal proteini kodlar. Mitokondriyal proteinlerin %95'inden fazlası nükleer DNA tarafından kodlanır.
GENLERİN YAPISI
Prokaryotlarda ve ökaryotlarda genin yapısını, benzerliklerini ve farklılıklarını düşünün. Bir gen, DNA'nın sadece bir proteini veya RNA'yı kodlayan bir bölümü olmasına rağmen, doğrudan kodlama kısmına ek olarak, prokaryot ve ökaryotlarda farklı yapıya sahip düzenleyici ve diğer yapısal unsurları da içerir.
kodlama dizisi- genin ana yapısal ve fonksiyonel birimi, içinde kodlayan nükleotitlerin üçlüleriamino asit dizisi. Bir başlangıç kodonu ile başlar ve bir durdurma kodonu ile biter.
Kodlama dizisinden önce ve sonra çevrilmemiş 5' ve 3' diziler. Düzenleyici ve yardımcı işlevleri yerine getirirler, örneğin ribozomun mRNA'ya inmesini sağlarlar.
Çevrilmemiş ve kodlama dizileri, transkripsiyon birimini oluşturur - kopyalanan DNA bölgesi, yani mRNA'nın sentezlendiği DNA bölgesi.
sonlandırıcı Bir genin sonunda, RNA sentezinin durduğu, kopyalanmayan bir DNA bölgesi.
Genin başlangıcında düzenleyici alan, içerir destekçi ve Şebeke.
destekçi- transkripsiyonun başlaması sırasında polimerazın bağlandığı dizi. Şebeke- bu, özel proteinlerin bağlanabileceği alandır - baskılayıcılar, bu genden RNA sentezinin aktivitesini azaltabilen - başka bir deyişle, onu azaltan ifade.
Prokaryotlarda genlerin yapısı
Prokaryotlarda ve ökaryotlarda genlerin yapısı için genel plan farklı değildir - her ikisi de bir promotör ve operatör içeren bir düzenleyici bölge, kodlama ve çevrilmemiş dizilere sahip bir transkripsiyon birimi ve bir sonlandırıcı içerir. Ancak prokaryotlarda ve ökaryotlarda genlerin organizasyonu farklıdır.

Pirinç. 18. Prokaryotlarda (bakteri) genin yapısının şeması -resim büyütülmüş
Operonun başında ve sonunda, birkaç yapısal gen için ortak düzenleyici bölgeler vardır. Operonun kopyalanan bölgesinden, her biri kendi başlangıç ve bitiş kodonu olan birkaç kodlama dizisi içeren bir mRNA molekülü okunur. Bu alanların her birindenbir protein sentezlenir. Böylece, Bir i-RNA molekülünden birkaç protein molekülü sentezlenir.
Prokaryotlar, birkaç genin tek bir işlevsel birime kombinasyonu ile karakterize edilir - operon. Operonun çalışması, operonun kendisinden gözle görülür şekilde çıkarılabilen diğer genler tarafından düzenlenebilir - düzenleyiciler. Bu genden çevrilen proteine ne ad verilir? baskılayıcı. Operonun operatörüne bağlanır ve içerdiği tüm genlerin ifadesini bir kerede düzenler.
Prokaryotlar ayrıca fenomen ile karakterize edilir transkripsiyon ve çeviri çekimleri.
![]()
Pirinç. 19 Prokaryotlarda transkripsiyon ve translasyonun konjugasyonu olgusu - resim büyütülmüş
Bu eşleşme, translasyonun gerçekleştiği sitoplazmayı, üzerinde transkripsiyonun gerçekleştiği genetik materyalden ayıran bir nükleer zarfın varlığından dolayı ökaryotlarda gerçekleşmez. Prokaryotlarda, bir DNA şablonu üzerinde RNA sentezi sırasında, bir ribozom sentezlenen RNA molekülüne hemen bağlanabilir. Böylece, çeviri, transkripsiyon tamamlanmadan önce başlar. Ayrıca, birkaç ribozom, aynı anda bir proteinin birkaç molekülünü sentezleyerek bir RNA molekülüne aynı anda bağlanabilir.
Ökaryotlarda genlerin yapısı
Ökaryotların genleri ve kromozomları çok karmaşık bir şekilde organize edilmiştir.
Pek çok türün bakterisinin sadece bir kromozomu vardır ve hemen hemen her durumda her kromozomda her genin bir kopyası vardır. rRNA genleri gibi sadece birkaç gen, çoklu kopyalarda bulunur. Genler ve düzenleyici diziler, prokaryotların neredeyse tüm genomunu oluşturur. Ayrıca, hemen hemen her gen, kodladığı amino asit dizisine (veya RNA dizisine) kesin olarak karşılık gelir (Şekil 14).
Ökaryotik genlerin yapısal ve işlevsel organizasyonu çok daha karmaşıktır. Ökaryotik kromozomların incelenmesi ve daha sonra tam ökaryotik genom dizilerinin dizilenmesi birçok sürpriz getirdi. Çoğu olmasa da çoğu ökaryotik genin ilginç bir özelliği vardır: onların nükleotid dizileri, polipeptit ürününün amino asit dizisini kodlamayan bir veya daha fazla DNA bölgesi içerir. Bu tür çevrilmemiş ekler, genin nükleotid dizisi ile kodlanmış polipeptidin amino asit dizisi arasındaki doğrudan yazışmayı bozar. Genlerdeki bu çevrilmemiş bölümlere denir. intronlar, veya yerleşik diziler, ve kodlama bölümleri ekzonlar. Prokaryotlarda sadece birkaç gen intron içerir.
Bu nedenle, ökaryotlarda, genlerin operonlara kombinasyonu pratik olarak yoktur ve ökaryotik bir genin kodlama dizisi çoğunlukla çevrilmiş bölgelere bölünür. - ekzonlar, ve çevrilmemiş bölümler - intronlar.
Çoğu durumda, intronların işlevi belirlenmemiştir. Genel olarak, insan DNA'sının yalnızca yaklaşık %1,5'i "kodlama"dır, yani proteinler veya RNA hakkında bilgi taşır. Ancak büyük intronlar dikkate alındığında, insan DNA'sının %30'unun genlerden oluştuğu ortaya çıkıyor. Genler, insan genomunun nispeten küçük bir bölümünü oluşturduğundan, önemli miktarda DNA açıklanmamıştır.

Pirinç. 16. Ökaryotlarda gen yapısının şeması - resim büyütülmüş
Her genden, önce hem intronları hem de eksonları içeren olgunlaşmamış veya ön RNA sentezlenir.
Bundan sonra, intron bölgelerinin eksize edildiği ve bir proteinin sentezlenebileceği olgun bir mRNA'nın oluşturulduğu ekleme işlemi gerçekleşir.

Pirinç. 20. Alternatif ekleme işlemi - resim büyütülmüş
Bu tür bir gen organizasyonu, örneğin, ekzonların ekleme sırasında farklı dizilerde kaynaşabilmesi nedeniyle bir genden farklı protein formlarının sentezlenebilmesine olanak tanır.

Pirinç. 21. Prokaryot ve ökaryot genlerinin yapısındaki farklılıklar - resim büyütülmüş
MUTASYONLAR VE MUTAGENEZ
mutasyon genotipte kalıcı bir değişiklik, yani nükleotid dizisindeki bir değişiklik olarak adlandırılır.
Mutasyona yol açan sürece denir mutajenez ve organizma Tümü hücreleri aynı mutasyonu taşıyan mutant.
mutasyon teorisiİlk olarak 1903 yılında Hugh de Vries tarafından formüle edilmiştir. Modern versiyonu aşağıdaki hükümleri içerir:
1. Mutasyonlar aniden, aniden ortaya çıkar.
2. Mutasyonlar nesilden nesile aktarılır.
3. Mutasyonlar faydalı, zararlı veya nötr, baskın veya çekinik olabilir.
4. Mutasyonları tespit etme olasılığı, çalışılan bireylerin sayısına bağlıdır.
5. Benzer mutasyonlar tekrar tekrar meydana gelebilir.
6. Mutasyonlar yönlendirilmez.
Mutasyonlar çeşitli faktörlerin etkisi altında meydana gelebilir. neden olduğu mutasyonları ayırt eder. mutajenik etkiler: fiziksel (örneğin ultraviyole veya radyasyon), kimyasal (örneğin kolşisin veya reaktif oksijen türleri) ve biyolojik (örneğin virüsler). Mutasyonlar da neden olabilir çoğaltma hataları.
Mutasyonların ortaya çıkması için koşullara bağlı olarak ayrılır doğal- yani normal şartlar altında ortaya çıkan mutasyonlar ve uyarılmış- yani, özel koşullar altında ortaya çıkan mutasyonlar.
Mutasyonlar sadece nükleer DNA'da değil, örneğin mitokondri veya plastidlerin DNA'sında da meydana gelebilir. Buna göre ayırt edebiliriz nükleer ve sitoplazmik mutasyonlar.
Mutasyonların ortaya çıkmasının bir sonucu olarak, sıklıkla yeni aleller ortaya çıkabilir. Mutant allel normal alleli geçersiz kılarsa mutasyon denir. baskın. Normal alel mutasyona uğramış olanı baskılarsa, mutasyon denir. çekinik. Yeni alellere yol açan mutasyonların çoğu çekiniktir.
Mutasyonlar etki ile ayırt edilir uyarlanabilir organizmanın çevreye uyum yeteneğinin artmasına neden olur, doğal hayatta kalmayı etkilemeyen zararlı organizmaların çevresel koşullara uyum kabiliyetini azaltan ve öldürücü gelişimin erken evrelerinde organizmanın ölümüne yol açar.
Sonuçlara göre, mutasyonlar ayırt edilir ve protein fonksiyon kaybı neden olan mutasyonlar ortaya çıkma proteinin yeni bir işlevi var mutasyonların yanı sıra bir genin dozunu değiştirmek ve buna göre, ondan sentezlenen protein dozu.
Vücudun herhangi bir hücresinde bir mutasyon meydana gelebilir. Bir germ hücresinde mutasyon meydana gelirse buna denir. tohum(germinal veya üretken). Bu tür mutasyonlar ortaya çıktıkları organizmada ortaya çıkmazlar, ancak yavrularda mutantların ortaya çıkmasına neden olur ve kalıtsaldır, bu nedenle genetik ve evrim için önemlidirler. Mutasyon başka bir hücrede meydana gelirse buna denir. somatik. Böyle bir mutasyon, ortaya çıktığı organizmada bir dereceye kadar kendini gösterebilir, örneğin kanserli tümörlerin oluşumuna yol açar. Bununla birlikte, böyle bir mutasyon kalıtsal değildir ve yavruları etkilemez.
Mutasyonlar, genomun farklı boyutlardaki kısımlarını etkileyebilir. tahsis genetik, kromozomal ve genomik mutasyonlar.
gen mutasyonları
Bir genden daha küçük ölçekte meydana gelen mutasyonlara mutasyon denir. genetik, veya noktalı (noktalı). Bu tür mutasyonlar, dizideki bir veya daha fazla nükleotitte bir değişikliğe yol açar. Gen mutasyonları şunları içerir:ikameler bir nükleotidin diğeriyle yer değiştirmesine yol açan,silmeler nükleotidlerden birinin kaybına yol açan,eklemeler diziye fazladan bir nükleotidin eklenmesine yol açar.

Pirinç. 23. Gen (nokta) mutasyonları
Protein üzerindeki etki mekanizmasına göre, gen mutasyonları ayrılır:eşanlamlı(genetik kodun dejenerasyonunun bir sonucu olarak) protein ürününün amino asit bileşiminde bir değişikliğe yol açmayan,yanlış anlamlı mutasyonlar bir amino asidin bir başkasıyla değiştirilmesine yol açan ve sentezlenen proteinin yapısını etkileyebilen, çoğu zaman önemsiz olmalarına rağmen,anlamsız mutasyonlar, kodlama kodonunun bir durdurma kodonu ile değiştirilmesine yol açar,yol açan mutasyonlar ekleme bozukluğu:

Pirinç. 24. Mutasyon şemaları
Ayrıca, protein üzerindeki etki mekanizmasına göre, mutasyonlar izole edilir ve aşağıdakilere yol açar: çerçeve kaydırma okumalar eklemeler ve silmeler gibi. Bu tür mutasyonlar, saçma mutasyonlar gibi, genin bir noktasında meydana gelmelerine rağmen, genellikle proteinin tüm yapısını etkiler ve bu da yapısında tam bir değişikliğe yol açabilir. kromozomun bir parçası 180 derece döndüğünde Pirinç. 28. Yer Değiştirme
kromozomun bir parçası 180 derece döndüğünde Pirinç. 28. Yer Değiştirme
Pirinç. 29. Çoğaltmadan önce ve sonra kromozom
genomik mutasyonlar
En sonunda, genomik mutasyonlar tüm genomu etkiler, yani kromozom sayısı değişir. Poliploidi ayırt edilir - hücrenin ploidisinde bir artış ve anöploidi, yani kromozom sayısında bir değişiklik, örneğin trizomi (kromozomlardan birinde ek bir homologun varlığı) ve monozomi (yokluğu) kromozomda bir homolog).
DNA ile ilgili video
DNA REPLİKASYONU, RNA KODLAMASI, PROTEİN SENTEZİ
(Video görüntülenmiyorsa, şurada mevcuttur:
DNA'nın kimyasal yapısına göre ( Deoksiribonükleik asit) bir biyopolimer, monomerleri olan nükleotidler. Yani, DNA polinükleotit. Ayrıca, DNA molekülü genellikle sarmal bir çizgi boyunca birbirine göre bükülmüş (genellikle "sarmal bükülmüş" olarak adlandırılır) ve hidrojen bağları ile birbirine bağlanmış iki zincirden oluşur.
Zincirler hem sola hem de sağa (çoğunlukla) bükülebilir.
Bazı virüslerin tek zincirli DNA'sı vardır.
Her DNA nükleotidi 1) azotlu bir baz, 2) deoksiriboz, 3) bir fosforik asit kalıntısından oluşur.
Çift sağ elini kullanan DNA sarmalı
DNA şunları içerir: adenin, guanin, timin ve sitozin. Adenin ve guanin vardır pürinler, ve timin ve sitozin - için pirimidinler. Bazen DNA, genellikle RNA'nın özelliği olan ve timinin yerini aldığı urasil içerir.
Bir DNA molekülünün bir zincirinin azotlu bazları, bir diğerinin azotlu bazlarına kesinlikle tamamlayıcılık ilkesine göre bağlanır: adenin sadece timinle (kendi aralarında iki hidrojen bağı oluştururlar) ve guanin sadece sitozinle (üç bağ).
Nükleotidin kendisindeki azotlu baz, siklik formun ilk karbon atomuna bağlanır. deoksiriboz bir pentozdur (beş karbon atomlu karbonhidrat). Bağ kovalenttir, glikozidiktir (C-N). Ribozdan farklı olarak, deoksiriboz hidroksil gruplarından birine sahip değildir. Deoksiriboz halkası dört karbon atomu ve bir oksijen atomundan oluşur. Beşinci karbon atomu halkanın dışındadır ve bir oksijen atomu aracılığıyla bir fosforik asit kalıntısına bağlanır. Ayrıca, üçüncü karbon atomundaki oksijen atomu aracılığıyla, komşu nükleotidin fosforik asit tortusu eklenir.
Böylece, bir DNA zincirinde, bitişik nükleotitler birbirine bağlıdır. kovalent bağlar deoksiriboz ve fosforik asit arasında (fosfodiester bağı). Bir fosfat-deoksiriboz omurgası oluşur. Buna dik, başka bir DNA ipliğine doğru, ikinci ipliğin bazlarına hidrojen bağları ile bağlanan azotlu bazlar yönlendirilir.
DNA'nın yapısı, hidrojen bağlarıyla birbirine bağlanan zincirlerin omurgaları farklı yönlere yönlendirilecek şekildedir ("çok yönlü", "antiparalel" derler). Birinin deoksiribozun beşinci karbon atomuna bağlı fosforik asit ile bittiği tarafta, diğeri ise "serbest" bir üçüncü karbon atomuyla bitiyor. Yani, bir zincirin iskeleti, diğerine göre olduğu gibi baş aşağı çevrilir. Böylece DNA zincirlerinin yapısında 5 "uç ve 3" uç ayırt edilir.
DNA'yı kopyalarken (ikiye katlarken), yeni zincirlerin sentezi her zaman 5. uçlarından üçüncülerine doğru ilerler, çünkü yeni nükleotitler sadece serbest üçüncü uca eklenebilir.
Nihayetinde (dolaylı olarak RNA yoluyla), DNA zincirindeki her ardışık üç nükleotit, proteinin bir amino asidini kodlar.
DNA molekülünün yapısının keşfi, 1953'te F. Crick ve D. Watson'ın (diğer bilim adamlarının erken çalışmaları tarafından da kolaylaştırılan) çalışmaları sayesinde gerçekleşti. Her ne kadar nasıl Kimyasal madde DNA 19. yüzyıldan beri bilinmektedir. 1940'larda DNA'nın genetik bilginin taşıyıcısı olduğu anlaşıldı.
Çift sarmal, DNA molekülünün ikincil yapısı olarak kabul edilir. Ökaryotik hücrelerde, DNA'nın ezici bir miktarı, proteinler ve diğer maddelerle ilişkili olduğu kromozomlarda bulunur ve ayrıca daha yoğun bir paketlemeye tabi tutulur.
Deoksiribonükleik asit veya DNA genetik bilginin taşıyıcısıdır. Hücrelerdeki DNA'nın çoğu çekirdekte bulunur. Kromozomların ana bileşenidir. Ökaryotlarda DNA, mitokondri ve plastidlerde de bulunur. DNA, uzun dalsız bir polimeri temsil eden, birbirine kovalent olarak bağlı mononükleotitlerden oluşur. DNA'yı oluşturan mononükleotitler, 4 azotlu bazdan (adenin, guanin, sitozin ve timin) biri olan deoksiriboz ve bir fosforik asit kalıntısından oluşur. Bu mononükleotidlerin sayısı çok fazladır. Örneğin, tek bir kromozom içeren prokaryotik hücrelerde DNA, moleküler ağırlığı 2 x 109'dan büyük olan tek bir makromoleküldür.
Bir DNA zincirinin mononükleotitleri, oluşum nedeniyle birbirine seri olarak bağlanır. kovalent fosfodiester bağları bir mononükleotidin deoksiriboz OH grubu ile diğerinin fosforik asit tortusu arasında. Bir DNA zincirinin oluşturduğu omurganın bir tarafında azotlu bazlar bulunur. Çünkü bir ipe konan dört farklı boncukla karşılaştırılabilirler. adeta bir şeker-fosfat zincirine bağlılar.
Soru ortaya çıkıyor, bu uzun polinükleotit zinciri, bir hücrenin veya hatta tüm organizmanın gelişimi için programı nasıl kodlayabilir? Bu sorunun cevabı, DNA'nın uzamsal yapısının nasıl oluştuğunun anlaşılmasıyla elde edilebilir. Bu molekülün yapısı 1953 yılında J. Watson ve F. Crick tarafından deşifre edilmiş ve tanımlanmıştır.
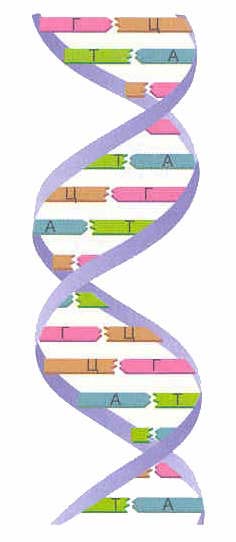
DNA molekülleri birbirine paralel olan ve şekillenen iki zincirdir. sağ elini kullanan sarmal
. Bu spiralin genişliği yaklaşık 2 nm'dir, ancak uzunluğu yüz binlerce nanometreye ulaşabilir. Watson ve Crick, tüm DNA bazlarının sarmalın içinde yer aldığı, şeker-fosfat omurgasının dışarıda olduğu bir DNA modeli önerdi. Böylece bir zincirin tabanları diğerinin tabanlarına mümkün olduğunca yakındır,
Böylece aralarında hidrojen bağları oluşur. DNA sarmalının yapısı öyledir ki onu oluşturan polinükleotid zincirleri ancak çözüldükten sonra ayrılabilir.
İki DNA dizisinin maksimum yakınlığı nedeniyle, bileşimi aynı miktarda bir tür azotlu baz (adenin ve guanin) ve başka bir tür azotlu baz (timin ve sitozin) içerir, yani formül geçerlidir: A+G=T+C. Bunun nedeni azotlu bazların boyutudur, yani adenin-timin ve guanin-sitozin çiftleri arasında hidrojen bağı oluşması nedeniyle oluşan yapıların uzunluğu yaklaşık 1,1 nm'dir. Bu çiftlerin toplam boyutları, DNA sarmalının iç kısmının boyutlarına karşılık gelir. Spiral oluşturmak için çift CTçok küçük olurdu çift A-G, aksine, çok büyük. Yani, DNA'nın ilk ipliğinin azotlu bazı, diğer DNA zincirinin aynı yerinde bulunan bazı belirtir. Bir DNA molekülünde birbirine paralel çift zincirler halinde bulunan nükleotitlerin kesin yazışmalarına denir. tamamlayıcılık (isteğe bağlı). tam üreme veya çoğaltma DNA molekülünün bu özelliği sayesinde genetik bilgi de mümkündür.
DNA'da biyolojik bilgiler tam olarak kopyalanabilecek ve sonraki hücrelere aktarılabilecek şekilde kaydedilir. Hücre bölünmesinden önce, çoğaltma (kendini ikiye katlama ) DNA'sı. Her zincir, partner zincirine tamamlayıcı bir nükleotid dizisi içerdiğinden, aslında aynı genetik bilgiyi taşırlar. Telleri ayırır ve her birini ikinci bir iplik oluşturmak için bir şablon (matris) olarak kullanırsanız, iki yeni özdeş DNA ipliği elde edersiniz. Bir hücrede DNA bu şekilde kopyalanır.


