ما هو مصنوع من الحمض النووي. النوكليوتيدات - ما هو؟ تكوين وتركيب وعدد وتسلسل النيوكليوتيدات في سلسلة DNA يتم تثبيت الحلزون المزدوج للحمض النووي بواسطة الأيونات
الأحماض النووية عبارة عن مواد جزيئية كبيرة تتكون من أحاديات النيوكليوتيدات ، والتي ترتبط ببعضها البعض في سلسلة بوليمر باستخدام روابط فوسفوديستر 3 "، 5" ومعبأة في الخلايا بطريقة معينة.
الأحماض النووية عبارة عن بوليمرات حيوية من نوعين: الحمض النووي الريبي (RNA) وحمض الديوكسي ريبونوكلييك (DNA). يتكون كل بوليمر حيوي من نيوكليوتيدات تختلف في بقايا الكربوهيدرات (ريبوز ، ديوكسيريبوز) وأحد القواعد النيتروجينية (اليوراسيل ، الثايمين). تبعا لذلك ، حصلت الأحماض النووية على اسمها.
هيكل حمض الديوكسي ريبونوكلييك
الأحماض النووية لها هياكل أولية وثانوية وثالثية.
الهيكل الأساسي للحمض النووي
الهيكل الأساسي للحمض النووي هو سلسلة خطية من عديد النوكليوتيدات حيث ترتبط أحاديات النيوكليوتيدات بواسطة روابط فوسفوديستر 3 "، 5". مادة البداية لتجميع سلسلة حمض نووي في خلية هي نيوكليوسيد 5-ثلاثي الفوسفات ، والذي نتيجة لإزالة β و بقايا حمض الفوسفوريك ، يكون قادرًا على ربط ذرة 3'-carbon لنيوكليوسيد آخر . وهكذا ، فإن ذرة الكربون 3 "لواحد من deoxyribose ترتبط تساهميًا بـ 5" ذرة كربون من مادة deoxyribose أخرى عبر بقايا حمض الفوسفوريك وتشكل سلسلة خطية متعددة النوكليوتيد من الحمض النووي. ومن هنا جاء الاسم: روابط فوسفوديستر 3 "، 5". لا تشارك القواعد النيتروجينية في توصيل النيوكليوتيدات لسلسلة واحدة (الشكل 1.).
يؤدي هذا الارتباط ، بين جزيء حمض الفوسفوريك لنيوكليوتيد واحد وكربوهيدرات آخر ، إلى تكوين العمود الفقري للبنتوز والفوسفات من جزيء متعدد النيوكليوتيد ، حيث يتم إضافة القواعد النيتروجينية واحدة تلو الأخرى من الجانب. تسلسلها في سلاسل جزيئات الحمض النووي محدد بدقة لخلايا الكائنات الحية المختلفة ، أي له طابع محدد (قاعدة Chargaff).
سلسلة الحمض النووي الخطية ، التي يعتمد طولها على عدد النيوكليوتيدات المتضمنة في السلسلة ، لها طرفان: أحدهما يسمى الطرف 3 ويحتوي على هيدروكسيل حر ، والآخر ، الطرف 5 ، يحتوي على حمض الفوسفوريك بقايا. الدائرة قطبية ويمكن أن تكون 5 "-> 3" و 3 "-> 5". استثناء هو دائري DNA.
يتكون "النص" الجيني للحمض النووي من "كلمات" شفرية - ثلاثة توائم من النيوكليوتيدات تسمى الكودونات. تسمى أجزاء الحمض النووي التي تحتوي على معلومات حول التركيب الأساسي لجميع أنواع الحمض النووي الريبي الجينات الهيكلية.
تصل سلاسل الحمض النووي متعدد النوى إلى أحجام هائلة ، لذا فهي معبأة بطريقة معينة في الخلية.
دراسة تكوين الحمض النووي ، أنشأ Chargaff (1949) انتظامات مهمة فيما يتعلق بمحتوى قواعد الحمض النووي الفردية. لقد ساعدوا في الكشف عن البنية الثانوية للحمض النووي. تسمى هذه الأنماط قواعد Chargaff. قواعد Chargaff
تنص هذه القواعد على أنه عند بناء الحمض النووي ، يجب مراعاة المراسلات الصارمة (الاقتران) ليس لقواعد البيورين والبيريميدين بشكل عام ، ولكن بشكل خاص مع الثايمين مع الأدينين والسيتوزين مع الجوانين. بناءً على هذه القواعد ، من بين أمور أخرى ، اقترح واتسون وكريك في عام 1953 نموذجًا للبنية الثانوية للحمض النووي ، يُسمى الحلزون المزدوج (الشكل). |

التركيب الثانوي للحمض النووي
التركيب الثانوي للحمض النووي هو حلزون مزدوج ، اقترح نموذجه D. Watson و F. Crick في عام 1953.
المتطلبات الأساسية لإنشاء نموذج DNA
كنتيجة للتحليلات الأولية ، كانت الفكرة أن الحمض النووي من أي أصل يحتوي على جميع النيوكليوتيدات الأربعة بكميات جزيئية متساوية. ومع ذلك ، في الأربعينيات من القرن الماضي ، أظهر E.Chargaff وزملاؤه ، نتيجة لتحليل الحمض النووي المعزول من كائنات مختلفة ، بوضوح أن القواعد النيتروجينية موجودة فيها بنسب كمية مختلفة. وجد Chargaff أنه على الرغم من أن هذه النسب هي نفسها بالنسبة للحمض النووي من جميع خلايا نفس النوع من الكائنات الحية ، يمكن أن يختلف الحمض النووي من أنواع مختلفة بشكل ملحوظ في محتوى بعض النيوكليوتيدات. يشير هذا إلى أن الاختلافات في نسبة القواعد النيتروجينية قد تكون مرتبطة ببعض الكود البيولوجي. على الرغم من أن نسبة قواعد البيورين والبيريميدين الفردية في عينات مختلفة من الحمض النووي لم تكن متماثلة ، عند مقارنة نتائج التحليلات ، تم الكشف عن نمط معين: في جميع العينات ، كان إجمالي كمية البيورينات مساويًا للكمية الإجمالية للبيريميدين ( A + G = T + C) ، كانت كمية الأدينين مساوية لكمية الثايمين (A = T) ، وكمية الجوانين - كمية السيتوزين (G = C). كان الحمض النووي المعزول من خلايا الثدييات أكثر ثراءً بشكل عام في الأدينين والثايمين وأفقر نسبيًا في الجوانين والسيتوزين ، بينما كان الحمض النووي من البكتيريا أكثر ثراءً في الجوانين والسيتوزين وأكثر فقرًا نسبيًا في الأدينين والثيمين. شكلت هذه البيانات جزءًا مهمًا من المادة الواقعية ، والتي تم على أساسها بناء نموذج بنية الحمض النووي لـ Watson-Crick لاحقًا.
ومن المؤشرات الأخرى الهامة غير المباشرة على التركيب المحتمل للحمض النووي ، بيانات ل. باولينج حول بنية جزيئات البروتين. أظهر Pauling أن العديد من التكوينات المستقرة المختلفة لسلسلة الأحماض الأمينية ممكنة في جزيء البروتين. أحد التكوينات الشائعة لسلسلة الببتيد - α-helix - هو هيكل حلزوني منتظم. مع مثل هذا الهيكل ، يمكن تكوين روابط هيدروجينية بين الأحماض الأمينية الموجودة على المنعطفات المجاورة للسلسلة. وصف بولينج التكوين الحلزوني α لسلسلة البولي ببتيد في عام 1950 واقترح أن جزيئات الحمض النووي من المحتمل أيضًا أن يكون لها بنية حلزونية مثبتة بواسطة روابط هيدروجينية.
ومع ذلك ، تم توفير المعلومات الأكثر قيمة حول بنية جزيء الحمض النووي من خلال نتائج تحليل حيود الأشعة السينية. تخضع الأشعة السينية ، التي تمر عبر بلورة DNA ، للحيود ، أي أنها تنحرف في اتجاهات معينة. تعتمد درجة وطبيعة انحراف الأشعة على بنية الجزيئات نفسها. يعطي نمط حيود الأشعة السينية (الشكل 3) للعين ذات الخبرة عددًا من المؤشرات غير المباشرة فيما يتعلق ببنية جزيئات المادة قيد الدراسة. أدى تحليل أنماط حيود الأشعة السينية للحمض النووي إلى استنتاج مفاده أن القواعد النيتروجينية (ذات الشكل المسطح) مكدسة مثل كومة من الصفائح. جعلت أنماط الأشعة السينية من الممكن تحديد ثلاث فترات رئيسية في بنية الحمض النووي البلوري: 0.34 و 2 و 3.4 نانومتر.

نموذج Watson-Crick DNA
بدءًا من بيانات Chargaff التحليلية ، وأشعة Wilkins للأشعة السينية ، وبحث الكيميائي ، الذي قدم معلومات حول المسافات الدقيقة بين الذرات في الجزيء ، وحول الزوايا بين روابط ذرة معينة ، وحول حجم الذرات ، Watson و Crick بدأ في بناء نماذج فيزيائية للمكونات الفردية لجزيء الحمض النووي على نطاق معين. و "ضبطها" مع بعضها البعض بطريقة يتوافق معها النظام الناتج مع البيانات التجريبية المختلفة [تبين] .
حتى في وقت سابق ، كان معروفًا أن النيوكليوتيدات المجاورة في سلسلة DNA مرتبطة بجسور الفوسفوديستر التي تربط ذرة 5'-carbon من deoxyribose لنيوكليوتيد واحد بذرة 3'-carbon من deoxyribose للنيوكليوتيد التالي. لم يكن لدى واطسون وكريك أدنى شك في أن فترة 0.34 نانومتر تقابل المسافة بين النيوكليوتيدات المتتالية في حبلا DNA. علاوة على ذلك ، يمكن افتراض أن فترة 2 نانومتر تتوافق مع سمك السلسلة. ومن أجل شرح البنية الحقيقية التي تتوافق مع فترة 3.4 نانومتر ، افترض واتسون وكريك ، وكذلك بولينج سابقًا ، أن السلسلة ملتوية على شكل حلزوني (أو بشكل أكثر دقة ، تشكل لولبًا ، منذ بالمعنى الدقيق للكلمة ، يتم الحصول على الكلمة عندما تشكل المنعطفات سطحًا مخروطيًا وليس أسطوانيًا في الفضاء). بعد ذلك ، سوف تتوافق فترة 3.4 نانومتر مع المسافة بين الدورات المتتالية لهذا اللولب. يمكن أن يكون هذا اللولب كثيفًا جدًا أو ممتدًا إلى حد ما ، أي يمكن أن يكون المنعطفات مسطحة أو شديدة الانحدار. نظرًا لأن فترة 3.4 نانومتر هي بالضبط 10 أضعاف المسافة بين النيوكليوتيدات المتتالية (0.34 نانومتر) ، فمن الواضح أن كل دورة كاملة للحلزون تحتوي على 10 نيوكليوتيدات. من هذه البيانات ، تمكن واطسون وكريك من حساب كثافة سلسلة متعددة النوكليوتيد الملتوية في حلزون بقطر 2 نانومتر ، بمسافة بين الدورات تساوي 3.4 نانومتر. اتضح أن مثل هذا الخيط سيكون له نصف كثافة الكثافة الفعلية للحمض النووي ، والتي كانت معروفة بالفعل. كان علي أن أفترض أن جزيء الحمض النووي يتكون من سلسلتين - أنه حلزون مزدوج من النيوكليوتيدات.
كانت المهمة التالية ، بالطبع ، لتوضيح العلاقة المكانية بين الخيوط التي تشكل اللولب المزدوج. بعد تجربة عدد من المتغيرات لترتيب السلاسل على نموذجهم المادي ، وجد Watson and Crick أن الأنسب لجميع البيانات المتاحة هو واحد حيث يذهب حلزون متعدد النوكليوتيد في اتجاهين متعاكسين ؛ في هذه الحالة ، تشكل السلاسل المكونة من مخلفات السكر والفوسفات سطح حلزون مزدوج ، وتوجد بداخلها البيورينات والبيريميدين. القواعد الموجودة مقابل بعضها البعض ، والتي تنتمي إلى سلسلتين ، متصلة في أزواج بواسطة روابط هيدروجينية ؛ هذه الروابط الهيدروجينية هي التي تربط السلاسل معًا ، وبالتالي تحدد التكوين العام للجزيء.
يمكن اعتبار الحلزون المزدوج للحمض النووي بمثابة سلم حبل حلزوني ، مع بقاء الدرجات أفقية. ثم يتطابق حبال طوليان مع سلاسل من مخلفات السكر والفوسفات ، وسوف تتوافق القضبان المتقاطعة مع أزواج من القواعد النيتروجينية المتصلة بواسطة روابط هيدروجينية.
نتيجة لمزيد من الدراسة للنماذج المحتملة ، توصل واطسون وكريك إلى استنتاج مفاده أن كل "عارضة" يجب أن تتكون من بيورين واحد وبيريميدين ؛ في فترة 2 نانومتر (المقابلة لقطر اللولب المزدوج) ، لن يكون هناك مساحة كافية لاثنين من البيورينات ، ولا يمكن أن يكون البيريميدين قريبين بما يكفي لتكوين روابط هيدروجينية مناسبة. أظهرت دراسة متعمقة للنموذج التفصيلي أن الأدينين والسيتوزين ، اللذان يشكلان مزيجًا من الحجم الصحيح ، لا يزال من غير الممكن ترتيبهما بطريقة تتشكل الروابط الهيدروجينية بينهما. كما أجبرت تقارير مماثلة أيضًا على استبعاد تركيبة الغوانين والثيمين ، في حين تم العثور على توليفات الأدينين - الثايمين والجوانين - السيتوزين لتكون مقبولة تمامًا. طبيعة الروابط الهيدروجينية هي أن أزواج الأدينين مع الثايمين ، وأزواج الجوانين مع السيتوزين. أتاح مفهوم الاقتران الأساسي المحدد شرح "قاعدة Chargaff" ، والتي بموجبها تكون كمية الأدينين في أي جزيء DNA مساوية دائمًا لمحتوى الثايمين ، وتكون كمية الجوانين دائمًا مساوية لكمية السيتوزين . يتكون رابطان هيدروجين بين الأدينين والثايمين ، وثلاثة بين الجوانين والسيتوزين. بسبب هذه الخصوصية في تكوين روابط هيدروجينية ضد كل أدينين في سلسلة واحدة ، يكون الثايمين في الآخر ؛ بنفس الطريقة ، يمكن وضع السيتوزين فقط ضد كل جوانين. وبالتالي ، فإن السلاسل مكملة لبعضها البعض ، أي أن تسلسل النيوكليوتيدات في إحدى السلاسل يحدد بشكل فريد تسلسلها في الأخرى. تعمل السلسلتان في اتجاهين متعاكسين وتكون مجموعاتهما الفوسفاتية على طرفي نقيض من الحلزون المزدوج.
نتيجة لأبحاثهم ، اقترح واطسون وكريك في عام 1953 نموذجًا لهيكل جزيء الحمض النووي (الشكل 3) ، والذي يظل وثيق الصلة بالحاضر. وفقًا للنموذج ، يتكون جزيء الحمض النووي من سلسلتين متكاملتين من عديد النوكليوتيدات. كل خيط DNA هو بولي نيوكليوتيد يتكون من عدة عشرات الآلاف من النيوكليوتيدات. في ذلك ، تشكل النيوكليوتيدات المجاورة العمود الفقري المنتظم للبنتوز والفوسفات بسبب مزيج من بقايا حمض الفوسفوريك و deoxyribose بواسطة رابطة تساهمية قوية. يتم ترتيب القواعد النيتروجينية لسلسلة واحدة من عديد النوكليوتيدات بترتيب محدد بدقة مقابل القواعد النيتروجينية للسلسلة الأخرى. تناوب القواعد النيتروجينية في سلسلة عديد النوكليوتيد غير منتظم.
ترتيب القواعد النيتروجينية في سلسلة الحمض النووي تكميلي (من "الإضافة" اليونانية ") ، أي ضد الأدينين (A) هو دائمًا الثايمين (T) ، وضد الجوانين (G) - فقط السيتوزين (C). يفسر ذلك حقيقة أن A و T ، وكذلك G و C ، يتوافقان تمامًا مع بعضهما البعض ، أي يكمل كل منهما الآخر. يتم إعطاء هذا التطابق من خلال التركيب الكيميائي للقواعد ، والذي يسمح بتكوين روابط هيدروجينية في زوج من البيورين والبيريميدين. بين A و T هناك رابطان ، بين G و C - ثلاثة. توفر هذه الروابط استقرارًا جزئيًا لجزيء الحمض النووي في الفضاء. يتناسب استقرار اللولب المزدوج بشكل مباشر مع عدد روابط G≡C ، والتي تكون أكثر استقرارًا من روابط A = T.
إن التسلسل المعروف للنيوكليوتيدات في أحد خيوط الدنا يجعل من الممكن ، من خلال مبدأ التكامل ، إنشاء نيوكليوتيدات خيط آخر.
بالإضافة إلى ذلك ، فقد ثبت أن القواعد النيتروجينية ذات البنية العطرية توجد واحدة فوق الأخرى في محلول مائي ، وتشكل ، كما كانت ، كومة من العملات المعدنية. هذه العملية لتشكيل أكوام من جزيئات عضويةيسمى التكديس. سلاسل عديد النوكليوتيد لجزيء DNA لنموذج Watson-Crick المدروس لها حالة كيميائية فيزيائية مماثلة ، حيث يتم ترتيب قواعدها النيتروجينية في شكل كومة من العملات المعدنية ، بين المستويات التي تحدث فيها تفاعلات van der Waals (تفاعلات التراص).
الروابط الهيدروجينية بين القواعد التكميلية (أفقيًا) والتفاعل المتراكم بين المستويات الأساسية في سلسلة متعددة النوكليوتيد بسبب قوى فان دير فالس (عموديًا) تزود جزيء الحمض النووي بثبات إضافي في الفضاء.
يتم قلب العمود الفقري للسكر والفوسفات لكلا السلسلتين إلى الخارج ، والقواعد للداخل ، تجاه بعضها البعض. اتجاه الخيوط في الحمض النووي غير متوازي (أحدهما له اتجاه 5 "-> 3" ، والآخر - 3 "-> 5" ، أي أن الطرف 3 "من خيط واحد يقع مقابل 5" نهاية من جهة أخرى.). تشكل السلاسل لولبًا صحيحًا مع محور مشترك. دورة واحدة من اللولب هي 10 نيوكليوتيدات ، وحجم المنعطف 3.4 نانومتر ، وارتفاع كل نيوكليوتيد 0.34 نانومتر ، وقطر اللولب 2.0 نانومتر. نتيجة لدوران خيط واحد حول الآخر ، يتشكل أخدود رئيسي (قطره حوالي 20) وأخدود صغير (حوالي 12 Å) في الحلزون المزدوج للحمض النووي. هذا الشكل من الحلزون المزدوج Watson-Crick سمي لاحقًا بالنموذج B. في الخلايا ، يوجد الحمض النووي عادةً في الشكل B ، وهو الأكثر استقرارًا.
وظائف الحمض النووي
شرح النموذج المقترح العديد من الخصائص البيولوجية للحمض النووي الريبي منقوص الأكسجين ، بما في ذلك تخزين المعلومات الجينية وتنوع الجينات ، التي توفرها مجموعة متنوعة من التوليفات المتتالية من 4 نيوكليوتيدات وحقيقة وجود رمز وراثي ، والقدرة على التكاثر الذاتي ونقل المعلومات الجينية ، التي توفرها عملية النسخ المتماثل ، وتنفيذ المعلومات الجينية في شكل بروتينات ، وكذلك أي مركبات أخرى تكونت بمساعدة بروتينات الإنزيم.
الوظائف الأساسية للحمض النووي.
- الحمض النووي هو الناقل للمعلومات الجينية ، والتي يتم ضمانها من خلال حقيقة وجود الشفرة الجينية.
- التكاثر والمعلومات الوراثية المنقولة في أجيال من الخلايا والكائنات الحية. يتم توفير هذه الوظيفة من خلال عملية النسخ المتماثل.
- تنفيذ المعلومات الجينية على شكل بروتينات وأي مركبات أخرى تكونت بمساعدة بروتينات الإنزيم. يتم توفير هذه الوظيفة من خلال عمليات النسخ والترجمة.

أشكال تنظيم الحمض النووي مزدوج الشريطة
يمكن أن يشكل الحمض النووي عدة أنواع من الحلزون المزدوج (الشكل 4). حاليًا ، هناك ستة أشكال معروفة بالفعل (من A إلى E و Z-form).
تعتمد الأشكال الهيكلية للحمض النووي ، كما حددتها روزاليند فرانكلين ، على تشبع جزيء الحمض النووي بالماء. في الدراسات التي أجريت على ألياف الحمض النووي باستخدام تحليل حيود الأشعة السينية ، تبين أن نمط حيود الأشعة السينية يعتمد بشكل جذري على الرطوبة النسبية ، ودرجة تشبع الماء لهذه الألياف التي تجري التجربة. إذا كانت الألياف مشبعة بدرجة كافية بالماء ، فسيتم الحصول على صورة شعاعية واحدة. عند التجفيف ، ظهر نمط مختلف تمامًا للأشعة السينية ، يختلف تمامًا عن نمط الأشعة السينية للألياف عالية الرطوبة.
يسمى جزيء الحمض النووي عالي الرطوبة شكل ب. في ظل الظروف الفسيولوجية (انخفاض تركيز الملح ، ودرجة عالية من الماء) ، فإن النوع الهيكلي السائد للحمض النووي هو الشكل B (الشكل الرئيسي للحمض النووي مزدوج الشريطة هو نموذج Watson-Crick). تبلغ درجة اللولب لمثل هذا الجزيء 3.4 نانومتر. يوجد 10 أزواج تكميلية لكل دور على شكل أكوام ملتوية من "العملات المعدنية" - قواعد نيتروجينية. يتم تثبيت الأكوام معًا بواسطة روابط هيدروجينية بين "عملتين" متقابلتين من الأكوام ، و "ملفوفة" بشريطين من العمود الفقري للفوسفودايستر ملفوفًا في لولب أيمن. مستويات القواعد النيتروجينية متعامدة على محور اللولب. يتم تدوير الأزواج التكميلية المجاورة بالنسبة لبعضها البعض بمقدار 36 درجة. يبلغ قطر اللولب 20 درجة مئوية ، مع احتلال نيوكليوتيد البيورين 12 درجة مئوية بينما يشغل نيوكليوتيد البيريميدين 8 درجات مئوية.
يسمى جزيء الحمض النووي ذو الرطوبة المنخفضة بالشكل A. يتكون النموذج A في ظل ظروف أقل نسبة ترطيب ومحتوى أعلى من أيونات الصوديوم أو البوتاسيوم. يحتوي هذا التشكل الأوسع لليد اليمنى على 11 زوجًا أساسيًا لكل منعطف. تتمتع مستويات القواعد النيتروجينية بميل أقوى لمحور اللولب ، فهي تنحرف عن المحور الطبيعي إلى محور اللولب بمقدار 20 درجة. هذا يعني وجود فراغ داخلي بقطر 5. المسافة بين النيوكليوتيدات المجاورة هي 0.23 نانومتر ، وطول الملف 2.5 نانومتر ، وقطر اللولب 2.3 نانومتر.
في البداية ، كان يُعتقد أن الشكل A من الحمض النووي أقل أهمية. ومع ذلك ، اتضح لاحقًا أن الشكل A من الحمض النووي ، وكذلك الشكل B ، لهما أهمية بيولوجية كبيرة. يحتوي حلزون RNA-DNA في مجمع البذور النموذجية على الشكل A ، بالإضافة إلى هياكل حلزون RNA-RNA و RNA (لا تسمح مجموعة 2'-hydroxyl من الريبوز لجزيئات RNA بتكوين الشكل B) . تم العثور على شكل A من الحمض النووي في الجراثيم. لقد ثبت أن الشكل A من الحمض النووي أكثر مقاومة للأشعة فوق البنفسجية بعشر مرات من الشكل B.
يتم استدعاء النموذج A والنموذج B الأشكال المتعارف عليهاالحمض النووي.
أشكال C-Eأيضًا في اليد اليمنى ، لا يمكن ملاحظة تكوينها إلا في تجارب خاصة ، ويبدو أنها غير موجودة في الجسم الحي. يحتوي الشكل C من الحمض النووي على بنية مشابهة لـ B-DNA. عدد أزواج القاعدة في كل دورة هو 9.33 ، وطول اللولب 3.1 نانومتر. تميل أزواج القاعدة بزاوية 8 درجات بالنسبة للموضع العمودي على المحور. الأخاديد قريبة في الحجم من أخاديد B-DNA. في هذه الحالة ، يكون الأخدود الرئيسي أصغر إلى حد ما ، ويكون الأخدود الصغير أعمق. يمكن للعديد من النيوكليوتيدات الطبيعية والاصطناعية أن تنتقل إلى الشكل C.
| الجدول 1. خصائص بعض أنواع هياكل الحمض النووي | |||
| نوع حلزوني | أ | ب | ض |
| الملعب الحلزوني | 0.32 نانومتر | 3.38 نانومتر | 4.46 نانومتر |
| تطور لولبي | حق | حق | اليسار |
| عدد أزواج القاعدة لكل دور | 11 | 10 | 12 |
| المسافة بين الطائرات الأساسية | 0.256 نانومتر | 0.338 نانومتر | 0.371 نانومتر |
| تشكيل الرابطة الجليكوسيدية | مضاد | مضاد | مضاد- C سين- G |
| شكل حلقة فورانوز | C3 "-يندو | C2 "-endo | C3 "-يندو-ج C2 "-endo-C |
| عرض الأخدود ، صغير / كبير | 1.11 / 0.22 نانومتر | 0.57 / 1.17 نانومتر | 0.2 / 0.88 نانومتر |
| عمق الأخدود ، صغير / كبير | 0.26 / 1.30 نانومتر | 0.82 / 0.85 نانومتر | 1.38 / 0.37 نانومتر |
| قطر حلزوني | 2.3 نانومتر | 2.0 نانومتر | 1.8 نانومتر |
العناصر الهيكلية للحمض النووي
(هياكل الحمض النووي غير الكنسي)
تشتمل العناصر الهيكلية للحمض النووي على تراكيب غير عادية مقيدة ببعض التسلسلات الخاصة:
|

شكل Z من DNAتم اكتشافه في عام 1979 أثناء دراسة hexanucleotide d (CG) 3 -. تم افتتاحه من قبل الأستاذ في معهد ماساتشوستس للتكنولوجيا ألكسندر ريتش وموظفيه. أصبح الشكل Z أحد أهم العناصر الهيكلية للحمض النووي بسبب حقيقة أن تكوينه لوحظ في مناطق الحمض النووي حيث تتناوب البيورينات مع البيريميدين (على سبيل المثال ، 5'-HCHCHC-3 ') ، أو في التكرارات 5' -CHCHCH-3 'تحتوي على سيتوزين ميثيل. كان الشرط الأساسي لتكوين Z-DNA وتثبيته هو وجود نيوكليوتيدات البيورين في التشكل المتزامن ، بالتناوب مع قواعد بيريميدين في مضاد التشكل.
توجد جزيئات الحمض النووي الطبيعية في الغالب في الشكل B الصحيح ما لم تحتوي على تسلسلات مثل (CG) n. ومع ذلك ، إذا كانت هذه التسلسلات جزءًا من الحمض النووي ، فإن هذه المناطق ، عندما تكون القوة الأيونية للمحلول أو الكاتيونات التي تحيد الشحنة السالبة على العمود الفقري للفوسفوديستر ، يمكن أن تتغير إلى شكل Z ، بينما تظل مناطق الحمض النووي الأخرى في السلسلة في النموذج الكلاسيكي ب. تشير إمكانية حدوث مثل هذا الانتقال إلى أن الخيطين في الحلزون المزدوج للحمض النووي في حالة ديناميكية ويمكنهما الاسترخاء بالنسبة لبعضهما البعض ، ويمر من الشكل الأيمن إلى الشكل الأيسر والعكس صحيح. لم يتم بعد فهم العواقب البيولوجية لهذه القابلية ، والتي تسمح بالتحولات التوافقية لبنية الحمض النووي. يُعتقد أن مناطق Z-DNA تلعب دورًا في تنظيم التعبير عن جينات معينة وتشارك في إعادة التركيب الجيني.
الشكل Z للحمض النووي هو حلزون مزدوج أعسر ، حيث يكون العمود الفقري للفوسفوديستر متعرجًا على طول محور الجزيء. ومن هنا جاء اسم الجزيء (zigzag) -DNA. Z-DNA هو الأقل تواءًا (12 زوجًا قاعديًا في كل منعطف) والأرق في الطبيعة. المسافة بين النيوكليوتيدات المجاورة 0.38 نانومتر ، وطول الملف 4.56 نانومتر ، وقطر Z-DNA 1.8 نانومتر. علاوة على ذلك، مظهر خارجييتميز جزيء الحمض النووي هذا بوجود أخدود واحد.
تم العثور على شكل Z من الحمض النووي في الخلايا بدائية النواة وحقيقية النواة. حتى الآن ، تم الحصول على أجسام مضادة يمكنها التمييز بين الشكل Z والشكل B للحمض النووي. ترتبط هذه الأجسام المضادة بمناطق معينة من الكروموسومات العملاقة لخلايا الغدة اللعابية (دكتور ميلانوجاستر). من السهل متابعة تفاعل الارتباط بسبب التركيب غير المعتاد لهذه الكروموسومات ، حيث تتناقض المناطق الأكثر كثافة (الأقراص) مع المناطق الأقل كثافة (interdisks). تقع مناطق Z-DNA في الشرائح. ويترتب على ذلك أن الشكل Z موجود بالفعل في ظروف طبيعية ، على الرغم من أن أحجام الأقسام الفردية للصيغة Z غير معروفة بعد.

(المتحولون) - التسلسلات الأساسية الأكثر شهرة وتكرارًا في الحمض النووي. المتماثل هو كلمة أو عبارة تقرأ من اليسار إلى اليمين والعكس صحيح بنفس الطريقة. أمثلة على هذه الكلمات أو العبارات هي: HUT ، و COSSACK ، و FLOOD ، و ROSE FALLED ON AZOR'S PAWS. عند تطبيقه على أقسام الحمض النووي ، فإن هذا المصطلح (المتناظر) يعني نفس تناوب النيوكليوتيدات على طول السلسلة من اليمين إلى اليسار ومن اليسار إلى اليمين (مثل الأحرف في كلمة "كوخ" ، إلخ).
يتميز التناظر المتناظر بوجود تكرارات مقلوبة لتسلسلات أساسية لها تناظر من الدرجة الثانية فيما يتعلق بسلاسل DNA. مثل هذه التسلسلات ، لأسباب واضحة ، مكملة ذاتيًا وتميل إلى تكوين هياكل دبوس الشعر أو صليبية الشكل (الشكل). تساعد دبابيس الشعر البروتينات المنظمة في التعرف على المكان الذي يتم فيه نسخ النص الجيني للحمض النووي للكروموسوم.
في الحالات التي يكون فيها التكرار المقلوب موجودًا في نفس شريط الحمض النووي ، يُطلق على هذا التسلسل تكرار المرآة. لا تتمتع عمليات تكرار المرآة بخصائص تكاملية ذاتية ، وبالتالي فهي غير قادرة على تكوين هياكل دبوس الشعر أو الصليبية. توجد تسلسلات من هذا النوع في جميع جزيئات الحمض النووي الكبيرة تقريبًا ويمكن أن تتراوح من بضعة أزواج أساسية إلى عدة آلاف من أزواج القواعد.
لم يتم إثبات وجود المتجانسات في شكل هياكل صليبية في الخلايا حقيقية النواة ، على الرغم من وجود عدد من الهياكل الصليبية في الجسم الحي في خلايا الإشريكية القولونية. إن وجود المتواليات الذاتية التكميلية في الحمض النووي الريبي أو الحمض النووي أحادي السلسلة هو السبب الرئيسي لطي السلسلة النووية في المحاليل إلى بنية مكانية معينة ، تتميز بتكوين العديد من "دبابيس الشعر".

شكل H من الحمض النووي- هذا حلزون يتكون من ثلاثة خيوط من الحمض النووي - الحلزون الثلاثي للحمض النووي. إنه مجمع من الحلزون المزدوج Watson-Crick مع خيط DNA أحادي السلسلة الثالث ، والذي يتناسب مع أخدوده الكبير ، مع تكوين ما يسمى بزوج Hoogsteen.
يحدث تكوين مثل هذا الثلاثي نتيجة لإضافة الحلزون المزدوج للحمض النووي بحيث يظل نصف قسمه على شكل حلزون مزدوج ، ويتم فصل النصف الثاني. في هذه الحالة ، تشكل إحدى الحلزونات المنفصلة هيكلًا جديدًا مع النصف الأول من اللولب المزدوج - وهو حلزون ثلاثي ، والثاني يتضح أنه غير منظم ، على شكل قسم أحادي الشعيرة. من سمات هذا التحول الهيكلي الاعتماد الحاد على الرقم الهيدروجيني للوسط ، حيث تعمل البروتونات على استقرار الهيكل الجديد. بسبب هذه الميزة ، تم تسمية الهيكل الجديد بالشكل H للحمض النووي ، والذي تم العثور على تكوينه في البلازميدات فائقة الالتفاف التي تحتوي على مناطق homopurine-homopyrimidine ، والتي هي تكرار المرآة.
في مزيد من الدراسات ، تم تحديد إمكانية الانتقال الهيكلي لبعض البولينيوكليوتيدات مزدوجة الشريطة homopurine-homopyrimidine من خلال تكوين بنية ثلاثية الشرائط تحتوي على:
- homopurine واحد واثنين من خيوط homopyrimidine ( ثلاثي Py-Pu-Py) [تفاعل هوغستين].
الكتل المكونة لثلاثي Py-Pu-Py هي ثلاثية متشابهة CGC + وثلاثيات TAT. يتطلب استقرار الثلاثي بروتون ثلاثي CGC + ، لذلك تعتمد هذه الثلاثيات على الرقم الهيدروجيني للمحلول.
- واحد homopyrimidine واثنين من خيوط homopurine ( ثلاثي Py-Pu-Pu) [تفاعل Hoogsteen العكسي].
الكتل المكونة لثلاثي Py-Pu-Pu هي الثلاثيات المتشابهة CGG و TAA. الخاصية الأساسية لثلاثي Py-Pu-Pu هي اعتماد ثباتها على وجود أيونات مشحونة مضاعفة ، وهناك حاجة إلى أيونات مختلفة لتثبيت الثلاثيات من متواليات مختلفة. نظرًا لأن تكوين ثلاثي Py-Pu-Pu لا يتطلب بروتونات النيوكليوتيدات المكونة لها ، يمكن أن توجد هذه الثلاثيات عند درجة الحموضة المحايدة.
ملحوظة: يتم تفسير تفاعل Hoogsteen المباشر والعكسي من خلال تناظر 1-methylthymine: يؤدي دوران 180 درجة إلى حقيقة أن مكان ذرة O4 تحتلها ذرة O2 ، بينما يتم الحفاظ على نظام الروابط الهيدروجينية.
هناك نوعان من اللولب الثلاثي:
- اللولب الثلاثي المتوازي الذي يكون فيه قطبية الخيط الثالث هو نفسه قطبية السلسلة المثلية في Watson-Crick doublelex
- اللولب الثلاثي المضاد الموازي ، حيث تكون أقطاب السلاسل الثالثة وسلسلة homopurine متعاكسة.

رباعي G- 4-الذين تقطعت بهم السبل الحمض النووي. يتشكل مثل هذا الهيكل إذا كان هناك أربعة جوانين ، والتي تشكل ما يسمى G-quadruplex - رقصة مستديرة من أربعة جوانين.
تم الحصول على الإشارات الأولى لإمكانية تكوين مثل هذه الهياكل قبل فترة طويلة من العمل الخارق لـ Watson and Crick - في وقت مبكر من عام 1910. ثم اكتشف الكيميائي الألماني إيفار بانغ أن أحد مكونات الحمض النووي - حمض جوانوسيك - يشكل مواد هلامية بتركيزات عالية ، في حين أن المكونات الأخرى للحمض النووي لا تمتلك هذه الخاصية.
في عام 1962 ، باستخدام طريقة حيود الأشعة السينية ، كان من الممكن تحديد بنية الخلية لهذا الهلام. اتضح أنها تتكون من أربعة بقايا جوانين ، وتربط بعضها البعض في دائرة وتشكل مربعًا مميزًا. في الوسط ، يتم دعم الرابطة بواسطة أيون معدني (Na ، K ، Mg). يمكن تكوين نفس الهياكل في الحمض النووي إذا كان يحتوي على الكثير من الجوانين. يتم تكديس هذه المربعات المسطحة (G-quartets) لتشكيل هياكل كثيفة ومستقرة إلى حد ما (G-quadruplexes).
يمكن نسج أربعة خيوط منفصلة من الحمض النووي في مجمعات من أربع خيوط ، لكن هذا استثناء. في كثير من الأحيان ، يتم ربط خيط واحد من الحمض النووي ببساطة في عقدة ، مما يؤدي إلى تكوين ثخانات مميزة (على سبيل المثال ، في نهايات الكروموسومات) ، أو يشكل الحمض النووي المزدوج الشريطة رباعيًا محليًا في بعض المواقع الغنية بالجوانين.
أكثر ما تم دراسته هو وجود الانعكاسات الرباعية في نهايات الكروموسومات - على التيلوميرات وفي المحفزات الورمية. ومع ذلك ، لا يزال الفهم الكامل لتوطين مثل هذا الحمض النووي في الكروموسومات البشرية غير معروف.
كل هذه الهياكل غير العادية للحمض النووي في الشكل الخطي غير مستقرة مقارنة بالشكل ب من الحمض النووي. ومع ذلك ، غالبًا ما يوجد الحمض النووي في شكل حلقة من التوتر الطوبولوجي عندما يحتوي على ما يعرف باسم الالتفاف الفائق. في ظل هذه الظروف ، تتشكل هياكل الحمض النووي غير المتعارف عليها بسهولة: أشكال Z ، و "الصلبان" و "دبابيس الشعر" ، وأشكال H ، و ganine quadruplexes ، و i-motif.
- شكل فائق الالتواء - يُلاحظ عند إطلاقه من نواة الخلية دون الإضرار بالعمود الفقري للبنتوز والفوسفات. لها شكل حلقات مغلقة مفرطة التواء. في حالة الالتواء الفائق ، يكون اللولب المزدوج للحمض النووي "ملتويًا على نفسه" مرة واحدة على الأقل ، أي أنه يحتوي على ملف فائق واحد على الأقل (يأخذ شكل الشكل الثامن).
- حالة استرخاء من الحمض النووي - لوحظ بقطع واحد (كسر حبلا واحد). في هذه الحالة ، تختفي الملفات الفائقة ويأخذ الحمض النووي شكل حلقة مغلقة.
- يُلاحظ الشكل الخطي للحمض النووي عندما يتم كسر خيطين من اللولب المزدوج.
البنية الثلاثية للحمض النووي
البنية الثلاثية للحمض النوويتتشكل نتيجة التواء إضافي في الفضاء لجزيء مزدوج الشريطة - الالتواء الفائق. يتم إجراء الالتواء الفائق لجزيء الحمض النووي في الخلايا حقيقية النواة ، على عكس بدائيات النوى ، في شكل مجمعات بها بروتينات.
يقع كل DNA حقيقيات النوى تقريبًا في كروموسومات النوى ، وتوجد كمية صغيرة منه في الميتوكوندريا والنباتات والبلاستيدات. المادة الرئيسية لكروموسومات الخلايا حقيقية النواة (بما في ذلك الكروموسومات البشرية) هي الكروماتين ، وتتكون من بروتينات DNA مزدوجة الشريطة وهيستون وبروتينات غير هيستون.
بروتينات هيستون من الكروماتين
الهيستونات عبارة عن بروتينات بسيطة تشكل ما يصل إلى 50٪ من الكروماتين. في جميع الخلايا المدروسة من الحيوانات والنباتات ، تم العثور على خمس فئات رئيسية من الهيستونات: H1 ، H2A ، H2B ، H3 ، H4 ، تختلف في الحجم ، وتركيب الأحماض الأمينية والشحنة (موجبة دائمًا).
يتكون هيستون Mammalian H1 من سلسلة بولي ببتيد واحدة تحتوي على ما يقرب من 215 حمضًا أمينيًا ؛ تختلف أحجام الهستونات الأخرى من 100 إلى 135 من الأحماض الأمينية. كل منهم حلزوني وملفوف في كرة قطرها حوالي 2.5 نانومتر ، تحتوي على كمية كبيرة بشكل غير عادي من الأحماض الأمينية موجبة الشحنة ليسين وأرجينين. يمكن أسيتات الهستونات ، ميثلة ، فسفرة ، بولي (ADP) - ريبوسيليتيد ، ويمكن ربط الهيستونات H2A و H2B تساهميًا مع يوبيكويتين. ما هو دور هذه التعديلات في تكوين هيكل وأداء الوظائف بواسطة الهيستونات لم يتم توضيحه بالكامل بعد. من المفترض أن هذه هي قدرتها على التفاعل مع الحمض النووي وتوفير إحدى الآليات لتنظيم عمل الجينات.
تتفاعل الهيستونات مع الحمض النووي بشكل أساسي من خلال الروابط الأيونية (جسور الملح) التي تتكون بين مجموعات الفوسفات سالبة الشحنة في الحمض النووي وبقايا الهيستونات ذات الشحنة الموجبة والليسين والأرجينين.
بروتينات الكروماتين غير هيستون
تتنوع البروتينات غير الهيستون بشكل كبير ، على عكس الهستونات. تم عزل ما يصل إلى 590 جزءًا مختلفًا من البروتينات غير الهيستونية المرتبطة بالحمض النووي. وتسمى أيضًا البروتينات الحمضية ، حيث تسود الأحماض الأمينية الحمضية في بنيتها (وهي بوليانيون). يرتبط التنظيم المحدد لنشاط الكروماتين بمجموعة متنوعة من البروتينات غير الهيستون. على سبيل المثال ، يمكن أن ترتبط الإنزيمات الضرورية لتكرار الحمض النووي والتعبير عنه بالكروماتين بشكل عابر. البروتينات الأخرى ، كما يقول المتورطون في عمليات تنظيمية مختلفة ، ترتبط بالحمض النووي فقط في أنسجة معينة أو في مراحل معينة من التمايز. كل بروتين مكمل لسلسلة معينة من نيوكليوتيدات الدنا (موقع DNA). تشمل هذه المجموعة:
- عائلة من بروتينات أصابع الزنك الخاصة بالموقع. يتعرف كل "إصبع من الزنك" على موقع محدد يتكون من 5 أزواج من النيوكليوتيدات.
- عائلة من البروتينات الخاصة بالموقع - المتجانسة. ولجزء من هذا البروتين المتصل بالحمض النووي بنية "حلزونية دورانية".
- البروتينات عالية الحركة (بروتينات HMG - من بروتينات الهلام الإنجليزية عالية الحركة) هي مجموعة من البروتينات الهيكلية والتنظيمية التي ترتبط باستمرار بالكروماتين. يقل وزنها الجزيئي عن 30 كيلو دالتون وتتميز بمحتوى عالٍ من الأحماض الأمينية المشحونة. نظرًا لوزنها الجزيئي المنخفض ، فإن بروتينات HMG شديدة الحركة أثناء الرحلان الكهربائي للهلام متعدد الأكريلاميد.
- إنزيمات النسخ والنسخ والإصلاح.
بمشاركة البروتينات الهيكلية والتنظيمية والإنزيمات المشاركة في تخليق DNA و RNA ، يتم تحويل الخيط النووي إلى مركب عالي الكثافة من البروتينات والحمض النووي. الهيكل الناتج أقصر بـ 10000 مرة من جزيء الحمض النووي الأصلي.

الكروماتينية
الكروماتين هو مركب من البروتينات مع DNA و مواد غير عضوية. معظم الكروماتين غير نشط. يحتوي على DNA مكثف ومكثف. هذا هو الهيتروكروماتين. هناك كروماتين تأسيسي غير نشط وراثيًا (DNA ساتلي) يتكون من مناطق غير معبر عنها ، واختياري - غير نشط في عدد من الأجيال ، ولكن في ظل ظروف معينة قادرة على التعبير.
الكروماتين النشط (كروماتين حقيقي) غير مكثف ، أي معبأة بإحكام أقل. في الخلايا المختلفة ، يتراوح محتواها من 2 إلى 11٪. في خلايا الدماغ هي الأكثر - 10-11 ٪ ، في خلايا الكبد - 3-4 والكلى - 2-3 ٪. هناك نسخ نشط من كروماتين حقيقي. في الوقت نفسه ، يتيح تنظيمها الهيكلي إمكانية استخدام نفس المعلومات الجينية للحمض النووي المتأصلة في نوع معين من الكائنات الحية بطرق مختلفة في الخلايا المتخصصة.
في المجهر الإلكتروني ، تشبه صورة الكروماتين الخرز: سماكات كروية يبلغ حجمها حوالي 10 نانومتر ، مفصولة بجسور خيطية. تسمى هذه التكثيفات الكروية nucleosomes. النواة هي الوحدة الهيكلية للكروماتين. يحتوي كل نيوكليوسوم على جرح مقطع DNA فائق الالتفاف بطول 146 نقطة أساس ليشكل 1.75 لفة يسار لكل نواة نيوكليوسوم. النواة النووية عبارة عن أوكتامر هيستون يتكون من H2A و H2B و H3 و H4 ، وهما جزيءان من كل نوع (الشكل 9) ، والذي يشبه قرص يبلغ قطره 11 نانومتر وسمكه 5.7 نانومتر. الهستون الخامس ، H1 ، ليس جزءًا من النواة النووية ولا يشارك في عملية لف الحمض النووي حول أوكتامر هيستون. إنه يتصل بالحمض النووي عند النقاط التي يدخل فيها اللولب المزدوج ويخرج من النواة النووية. هذه أقسام (رابط) بين النوى من الحمض النووي ، ويختلف طولها اعتمادًا على نوع الخلية من 40 إلى 50 زوجًا من النوكليوتيدات. نتيجة لذلك ، يختلف طول جزء الحمض النووي الذي يعد جزءًا من النيوكليوسومات أيضًا (من 186 إلى 196 زوجًا من النيوكليوتيدات).
يحتوي النوكليوسوم على حوالي 90٪ من الحمض النووي ، والباقي منه هو الرابط. يُعتقد أن النيوكليوسومات عبارة عن أجزاء من الكروماتين "الصامت" ، بينما يكون الرابط نشطًا. ومع ذلك ، يمكن أن تتكشف النيوكليوسومات وتصبح خطية. النيوكليوسومات غير المطوية هي كروماتين نشط بالفعل. يوضح هذا بوضوح اعتماد الوظيفة على الهيكل. يمكن افتراض أنه كلما زاد عدد الكروماتين في تكوين النيوكليوزومات الكروية ، كان نشاطه أقل. من الواضح ، في الخلايا المختلفة ، ترتبط النسبة غير المتكافئة من الكروماتين الساكن بعدد هذه النيوكليوسومات.
في الصور المجهرية الإلكترونية ، اعتمادًا على ظروف العزل ودرجة التمدد ، يمكن أن يبدو الكروماتين ليس فقط كخيط طويل مع سماكة - "خرزات" من الجسيمات النووية ، ولكن أيضًا كليف أقصر وأكثر كثافة بقطر من 30 نانومتر ، لوحظ تكوينها أثناء تفاعل هيستون H1 المرتبط بمنطقة رابط الحمض النووي وهيستون H3 ، مما يؤدي إلى التواء إضافي للحلزون المكون من ستة نيوكليوسومات في كل دور مع تكوين ملف لولبي بقطر 30 نانومتر . في هذه الحالة ، يمكن أن يتداخل بروتين هيستون مع نسخ عدد من الجينات وبالتالي تنظيم نشاطها.
نتيجة لتفاعلات الحمض النووي مع الهيستونات الموصوفة أعلاه ، يتحول جزء من الحلزون المزدوج للحمض النووي المكون من 186 زوجًا قاعديًا بمتوسط قطر 2 نانومتر وطول 57 نانومتر إلى حلزون بقطر 10 نانومتر وطوله من 5 نانومتر. مع الضغط اللاحق لهذا اللولب على ألياف قطرها 30 نانومتر ، تزداد درجة التكثيف ست مرات أخرى.
في النهاية ، ينتج عن تغليف الحمض النووي المزدوج بخمس هيستونات تكاثف للحمض النووي بمقدار 50 ضعفًا. ومع ذلك ، حتى هذه الدرجة العالية من التكثيف لا يمكن أن تفسر ما يقرب من 50000 إلى 100000 ضعف انضغاط الحمض النووي في كروموسوم الطور الطوري. لسوء الحظ ، لم تُعرف بعد تفاصيل التعبئة الإضافية للكروماتين حتى كروموسوم الطور الطوري ؛ لذلك ، يمكن فقط النظر في السمات العامة لهذه العملية.
مستويات انضغاط الحمض النووي في الكروموسومات
يتم حزم كل جزيء DNA في كروموسوم منفصل. تحتوي الخلايا البشرية ثنائية الصبغيات على 46 كروموسومًا ، والتي تقع في نواة الخلية. يبلغ الطول الإجمالي للحمض النووي لجميع كروموسومات الخلية 1.74 مترًا ، لكن قطر النواة التي تتراكم فيها الكروموسومات أصغر بملايين المرات. يتم توفير مثل هذا الحشو المضغوط للحمض النووي في الكروموسومات والكروموسومات في نواة الخلية من خلال مجموعة متنوعة من بروتينات هيستون وغير هيستون التي تتفاعل في تسلسل معين مع الحمض النووي (انظر أعلاه). يجعل انضغاط الحمض النووي في الكروموسومات من الممكن تقليل أبعاده الخطية بحوالي 10000 مرة - بشكل مشروط من 5 سم إلى 5 ميكرون. هناك عدة مستويات من الانضغاط (الشكل 10).


- الحلزون المزدوج للحمض النووي هو جزيء سالب الشحنة يبلغ قطره 2 نانومتر ويبلغ طوله عدة سنتيمترات.
- المستوى النووي- ينظر الكروماتين في المجهر الإلكتروني على أنه سلسلة من "الخرزات" - النيوكليوزومات - "على خيط". النواة هي وحدة هيكلية عالمية توجد في كل من euchromatin و heterochromatin ، في نواة الطور البيني والكروموسومات الطورية.
يتم توفير المستوى النووي للضغط بواسطة بروتينات خاصة - هيستونات. تشكل ثمانية نطاقات هيستون موجبة الشحنة نواة (نواة) الجسيم النووي الذي يلتف حوله جزيء DNA سالب الشحنة. هذا يعطي تقصيرًا بمعامل 7 ، بينما يزيد القطر من 2 إلى 11 نانومتر.
- مستوى الملف اللولبي
يتميز مستوى الملف اللولبي لتنظيم الكروموسوم بالتواء الخيوط النووية وتشكيل ألياف أكثر سمكًا بقطر 20-35 نانومتر منه - ملفات لولبية أو سوبيربد. تبلغ مساحة الملف اللولبي 11 نانومتر ، ويوجد حوالي 6-10 نيوكليوسومات في كل دورة. تعتبر تعبئة الملف اللولبي أكثر احتمالية من التعبئة الفائقة ، والتي بموجبها يكون ليف الكروماتين بقطر 20-35 نانومتر عبارة عن سلسلة من الحبيبات أو المواد الفائقة ، كل منها يتكون من ثمانية نيوكليوسومات. على مستوى الملف اللولبي ، يتم تقليل الحجم الخطي للحمض النووي بمقدار 6-10 مرات ، ويزيد القطر إلى 30 نانومتر.
- مستوى الحلقة
يتم توفير مستوى الحلقة بواسطة بروتينات مرتبطة بالحمض النووي المرتبطة بموقع غير هيستون والتي تتعرف على متواليات DNA محددة وترتبط بها ، وتشكل حلقات من حوالي 30-300 كيلو بايت. تضمن الحلقة التعبير الجيني ، أي الحلقة ليست هيكلية فحسب ، بل هي أيضًا تشكيل وظيفي. التقصير عند هذا المستوى يحدث بنسبة 20-30 مرة. يزيد القطر إلى 300 نانومتر. يمكن رؤية هياكل "فرشاة المصباح" الشبيهة بالحلقة في البويضات البرمائية في المستحضرات الخلوية. يبدو أن هذه الحلقات ملفوفة بشكل فائق وتمثل مجالات الحمض النووي ، وربما تتوافق مع وحدات نسخ الكروماتين وتكرارها. تقوم بروتينات معينة بإصلاح قواعد الحلقات ، وربما بعض مناطقها الداخلية. تسهل منظمة المجال الشبيه بالحلقة طي الكروماتين في الكروموسومات الطورية إلى هياكل حلزونية ذات أوامر أعلى.
- مستوى المجال
لم يتم دراسة مستوى مجال تنظيم الكروموسوم بشكل كافٍ. في هذا المستوى ، يُلاحظ تكوين مجالات الحلقة - هياكل الشعيرات (الألياف) بسماكة 25-30 نانومتر ، والتي تحتوي على 60٪ بروتين و 35٪ DNA و 5٪ RNA ، تكون غير مرئية عمليًا في جميع مراحل دورة الخلية مع باستثناء الانقسام الفتيلي ويتم توزيعها بشكل عشوائي إلى حد ما على نواة الخلية. يمكن رؤية هياكل "فرشاة المصباح" الشبيهة بالحلقة في البويضات البرمائية في المستحضرات الخلوية.
ترتبط مجالات الحلقة بقاعدتها بمصفوفة البروتين داخل النواة في ما يسمى بمواقع المرفقات المضمنة ، والتي يشار إليها غالبًا باسم متواليات MAR / SAR (MAR ، من المنطقة المرتبطة بالمصفوفة الإنجليزية ؛ SAR ، من مناطق مرفق السقالة الإنجليزية) - يتكسر الحمض النووي عدة مئات من أزواج القواعد الطويلة التي تتميز بمحتوى عالٍ (> 65٪) من أزواج قاعدة A / T. يبدو أن كل مجال له أصل واحد من النسخ المتماثل ويعمل كوحدة مستقلة ذات ملف فائق. يحتوي أي مجال حلقة على العديد من وحدات النسخ ، والتي من المحتمل أن يتم تنسيق عملها - يكون المجال بأكمله إما في حالة نشطة أو غير نشطة.
على مستوى المجال ، نتيجة للتعبئة المتسلسلة للكروماتين ، تقل الأبعاد الخطية للحمض النووي بنحو 200 مرة (700 نانومتر).
- مستوى الكروموسوم
على المستوى الكروموسومي ، يتكثف كروموسوم الطور الأول في الطور الأول مع ضغط مجالات الحلقة حول الإطار المحوري للبروتينات غير الهيستون. ويرافق هذا الالتفاف الفائق عملية فسفرة لجميع جزيئات H1 في الخلية. نتيجة لذلك ، يمكن تصوير كروموسوم الطور الطوري على أنه حلقات ملف لولبي معبأة بكثافة ملفوفة في دوامة ضيقة. يمكن أن يحتوي الكروموسوم البشري النموذجي على ما يصل إلى 2600 حلقة. يصل سمك هذا الهيكل إلى 1400 نانومتر (كروماتيدان) ، بينما يتم تقصير جزيء الحمض النووي بمقدار 104 مرات ، أي امتدت من 5 سم DNA إلى 5 ميكرومتر.
وظائف الكروموسومات
في التفاعل مع الآليات خارج الصبغيات ، توفر الكروموسومات
- تخزين المعلومات الوراثية
- باستخدام هذه المعلومات لإنشاء التنظيم الخلوي والحفاظ عليه
- تنظيم قراءة المعلومات الوراثية
- الاستنساخ الذاتي للمادة الوراثية
- نقل المادة الوراثية من الخلية الأم إلى الخلايا الوليدة.
هناك دليل على أنه عند تنشيط منطقة الكروماتين ، أي أثناء النسخ ، تتم إزالة هيستون H1 بشكل عكسي منه أولاً ، ثم ثماني بتات هيستون. يؤدي هذا إلى إلغاء تكثيف الكروماتين ، والانتقال المتتالي لليف كروماتين يبلغ طوله 30 نانومتر إلى خيوط بطول 10 نانومتر ، ثم تتكشف في مناطق الحمض النووي الحرة ، أي فقدان التركيب النووي.
نعلم جميعًا أن ظهور الإنسان وبعض العادات وحتى الأمراض موروثة. كل هذه المعلومات عن كائن حي مشفرة في الجينات. إذن كيف تبدو هذه الجينات سيئة السمعة ، وكيف تعمل ، وأين توجد؟
لذلك ، فإن الناقل لجميع الجينات لأي شخص أو حيوان هو الحمض النووي. تم اكتشاف هذا المركب في عام 1869 من قبل يوهان فريدريش ميشر.كيميائيا ، الحمض النووي هو حمض ديوكسي ريبونوكلييك. ماذا يعني هذا؟ كيف يحمل هذا الحمض الشفرة الجينية لجميع أشكال الحياة على كوكبنا؟
لنبدأ بالنظر إلى مكان وجود الحمض النووي. هناك العديد من العضيات في الخلية البشرية تؤدي وظائف مختلفة. يقع الحمض النووي في النواة. النواة عبارة عن عضية صغيرة محاطة بغشاء خاص يخزن جميع المواد الوراثية - DNA.
ما هي بنية جزيء الحمض النووي؟
أولاً ، دعونا نلقي نظرة على ماهية الحمض النووي. الحمض النووي جزيء طويل جدًا يتكون من عناصر هيكلية - نيوكليوتيدات. هناك 4 أنواع من النيوكليوتيدات - الأدينين (A) ، الثايمين (T) ، الجوانين (G) والسيتوزين (C). تبدو سلسلة النيوكليوتيدات بشكل تخطيطي كما يلي: GGAATTSTAAG .... هذا التسلسل من النيوكليوتيدات هو سلسلة DNA.تم فك شفرة بنية الحمض النووي لأول مرة في عام 1953 بواسطة جيمس واتسون وفرانسيس كريك.
في جزيء DNA واحد ، توجد سلسلتان من النيوكليوتيدات ملتوية حلزونيًا حول بعضها البعض. كيف تلتصق سلاسل النيوكليوتيدات هذه ببعضها البعض وتتحول إلى دوامة؟ هذه الظاهرة ترجع إلى خاصية التكامل. يعني التكامل أن بعض النيوكليوتيدات (التكميلية) فقط يمكن أن تكون معاكسة لبعضها البعض في سلسلتين. لذلك ، فإن الأدينين المعاكس هو دائمًا الثايمين ، والجوانين المقابل هو دائمًا السيتوزين فقط. وهكذا ، فإن الجوانين مكمل مع السيتوزين والأدينين مع الثايمين ، وتسمى هذه الأزواج من النيوكليوتيدات المتقابلة في سلاسل مختلفة أيضًا تكميلية.
 يمكن تمثيلها بشكل تخطيطي على النحو التالي:
يمكن تمثيلها بشكل تخطيطي على النحو التالي:
جي - سي
تي - أ
تي - أ
ج - ز
تشكل هذه الأزواج التكميلية A - T و G - C رابطة كيميائيةبين النيوكليوتيدات في الزوج ، والرابطة بين G و C أقوى من الرابطة بين A و T. وتتكون الرابطة بشكل صارم بين القواعد التكميلية ، أي أن تكوين رابطة بين G و A غير التكميليين أمر مستحيل.
"تغليف" الحمض النووي ، كيف يمكن لشريط الحمض النووي أن يصبح كروموسومًا؟
لماذا تلتف هذه السلاسل النوكليوتيدية للحمض النووي حول بعضها البعض؟ لماذا هذا مطلوب؟ الحقيقة هي أن عدد النيوكليوتيدات ضخم وتحتاج إلى مساحة كبيرة لاستيعاب هذه السلاسل الطويلة. لهذا السبب ، هناك التواء حلزوني من شريطين من الحمض النووي حول الآخر. هذه الظاهرةيسمى تصاعد. نتيجة التصعيد ، يتم تقصير سلاسل الحمض النووي بمقدار 5-6 مرات.يستخدم الجسم بعض جزيئات الحمض النووي بنشاط ، بينما نادرًا ما يستخدم البعض الآخر. تخضع جزيئات الحمض النووي التي نادرًا ما تستخدم ، بالإضافة إلى التحويل إلى حلزوني ، لـ "تغليف" أكثر إحكاما. تسمى هذه الحزمة المدمجة Supercoiling وتقصير حبلا DNA بمقدار 25-30 مرة!
كيف يتم تغليف حلزون الحمض النووي؟
 بالنسبة للفائق ، يتم استخدام بروتينات هيستون ، والتي لها مظهر وهيكل قضيب أو بكرة خيط. يتم لف خيوط لولبية من الحمض النووي على هذه "الملفات" - بروتينات هيستون. بهذه الطريقة ، يصبح الخيوط الطويلة معبأة بشكل مضغوط جدًا ولا تشغل مساحة صغيرة جدًا.
بالنسبة للفائق ، يتم استخدام بروتينات هيستون ، والتي لها مظهر وهيكل قضيب أو بكرة خيط. يتم لف خيوط لولبية من الحمض النووي على هذه "الملفات" - بروتينات هيستون. بهذه الطريقة ، يصبح الخيوط الطويلة معبأة بشكل مضغوط جدًا ولا تشغل مساحة صغيرة جدًا. إذا كان من الضروري استخدام جزيء DNA واحد أو آخر ، فإن عملية "فك اللف" تحدث ، أي أن خيط DNA "يتم فكه" من "الملف" - بروتين هيستون (إذا كان قد جُرح عليه) ويتلاشى من الحلزون إلى سلسلتين متوازيتين. وعندما يكون جزيء الحمض النووي في مثل هذه الحالة غير الملتوية ، فيمكن قراءة المعلومات الجينية الضرورية منه. علاوة على ذلك ، فإن قراءة المعلومات الجينية تحدث فقط من خيوط الحمض النووي غير الملتوية!
تسمى مجموعة من الكروموسومات فائقة الالتفاف الهيتروكروماتين، والكروموسومات المتاحة لقراءة المعلومات - كروماتين حقيقي.
ما هي الجينات وما علاقتها بالحمض النووي؟
الآن دعونا نلقي نظرة على ماهية الجينات. من المعروف أن هناك جينات تحدد فصيلة الدم ولون العينين والشعر والجلد والعديد من الخصائص الأخرى لجسمنا. الجين هو قسم محدد بدقة من الحمض النووي ، يتكون من عدد معين من النيوكليوتيدات مرتبة في تركيبة محددة بدقة. الموقع في قسم محدد بدقة من الحمض النووي يعني أن جينًا معينًا له مكانه ، ومن المستحيل تغيير هذا المكان. من المناسب إجراء مثل هذه المقارنة: يعيش الشخص في شارع معين ، في منزل وشقة معينة ، ولا يمكن لأي شخص الانتقال بشكل تعسفي إلى منزل أو شقة أخرى أو إلى شارع آخر. يعني عدد معين من النيوكليوتيدات في الجين أن كل جين لديه عدد محدد من النيوكليوتيدات ولا يمكن أن يصبح أكثر أو أقل. على سبيل المثال ، يبلغ طول الجين المشفر لإنتاج الأنسولين 60 زوجًا أساسيًا ؛ الجين الذي يشفر إنتاج هرمون الأوكسيتوسين هو 370 نقطة أساس. يعتبر تسلسل النوكليوتيدات الصارم فريدًا لكل جين ومُحدد بدقة. على سبيل المثال ، فإن تسلسل AATTAATA هو جزء من جين يرمز لإنتاج الأنسولين. من أجل الحصول على الأنسولين ، يتم استخدام مثل هذا التسلسل ؛ للحصول على الأدرينالين ، على سبيل المثال ، يتم استخدام مزيج مختلف من النيوكليوتيدات. من المهم أن نفهم أن مجموعة معينة فقط من النيوكليوتيدات تشفر "منتجًا" معينًا (الأدرينالين ، الأنسولين ، إلخ). هذا المزيج الفريد لعدد معين من النيوكليوتيدات ، يقف في "مكانه" - هذا هو الجين.
بالإضافة إلى الجينات ، توجد ما يسمى بـ "التسلسلات غير المشفرة" في سلسلة DNA. تنظم تسلسلات النوكليوتيدات غير المشفرة عمل الجينات ، وتساعد على تصاعد الكروموسوم ، وتضع علامة على نقطتي البداية والنهاية للجين. ومع ذلك ، حتى الآن ، لا يزال دور معظم التسلسلات غير المشفرة غير واضح.
ما هو الكروموسوم؟ الكروموسومات الجنسية
يُطلق على مجموع جينات الفرد اسم الجينوم. بطبيعة الحال ، لا يمكن تجميع الجينوم بأكمله في DNA واحد. ينقسم الجينوم إلى 46 زوجًا من جزيئات الحمض النووي. زوج واحد من جزيئات الحمض النووي يسمى كروموسوم. إذن فهذه الكروموسومات بالتحديد هي التي يمتلك الإنسان فيها 46 قطعة. يحمل كل كروموسوم مجموعة محددة من الجينات ، على سبيل المثال ، يحتوي الكروموسوم الثامن عشر على جينات ترميز لون العين ، إلخ. تختلف الكروموسومات عن بعضها البعض في الطول والشكل. الأشكال الأكثر شيوعًا هي في شكل X أو Y ، ولكن هناك أيضًا أشكال أخرى. يمتلك الشخص كروموسومين من نفس الشكل ، ويطلق عليهما أزواج (أزواج). فيما يتعلق بهذه الاختلافات ، فإن جميع الكروموسومات المزدوجة مرقمة - هناك 23 زوجًا. هذا يعني أن هناك زوجًا من الكروموسومات رقم 1 ، وزوج رقم 2 ، ورقم 3 ، وهكذا. يقع كل جين مسؤول عن سمة معينة على نفس الكروموسوم. في الكتيبات الحديثة للمتخصصين ، يمكن الإشارة إلى توطين الجين ، على سبيل المثال ، على النحو التالي: كروموسوم 22 ، ذراع طويل.ما هي الاختلافات بين الكروموسومات؟
 كيف تختلف الكروموسومات عن بعضها البعض؟ ماذا يعني مصطلح الذراع الطويلة؟ لنأخذ الكروموسومات على شكل X. يمكن أن يحدث تقاطع خيوط الحمض النووي بشكل صارم في المنتصف (X) ، أو يمكن أن يحدث بشكل غير مركزي. عندما لا يحدث مثل هذا التقاطع بين خيوط الحمض النووي مركزيًا ، فبالنسبة إلى نقطة التقاطع ، تكون بعض الأطراف أطول ، والبعض الآخر ، على التوالي ، أقصر. تسمى هذه النهايات الطويلة عادةً الذراع الطويلة للكروموسوم والنهايات القصيرة ، على التوالي ، الذراع القصيرة. تشغل الكروموسومات على شكل Y في الغالب أذرع طويلة ، بينما تكون الكروموسومات القصيرة صغيرة جدًا (لم يتم الإشارة إليها حتى في الصورة التخطيطية).
كيف تختلف الكروموسومات عن بعضها البعض؟ ماذا يعني مصطلح الذراع الطويلة؟ لنأخذ الكروموسومات على شكل X. يمكن أن يحدث تقاطع خيوط الحمض النووي بشكل صارم في المنتصف (X) ، أو يمكن أن يحدث بشكل غير مركزي. عندما لا يحدث مثل هذا التقاطع بين خيوط الحمض النووي مركزيًا ، فبالنسبة إلى نقطة التقاطع ، تكون بعض الأطراف أطول ، والبعض الآخر ، على التوالي ، أقصر. تسمى هذه النهايات الطويلة عادةً الذراع الطويلة للكروموسوم والنهايات القصيرة ، على التوالي ، الذراع القصيرة. تشغل الكروموسومات على شكل Y في الغالب أذرع طويلة ، بينما تكون الكروموسومات القصيرة صغيرة جدًا (لم يتم الإشارة إليها حتى في الصورة التخطيطية). يتقلب حجم الكروموسومات: أكبرها كروموسومات الأزواج رقم 1 ورقم 3 ، أصغر كروموسومات الأزواج رقم 17 ، رقم 19.
بالإضافة إلى الأشكال والأحجام ، تختلف الكروموسومات في وظائفها. من 23 زوجًا ، 22 زوجًا جسديًا وزوجًا واحدًا جنسيًا. ماذا يعني ذلك؟ تحدد الكروموسومات الجسدية جميع العلامات الخارجية للفرد ، وخصائصه الاستجابات السلوكية، النمط النفسي الوراثي ، أي جميع سمات وخصائص كل فرد. يحدد زوج من الكروموسومات الجنسية جنس الشخص: ذكر أو أنثى. هناك نوعان من الكروموسومات الجنسية البشرية - X (X) و Y (Y). إذا تم دمجهما كـ XX (x - x) - فهذه امرأة ، وإذا كان XY (x - y) - لدينا رجل أمامنا.
الأمراض الوراثية وتلف الكروموسومات
ومع ذلك ، هناك "انهيار" في الجينوم ، ثم يتم الكشف عن الأمراض الوراثية لدى البشر. على سبيل المثال ، عندما يكون هناك ثلاثة كروموسومات في 21 زوجًا من الكروموسومات بدلاً من اثنين ، يولد الشخص بمتلازمة داون.هناك العديد من "الأعطال" الصغيرة للمادة الجينية التي لا تؤدي إلى ظهور المرض ، بل على العكس من ذلك ، تعطي خصائص جيدة. تسمى كل "انهيار" المادة الجينية الطفرات. تعتبر الطفرات التي تؤدي إلى المرض أو تدهور خصائص الكائن الحي سلبية ، وتعتبر الطفرات التي تؤدي إلى تكوين خصائص مفيدة جديدة إيجابية.
ومع ذلك ، فيما يتعلق بمعظم الأمراض التي يعاني منها الناس اليوم ، فإن المرض ليس مرضًا موروثًا ، ولكنه مجرد استعداد. على سبيل المثال ، عند والد الطفل ، يمتص السكر ببطء. هذا لا يعني أن الطفل سيولد مصابًا بمرض السكري ، لكن الطفل سيكون لديه استعداد. هذا يعني أنه إذا أساء الطفل تناول الحلويات ومنتجات الدقيق ، فإنه سيصاب بمرض السكري.
اليوم ، ما يسمى ب مسندالدواء. كجزء من هذه الممارسة الطبية ، يتم تحديد الميول لدى الشخص (بناءً على تحديد الجينات المقابلة) ، ثم يتم تقديم التوصيات له - ما هو النظام الغذائي الذي يجب اتباعه ، وكيفية تبديل العمل وأنظمة الراحة بشكل صحيح حتى لا يحصل على مريض.
كيف تقرأ المعلومات المشفرة في الحمض النووي؟
لكن كيف يمكنك قراءة المعلومات الواردة في الحمض النووي؟ كيف يستخدمه جسدها؟ الحمض النووي نفسه هو نوع من المصفوفة ، لكنه ليس بسيطًا ، ولكنه مشفر. لقراءة المعلومات من مصفوفة DNA ، يتم نقلها أولاً إلى ناقل خاص - RNA. الحمض النووي الريبي هو حمض نووي كيميائيًا. وهو يختلف عن الحمض النووي في أنه يمكن أن يمر عبر الغشاء النووي إلى الخلية ، بينما يفتقر الحمض النووي لهذه القدرة (لا يمكن العثور عليه إلا في النواة). يتم استخدام المعلومات المشفرة في الخلية نفسها. لذا ، فإن الحمض النووي الريبي هو ناقل للمعلومات المشفرة من النواة إلى الخلية.كيف يحدث تخليق الحمض النووي الريبي ، كيف يتم تصنيع البروتين بمساعدة الحمض النووي الريبي؟
 خيوط الحمض النووي التي يجب "قراءة" المعلومات منها غير ملتوية ، يقترب منها إنزيم خاص ، "الباني" ، ويصنع سلسلة RNA تكميلية بالتوازي مع خيط DNA. يتكون جزيء الحمض النووي الريبي أيضًا من 4 أنواع من النيوكليوتيدات - الأدينين (A) واليوراسيل (U) والجوانين (G) والسيتوزين (C). في هذه الحالة ، تكون الأزواج التالية مكملة: الأدينين - اليوراسيل ، الجوانين - السيتوزين. كما ترون ، على عكس الحمض النووي ، يستخدم الحمض النووي الريبي اليوراسيل بدلاً من الثايمين. أي أن إنزيم "البناء" يعمل على النحو التالي: إذا رأى A في خيط DNA ، فإنه يربط Y بحبل RNA ، إذا كان G ، فإنه يعلق C ، إلخ. وهكذا ، يتم تكوين قالب من كل جين نشط أثناء النسخ - نسخة من الحمض النووي الريبي يمكن أن تمر عبر الغشاء النووي.
خيوط الحمض النووي التي يجب "قراءة" المعلومات منها غير ملتوية ، يقترب منها إنزيم خاص ، "الباني" ، ويصنع سلسلة RNA تكميلية بالتوازي مع خيط DNA. يتكون جزيء الحمض النووي الريبي أيضًا من 4 أنواع من النيوكليوتيدات - الأدينين (A) واليوراسيل (U) والجوانين (G) والسيتوزين (C). في هذه الحالة ، تكون الأزواج التالية مكملة: الأدينين - اليوراسيل ، الجوانين - السيتوزين. كما ترون ، على عكس الحمض النووي ، يستخدم الحمض النووي الريبي اليوراسيل بدلاً من الثايمين. أي أن إنزيم "البناء" يعمل على النحو التالي: إذا رأى A في خيط DNA ، فإنه يربط Y بحبل RNA ، إذا كان G ، فإنه يعلق C ، إلخ. وهكذا ، يتم تكوين قالب من كل جين نشط أثناء النسخ - نسخة من الحمض النووي الريبي يمكن أن تمر عبر الغشاء النووي. كيف يتم تكوين بروتين مشفر بواسطة جين معين؟
بعد مغادرة النواة ، يدخل الحمض النووي الريبي السيتوبلازم. بالفعل في السيتوبلازم ، يمكن أن يكون الحمض النووي الريبي ، كمصفوفة ، مدمجًا في أنظمة إنزيمية خاصة (الريبوسومات) ، والتي يمكنها توليف ، مسترشدة بمعلومات الحمض النووي الريبي ، تسلسل الأحماض الأمينية المقابل للبروتين. كما تعلم ، يتكون جزيء البروتين من الأحماض الأمينية. كيف يتمكن الريبوسوم من معرفة أي حمض أميني يرتبط بسلسلة البروتين النامية؟ يتم ذلك على أساس رمز ثلاثي. الكود الثلاثي يعني أن تسلسل ثلاثة نيوكليوتيدات من سلسلة RNA ( ثلاثة توائمعلى سبيل المثال ، GGU) رمز حمض أميني واحد (في هذه الحالة ، الجلايسين). يتم ترميز كل حمض أميني بواسطة ثلاثة توائم محددة. وهكذا ، فإن الريبوسوم "يقرأ" الثلاثي ، ويحدد الحمض الأميني الذي يجب إضافته بعد ذلك عند قراءة المعلومات في الحمض النووي الريبي. عندما تتشكل سلسلة من الأحماض الأمينية ، فإنها تأخذ شكلاً مكانيًا معينًا وتصبح بروتينًا قادرًا على القيام بالوظائف الأنزيمية والبنائية والهرمونية والوظائف الأخرى المخصصة لها.البروتين لأي كائن حي هو منتج جيني. البروتينات هي التي تحدد جميع الخصائص والصفات والمظاهر الخارجية للجينات.
على اليمين يوجد أكبر حلزون للحمض النووي البشري تم بناؤه من أشخاص على الشاطئ في فارنا (بلغاريا) ، والذي تم تضمينه في كتاب غينيس للأرقام القياسية في 23 أبريل 2016
حمض النووي الريبي منقوص الأكسجين. معلومات عامة
الحمض النووي (الحمض النووي الريبي منقوص الأكسجين) هو نوع من مخطط الحياة ، وهو رمز معقد يحتوي على بيانات حول المعلومات الوراثية. هذا الجزيء المعقد قادر على تخزين ونقل المعلومات الجينية الوراثية من جيل إلى جيل. يحدد الحمض النووي خصائص أي كائن حي مثل الوراثة والتنوع. تحدد المعلومات المشفرة فيه برنامج التطوير الكامل لأي كائن حي. العوامل المضمنة وراثيًا تحدد مسبقًا مجرى حياة كل من الشخص وأي كائن حي آخر. يمكن أن يؤثر التأثير الاصطناعي أو الطبيعي للبيئة الخارجية بشكل طفيف فقط على الشدة الكلية للصفات الجينية الفردية أو يؤثر على تطوير العمليات المبرمجة.
حمض النووي الريبي منقوص الأكسجين(DNA) هو جزيء ضخم (أحد الجزيئات الرئيسية الثلاثة ، والاثنان الآخران هما RNA والبروتينات) ، والذي يوفر التخزين والانتقال من جيل إلى جيل وتنفيذ البرنامج الجيني لتطوير وتشغيل الكائنات الحية. يحتوي الحمض النووي على معلومات حول بنية أنواع مختلفة من الحمض النووي الريبي والبروتينات.
في الخلايا حقيقية النواة (الحيوانات والنباتات والفطريات) ، يوجد الحمض النووي في نواة الخلية كجزء من الكروموسومات ، وكذلك في بعض عضيات الخلية (الميتوكوندريا والبلاستيدات). في خلايا الكائنات بدائية النواة (البكتيريا والعتائق) ، يتم توصيل جزيء DNA دائري أو خطي ، يسمى نوكليويد ، من الداخل بغشاء الخلية. لديهم وكذلك حقيقيات النوى السفلية (على سبيل المثال ، الخميرة) أيضًا جزيئات DNA صغيرة مستقلة ، معظمها دائرية تسمى البلازميدات.
من وجهة نظر كيميائية ، الحمض النووي عبارة عن جزيء بوليمري طويل يتكون من كتل متكررة - نيوكليوتيدات. يتكون كل نوكليوتيد من قاعدة نيتروجينية وسكر (ديوكسيريبوز) ومجموعة فوسفات. تتشكل الروابط بين النيوكليوتيدات في سلسلة بواسطة deoxyribose ( مع) والفوسفات ( F) المجموعات (روابط الفوسفوديستر).

أرز. 2. يتكون نوكليرتيد من قاعدة نيتروجينية وسكر (ديوكسيريبوز) ومجموعة فوسفات
في الغالبية العظمى من الحالات (باستثناء بعض الفيروسات التي تحتوي على DNA أحادي الجديلة) ، يتكون جزيء الحمض النووي من سلسلتين موجهتين بقواعد نيتروجينية لبعضهما البعض. هذا الجزيء مزدوج الشريطة ملتوي في حلزون.
هناك أربعة أنواع من القواعد النيتروجينية الموجودة في الحمض النووي (الأدينين ، والجوانين ، والثيمين ، والسيتوزين). ترتبط القواعد النيتروجينية لإحدى السلاسل بالقواعد النيتروجينية للسلسلة الأخرى بواسطة روابط هيدروجينية وفقًا لمبدأ التكامل: يتحد الأدينين مع الثايمين فقط ( في) ، الجوانين - فقط مع السيتوزين ( جي سي). هذه الأزواج هي التي تشكل "درجات" "السلم" الحلزوني للحمض النووي (انظر: الشكل 2 و 3 و 4).

أرز. 2. القواعد النيتروجينية
يسمح لك تسلسل النيوكليوتيدات "بتشفير" المعلومات الخاصة بها أنواع مختلفة RNA ، وأهمها المعلومات أو القالب (mRNA) ، الريبوسوم (rRNA) والنقل (tRNA). يتم تصنيع كل هذه الأنواع من الحمض النووي الريبي في قالب الحمض النووي عن طريق نسخ تسلسل الحمض النووي في تسلسل الحمض النووي الريبي المركب أثناء النسخ والمشاركة في التخليق الحيوي للبروتين (عملية الترجمة). بالإضافة إلى تسلسلات الترميز ، يحتوي الحمض النووي للخلية على متواليات تؤدي وظائف تنظيمية وهيكلية.

أرز. 3. تكرار الحمض النووي
يوفر موقع التركيبات الأساسية لمركبات الحمض النووي الكيميائية والنسب الكمية بين هذه المجموعات ترميزًا للمعلومات الوراثية.
تعليم DNA جديد (تكرار)
- عملية التكاثر: فك الحلزون المزدوج للحمض النووي - تخليق الخيوط التكميلية بواسطة بوليميراز الدنا - تكوين جزيئين DNA من واحد.
- ينفصل اللولب المزدوج عن فرعين عندما تكسر الإنزيمات الرابطة بين أزواج القواعد من المركبات الكيميائية.
- كل فرع هو عنصر DNA جديد. ترتبط الأزواج الأساسية الجديدة بنفس التسلسل كما في الفرع الأصلي.
عند الانتهاء من الازدواجية ، يتم تكوين حلزونات مستقلة ، يتم إنشاؤها من المركبات الكيميائية للحمض النووي الأصل ولها نفس الشفرة الجينية معها. بهذه الطريقة ، يمكن للحمض النووي أن يمزق المعلومات من خلية إلى أخرى.
معلومات أكثر تفصيلا:
هيكل الأحماض النووية

أرز. 4. القواعد النيتروجينية: الأدينين ، الجوانين ، السيتوزين ، الثايمين
حمض النووي الريبي منقوص الأكسجينيشير (DNA) إلى الأحماض النووية. احماض نوويةهي فئة من البوليمرات الحيوية غير المنتظمة التي تكون مونومراتها نيوكليوتيدات.
نيوكليوتيداتيتألف من قاعدة نيتروجينية، متصلة بخمسة كربون كربوهيدرات (بنتوز) - ديوكسيريبوز(في حالة DNA) أو ريبوز(في حالة الحمض النووي الريبي) ، الذي يتحد مع بقايا حمض الفوسفوريك (H 2 PO 3 -).
القواعد النيتروجينيةهناك نوعان: قواعد بيريميدين - اليوراسيل (فقط في الحمض النووي الريبي) ، وقواعد السيتوزين والثايمين ، وقواعد البيورين - الأدينين والجوانين.

أرز. الشكل 5. بنية النيوكليوتيدات (يسار) ، موقع النيوكليوتيدات في الحمض النووي (أسفل) وأنواع القواعد النيتروجينية (على اليمين): بيريميدين وبيورين

يتم ترقيم ذرات الكربون في جزيء البنتوز من 1 إلى 5. يتحد الفوسفات مع ذرات الكربون الثالثة والخامسة. هذه هي الطريقة التي ترتبط بها الأحماض النووية معًا لتشكيل سلسلة من الأحماض النووية. وبالتالي ، يمكننا عزل الأطراف 3 'و 5' من حبلا DNA:

أرز. 6. عزل طرفي 3 و 5 من خيط DNA
شريطين من شكل DNA الحلزون المزدوج. هذه السلاسل في دوامة موجهة في اتجاهين متعاكسين. في خيوط مختلفة من الحمض النووي ، ترتبط القواعد النيتروجينية ببعضها البعض عن طريق روابط هيدروجينية. يتحد الأدينين دائمًا مع الثايمين ، ويجمع السيتوزين دائمًا مع الجوانين. يدعي حكم التكامل(سم. مبدأ التكامل).
قاعدة التكامل:
| أيه تي جي سي |
على سبيل المثال ، إذا تم إعطاؤنا خيط DNA يحتوي على التسلسل
3'-ATGTCCTAGCTGCTCG - 5 '،
ثم ستكون السلسلة الثانية مكملة لها وموجهة في الاتجاه المعاكس - من الطرف 5 إلى النهاية 3:
5'- TACAGGATCGACGAGC- 3 '.

أرز. 7. اتجاه سلاسل جزيء DNA وربط القواعد النيتروجينية باستخدام روابط الهيدروجين
تكرار الحمض النووي
تكرار الحمض النوويهي عملية مضاعفة جزيء الحمض النووي عن طريق تخليق القالب. في معظم حالات تكاثر الحمض النووي الطبيعيالتمهيديلتخليق الحمض النووي مقتطف قصير (تم إنشاؤه مرة أخرى). يتم إنشاء مثل هذا التمهيدي الريبونوكليوتيد بواسطة إنزيم بريماز (إنزيم بريميز في بدائيات النوى ، بوليميريز الحمض النووي في حقيقيات النوى) ، ويتم استبداله لاحقًا ببوليميراز ديوكسي ريبونوكليوتيد ، والذي يؤدي عادةً وظائف الإصلاح (تصحيح الضرر الكيميائي والكسر في جزيء الحمض النووي).
يحدث النسخ المتماثل بطريقة شبه محافظة. هذا يعني أن اللولب المزدوج للحمض النووي ينفصل ويتم إكمال سلسلة جديدة على كل من سلاسله وفقًا لمبدأ التكامل. وهكذا يحتوي جزيء الدنا الوليدي على خيط واحد من الجزيء الأصلي وآخر مُصنَّع حديثًا. يحدث النسخ المتماثل في الاتجاه من 3 إلى 5 من الخيط الأصلي.

أرز. 8. تكرار (مضاعفة) جزيء DNA
تخليق الحمض النووي- هذه ليست عملية معقدة كما قد تبدو للوهلة الأولى. إذا فكرت في الأمر ، فأنت بحاجة أولاً إلى معرفة ما هو التوليف. إنها عملية الجمع بين شيء ما. يتم تكوين جزيء DNA الجديد على عدة مراحل:
1) يقطع DNA topoisomerase ، الموجود أمام شوكة النسخ ، الحمض النووي لتسهيل فكه وفكه.
2) إن هيليكاز DNA ، التي تتبع توبويزوميراز ، تؤثر على عملية "فك" حلزون الحمض النووي.
3) تقوم البروتينات المرتبطة بالحمض النووي بربط خيوط الدنا ، وكذلك تقوم بتثبيتها ، مما يمنعها من الالتصاق ببعضها البعض.
4) بوليميريز الحمض النووي δ(دلتا) ، بالتنسيق مع سرعة حركة شوكة النسخ المتماثل ، يؤدي التوليفقيادةالسلاسلشركة فرعية الحمض النووي في الاتجاه 5 "→ 3" على المصفوفةالأم خيوط من الحمض النووي في الاتجاه من نهايته 3 "إلى 5" (سرعة تصل إلى 100 زوج قاعدي في الثانية). هذه الأحداث على هذا الأمخيوط الحمض النووي محدودة.

أرز. 9. تمثيل تخطيطي لعملية تكرار الحمض النووي: (1) الخيط المتأخر (الخيط المتأخر) ، (2) الخيط الرئيسي (الخيط الرئيسي) ، (3) بوليميريز الحمض النووي α (Polα) ، (4) DNA ligase ، (5) RNA - أولي ، (6) بريماز ، (7) جزء أوكازاكي ، (8) بوليميريز الحمض النووي ، (بولي) ، (9) هيليكاز ، (10) بروتينات ربط الحمض النووي أحادية السلسلة ، (11) توبويزوميراز.
يتم وصف تركيب خيط DNA الابنة المتأخر أدناه (انظر أدناه). مخططشوكة النسخ المتماثل ووظيفة إنزيمات النسخ)
لمزيد من المعلومات حول تكرار الحمض النووي ، انظر
5) مباشرة بعد فك وتثبيت حبلا آخر من الجزيء الأصل ، ينضمبوليميريز DNA α(ألفا)وفي الاتجاه 5 "→ 3" يصنع التمهيدي (التمهيدي RNA) - تسلسل RNA على قالب DNA بطول يتراوح من 10 إلى 200 نيوكليوتيد. بعد ذلك الانزيمإزالتها من حبلا الحمض النووي.
بدلا من بوليميريز الحمض النوويα
تعلق على 3 "نهاية التمهيديبوليميريز الحمض النوويε
.
6)
بوليميريز الحمض النوويε
(إبسيلون) كما لو استمر في إطالة التمهيدي ، ولكن كركيزة مضمنةديوكسي ريبونوكليوتيدات(بمبلغ 150-200 نيوكليوتيدات). نتيجة لذلك ، يتكون خيط صلب من جزأين -RNA(أي التمهيدي) و الحمض النووي.
بوليميريز الحمض النووي εيعمل حتى يصادف التمهيدي السابقجزء اوكازاكي(تم تصنيعه قبل ذلك بقليل). ثم يتم إزالة هذا الإنزيم من السلسلة.
7) بوليميريز الحمض النووي β(بيتا) يقف في مكانبوليميرات الحمض النووي ε ،يتحرك في نفس الاتجاه (5 "→ 3") ويزيل الريبونوكليوتيدات التمهيدي أثناء إدخال ديوكسي ريبونوكليوتيدات في مكانها. يعمل الإنزيم حتى الإزالة الكاملة للبرايمر ، أي حتى ديوكسي ريبونوكليوتيد (تم تصنيعه أكثر من قبلبوليميريز الحمض النووي ε). الإنزيم غير قادر على ربط نتيجة عمله بالحمض النووي الموجود في المقدمة ، فيخرج من السلسلة.
نتيجة لذلك ، "يقع" جزء من DNA الابنة على مصفوفة الخيط الأم. يدعيجزء من أوكازاكي.
8) ligase DNA ligates اثنان متجاوران شظايا أوكازاكي ، بمعنى آخر. 5 "- نهاية المقطع ، توليفهابوليميريز الحمض النووي ε ،و 3 "نهاية سلسلة مدمجةبوليميريز الحمض النوويβ .
هيكل الحمض النووي الريبي
حمض النووي الريبي(RNA) هو واحد من ثلاثة جزيئات رئيسية (الاثنان الآخران هما DNA والبروتينات) الموجودة في خلايا جميع الكائنات الحية.
تمامًا مثل الحمض النووي ، يتكون الحمض النووي الريبي من سلسلة طويلة يُسمى فيها كل رابط النوكليوتيدات. يتكون كل نوكليوتيد من قاعدة نيتروجينية وسكر ريبوز ومجموعة فوسفات. ومع ذلك ، على عكس الحمض النووي ، فإن الحمض النووي الريبي عادة ما يحتوي على واحد بدلاً من خيطين. يتم تمثيل Pentose في RNA بواسطة الريبوز ، وليس deoxyribose (يحتوي الريبوز على مجموعة هيدروكسيل إضافية على ذرة الكربوهيدرات الثانية). أخيرًا ، يختلف الحمض النووي عن الحمض النووي الريبي في تكوين القواعد النيتروجينية: بدلا من الثايمين ( تي) اليوراسيل موجود في الحمض النووي الريبي ( يو) ، وهو أيضًا مكمل للأدينين.
يسمح تسلسل النيوكليوتيدات لـ RNA بتشفير المعلومات الجينية. تستخدم جميع الكائنات الخلوية الحمض النووي الريبي (مرنا) لبرمجة تخليق البروتين.
يتم تشكيل الحمض النووي الريبي الخلوي في عملية تسمى النسخ ، أي تخليق الحمض النووي الريبي (RNA) على قالب DNA ، بواسطة إنزيمات خاصة - بوليميرات الحمض النووي الريبي.
ثم تشارك Messenger RNAs (mRNAs) في عملية تسمى إذاعة، أولئك. تخليق البروتين على قالب الرنا المرسال بمشاركة الريبوسومات. تخضع الرناوات الأخرى لتعديلات كيميائية بعد النسخ ، وبعد تكوين الهياكل الثانوية والثالثية ، فإنها تؤدي وظائف تعتمد على نوع الحمض النووي الريبي.

أرز. 10. الفرق بين DNA و RNA من حيث القاعدة النيتروجينية: بدلاً من الثايمين (T) ، يحتوي RNA على uracil (U) ، وهو أيضًا مكمل للأدينين.
الكتابة
هذه هي عملية تخليق الحمض النووي الريبي على قالب الحمض النووي. يسترخى الحمض النووي في أحد المواقع. تحتوي إحدى السلاسل على معلومات يجب نسخها على جزيء الحمض النووي الريبي - وتسمى هذه السلسلة بالترميز. يُطلق على الخيط الثاني من الحمض النووي ، وهو مكمل لخيط التشفير ، خيط القالب. في عملية النسخ على سلسلة القالب في اتجاه 3'-5 '(على طول سلسلة DNA) ، يتم تصنيع سلسلة RNA مكملة لها. وبالتالي ، يتم إنشاء نسخة RNA من حبلا التشفير.
![]()
أرز. 11. تمثيل تخطيطي للنسخ
على سبيل المثال ، إذا تم إعطاؤنا تسلسل حبلا الترميز
3'-ATGTCCTAGCTGCTCG - 5 '،
ثم ، وفقًا لقاعدة التكامل ، ستحمل سلسلة المصفوفة التسلسل
5'- TACAGGATCGACGAGC- 3 '،
و RNA المركب منه هو التسلسل
إذاعة
ضع في اعتبارك الآلية تخليق البروتينعلى مصفوفة الحمض النووي الريبي ، وكذلك الكود الجيني وخصائصه. للتوضيح أيضًا ، على الرابط أدناه ، نوصي بمشاهدة مقطع فيديو قصير حول عمليات النسخ والترجمة التي تحدث في خلية حية:

أرز. 12. عملية تخليق البروتين: أكواد DNA لـ RNA ، رموز RNA للبروتين
الكود الجيني
الكود الجيني- طريقة ترميز تسلسل الأحماض الأمينية للبروتينات باستخدام سلسلة من النيوكليوتيدات. يتم ترميز كل حمض أميني بواسطة سلسلة من ثلاثة نيوكليوتيدات - كودون أو ثلاثي.
الكود الجيني شائع لمعظم الكائنات الأولية وحقيقيات النوى. يسرد الجدول جميع الكودونات الـ 64 ويسرد الأحماض الأمينية المقابلة. الترتيب الأساسي من 5 "إلى 3" نهاية mRNA.
الجدول 1. الكود الجيني القياسي
|
الأول نيي |
القاعدة الثانية |
الثالث نيي |
|||||||
|
يو |
ج |
أ |
جي |
||||||
|
يو |
يو يو يو |
(Phe / F) |
يو سي يو |
(Ser / S) |
يو ايه يو |
(صور / ص) |
يو جي يو |
(Cys / C) |
يو |
|
يو يو سي |
يو سي سي |
يو أ ج |
يو جي سي |
ج |
|||||
|
يو يو أ |
(ليو / لتر) |
ش كاليفورنيا |
يو أ أ |
وقف كودون ** |
يو جي أ |
وقف كودون ** |
أ |
||
|
يو يو جي |
يو سي جي |
يو ايه جي |
وقف كودون ** |
يو جي جي |
(Trp / W) |
جي |
|||
|
ج |
سي يو يو |
سي سي يو |
(دعم) |
سي أ يو |
(صاحب / ح) |
سي جي يو |
(أرج / ص) |
يو |
|
|
سي يو سي |
ج ج ج |
ج أ ج |
ج ج ج |
ج |
|||||
|
سي يو أ |
ج ج أ |
ج أ أ |
(Gln / Q) |
CGA |
أ |
||||
|
سي يو جي |
سي سي جي |
سي أ ز |
سي جي جي |
جي |
|||||
|
أ |
يو يو |
(إيل / أنا) |
أ سي يو |
(Thr / T) |
أ أ يو |
(أسن / ن) |
أ جي يو |
(Ser / S) |
يو |
|
أ يو سي |
أ ج ج |
أ أ ج |
أ ج ج |
ج |
|||||
|
أ يو أ |
أ ج أ |
أ أ أ |
(ليس / ك) |
أ ز أ |
أ |
||||
|
أ يو جي |
(ميت / م) |
أ سي جي |
أ أ ج |
ايه جي جي |
جي |
||||
|
جي |
جي يو يو |
(فال / V) |
جي سي يو |
(علاء) |
G A U |
(جنوب / د) |
جي جي يو |
(Gly / G) |
يو |
|
جي يو سي |
مجلس التعاون الخليجي |
ز أ ج |
جي جي سي |
ج |
|||||
|
جي يو أ |
جي سي أ |
جي أ أ |
(صمغ) |
جي جي أ |
أ |
||||
|
G U G |
جي سي جي |
جي ايه جي |
جي جي جي |
جي |
|||||
من بين الثلاثة توائم ، هناك 4 متواليات خاصة تعمل كـ "علامات ترقيم":
- * ثلاثي أغسطس، ويسمى أيضا ترميز الميثيونين ابدأ الكودون. يبدأ هذا الكودون في تخليق جزيء البروتين. وهكذا ، أثناء تخليق البروتين ، سيكون أول حمض أميني في التسلسل هو المثيونين دائمًا.
- **ثلاثة توائم UAA, UAGو UGAاتصل وقف الكودوناتولا ترميز لأي أحماض أمينية. في هذه التسلسلات ، يتوقف تخليق البروتين.
خصائص الكود الجيني
1. الثلاثية. يتم ترميز كل حمض أميني بواسطة سلسلة من ثلاثة نيوكليوتيدات - ثلاثي أو كودون.
2. الاستمرارية. لا توجد نيوكليوتيدات إضافية بين التوائم الثلاثة ، المعلومات تُقرأ باستمرار.
3. عدم التداخل. لا يمكن أن يكون نوكليوتيد واحد جزءًا من اثنين من ثلاثة توائم في نفس الوقت.
4. التفرد. يمكن أن يرمز كودون واحد لحمض أميني واحد فقط.

5. الانحطاط. يمكن ترميز أحد الأحماض الأمينية بعدة أكواد مختلفة.

6. براعة. الشفرة الجينية هي نفسها لجميع الكائنات الحية.
مثال. حصلنا على تسلسل حبلا الترميز:
3’- CCGATTGCACGTCGATCGTATA- 5’.
سلسلة المصفوفة سيكون لها التسلسل:
5’- جكتاكجتجكاجكتاجكات- 3’.
الآن نقوم "بتوليف" المعلوماتية RNA من هذه السلسلة:
3’- CCGAUGCACGUCGAUCGUAUA- 5’.
يذهب تخليق البروتين في الاتجاه 5 '→ 3' ، لذلك ، نحتاج إلى قلب التسلسل من أجل "قراءة" الشفرة الجينية:
5’- آوغكواغكوجكاكجواغكك- 3’.
الآن ابحث عن كود البدء AUG:
5’- AU أوغ كواجكوجكاكجواغكك- 3’.
قسّم التسلسل إلى ثلاثة توائم:
يبدو كالتالي: يتم نقل المعلومات من الحمض النووي إلى RNA (النسخ) ، من RNA إلى البروتين (الترجمة). يمكن أيضًا تكرار الحمض النووي عن طريق النسخ المتماثل ، كما أن عملية النسخ العكسي ممكنة أيضًا ، عندما يتم تصنيع الحمض النووي من قالب RNA ، ولكن هذه العملية تتميز بشكل أساسي بالفيروسات.

أرز. 13. العقيدة المركزية البيولوجيا الجزيئية
الجينوم: الجينات والكروموسومات
(المفاهيم العامة)
الجينوم - مجموع كل جينات الكائن الحي ؛ مجموعة الكروموسوم الكاملة.
تم اقتراح مصطلح "الجينوم" من قبل G. أشار المعنى الأصلي لهذا المصطلح إلى أن مفهوم الجينوم ، على عكس النمط الجيني ، هو خاصية وراثية للأنواع ككل ، وليس للفرد. مع تطور علم الوراثة الجزيئي ، تغير معنى هذا المصطلح. من المعروف أن الحمض النووي ، الذي يحمل المعلومات الجينية في معظم الكائنات الحية ، وبالتالي يشكل أساس الجينوم ، لا يشمل فقط الجينات بالمعنى الحديث للكلمة. معظميتم تمثيل الحمض النووي للخلايا حقيقية النواة من خلال تسلسل النيوكليوتيدات غير المشفر ("الزائد") الذي لا يحتوي على معلومات حول البروتينات والأحماض النووية. وبالتالي ، فإن الجزء الرئيسي من جينوم أي كائن حي هو الحمض النووي الكامل لمجموعته الفردية من الكروموسومات.
الجينات هي أجزاء من جزيئات الحمض النووي التي ترمز إلى عديد الببتيدات وجزيئات الحمض النووي الريبي.
على مدى القرن الماضي ، تغير فهمنا للجينات بشكل كبير. في السابق ، كان الجينوم عبارة عن منطقة من الكروموسوم تقوم بترميز أو تحديد سمة واحدة أو النمط الظاهري(مرئية) ، مثل لون العين.
في عام 1940 ، اقترح جورج بيدل وإدوارد تاثام تعريفًا جزيئيًا للجين. عالج العلماء جراثيم الفطريات نيوروسبورا كراساالأشعة السينية والعوامل الأخرى التي تسبب تغيرات في تسلسل الحمض النووي ( الطفرات) ، ووجدت سلالات متحولة من الفطريات فقدت بعض الإنزيمات المحددة ، والتي أدت في بعض الحالات إلى تعطيل المسار الأيضي بأكمله. توصل Beadle و Tatham إلى استنتاج مفاده أن الجين هو جزء من المادة الجينية التي تحدد أو ترمز لإنزيم واحد. هذه هي طريقة الفرضية "جين واحد ، إنزيم واحد". تم توسيع هذا المفهوم لاحقًا ليشمل التعريف "جين واحد - عديد ببتيد واحد"، نظرًا لأن العديد من الجينات تشفر بروتينات ليست إنزيمات ، ويمكن أن يكون عديد الببتيد وحدة فرعية لمركب بروتيني معقد.
على التين. يوضح الشكل 14 مخططًا لكيفية تحديد ثلاثة توائم من الحمض النووي متعدد الببتيد ، وهو تسلسل الأحماض الأمينية للبروتين ، بوساطة mRNA. يلعب أحد خيوط الحمض النووي دور قالب لتخليق الرنا المرسال ، وثلاثيات النيوكليوتيدات (الكودونات) التي تعد مكملة لثلاثة توائم من الحمض النووي. في بعض البكتيريا والعديد من حقيقيات النوى ، يتم مقاطعة تسلسلات الترميز بواسطة مناطق غير مشفرة (تسمى الإنترونات).
التعريف الكيميائي الحيوي الحديث للجين بشكل أكثر تحديدًا. الجينات هي جميع أقسام الحمض النووي التي تشفر التسلسل الأساسي للمنتجات النهائية ، والتي تشمل عديد الببتيدات أو الحمض النووي الريبي التي لها وظيفة هيكلية أو تحفيزية.
إلى جانب الجينات ، يحتوي الحمض النووي أيضًا على متواليات أخرى تؤدي وظيفة تنظيمية حصرية. التسلسلات التنظيميةقد يشير إلى بداية الجينات أو نهايتها ، أو يؤثر على النسخ ، أو يشير إلى موقع بدء النسخ المتماثل أو إعادة التركيب. يمكن التعبير عن بعض الجينات بطرق مختلفة ، حيث تعمل نفس القطعة من الحمض النووي كقالب لتشكيل منتجات مختلفة.
يمكننا الحساب تقريبًا الحد الأدنى لحجم الجينالترميز للبروتين الوسيط. يتم ترميز كل حمض أميني في سلسلة بولي ببتيد بواسطة سلسلة من ثلاثة نيوكليوتيدات ؛ تتوافق تسلسلات هذه الثلاثيات (الكودونات) مع سلسلة الأحماض الأمينية في عديد الببتيد المشفر بواسطة الجين المحدد. تتوافق سلسلة البولي ببتيد المكونة من 350 من مخلفات الأحماض الأمينية (سلسلة متوسطة الطول) مع تسلسل 1050 زوج قاعدي. ( بي بي). ومع ذلك ، فإن العديد من الجينات حقيقية النواة وبعض الجينات بدائية النواة تنقطع عن طريق قطع الحمض النووي التي لا تفعل ذلك ناقلات المعلوماتحول البروتين ، وبالتالي تبين أنها أطول بكثير مما تظهره عملية حسابية بسيطة.
كم عدد الجينات الموجودة على كروموسوم واحد؟
 أرز. 15. منظر للكروموسومات في الخلايا بدائية النواة (يسار) وحقيقية النواة. الهستونات هي فئة واسعة من البروتينات النووية التي تؤدي وظيفتين رئيسيتين: تشارك في تغليف خيوط الحمض النووي في النواة وفي التنظيم اللاجيني للعمليات النووية مثل النسخ والتكرار والإصلاح.
أرز. 15. منظر للكروموسومات في الخلايا بدائية النواة (يسار) وحقيقية النواة. الهستونات هي فئة واسعة من البروتينات النووية التي تؤدي وظيفتين رئيسيتين: تشارك في تغليف خيوط الحمض النووي في النواة وفي التنظيم اللاجيني للعمليات النووية مثل النسخ والتكرار والإصلاح.
الحمض النووي بدائيات النوى أكثر بساطة: لا تحتوي خلاياها على نواة ، لذلك يقع الحمض النووي مباشرة في السيتوبلازم في شكل نوكليويد.
 كما تعلم ، تحتوي الخلايا البكتيرية على كروموسوم على شكل خيط DNA ، معبأ في بنية مدمجة - نواة. كروموسوم بدائية النواة الإشريكية القولونية، الذي تم فك تشفير جينومه بالكامل ، هو جزيء دنا دائري (في الواقع ، ليس كذلك الدائرة اليمنى، بل حلقة بدون بداية أو نهاية) ، وتتألف من 4639675 برميلًا. يحتوي هذا التسلسل على ما يقرب من 4300 جين بروتين و 157 جينًا آخر لجزيئات الحمض النووي الريبي المستقرة. الخامس الجينات البشريةما يقرب من 3.1 مليار زوج أساسي يقابل ما يقرب من 29000 جين موجود في 24 كروموسومًا مختلفًا.
كما تعلم ، تحتوي الخلايا البكتيرية على كروموسوم على شكل خيط DNA ، معبأ في بنية مدمجة - نواة. كروموسوم بدائية النواة الإشريكية القولونية، الذي تم فك تشفير جينومه بالكامل ، هو جزيء دنا دائري (في الواقع ، ليس كذلك الدائرة اليمنى، بل حلقة بدون بداية أو نهاية) ، وتتألف من 4639675 برميلًا. يحتوي هذا التسلسل على ما يقرب من 4300 جين بروتين و 157 جينًا آخر لجزيئات الحمض النووي الريبي المستقرة. الخامس الجينات البشريةما يقرب من 3.1 مليار زوج أساسي يقابل ما يقرب من 29000 جين موجود في 24 كروموسومًا مختلفًا.
بدائيات النوى (البكتيريا).
 البكتيريا بكتريا قولونيةيحتوي على جزيء DNA دائري مزدوج السلسلة. يتكون من 4،639،675 برميل. ويصل طولها إلى 1.7 مم تقريبًا ، وهو ما يتجاوز طول الخلية نفسها بكتريا قولونيةحوالي 850 مرة. بالإضافة إلى الكروموسوم الدائري الكبير كجزء من النواة ، تحتوي العديد من البكتيريا على واحد أو أكثر من جزيئات DNA الدائرية الصغيرة الموجودة بحرية في العصارة الخلوية. تسمى هذه العناصر خارج الصبغية البلازميدات(الشكل 16).
البكتيريا بكتريا قولونيةيحتوي على جزيء DNA دائري مزدوج السلسلة. يتكون من 4،639،675 برميل. ويصل طولها إلى 1.7 مم تقريبًا ، وهو ما يتجاوز طول الخلية نفسها بكتريا قولونيةحوالي 850 مرة. بالإضافة إلى الكروموسوم الدائري الكبير كجزء من النواة ، تحتوي العديد من البكتيريا على واحد أو أكثر من جزيئات DNA الدائرية الصغيرة الموجودة بحرية في العصارة الخلوية. تسمى هذه العناصر خارج الصبغية البلازميدات(الشكل 16).
تتكون معظم البلازميدات فقط من بضعة آلاف من أزواج القواعد ، وبعضها يحتوي على أكثر من 10000 زوج قاعدي. تحمل المعلومات الجينية وتتضاعف لتكوين اللازميدات الوليدة ، التي تدخل الخلايا الوليدة أثناء انقسام الخلية الأم. توجد البلازميدات ليس فقط في البكتيريا ، ولكن أيضًا في الخميرة والفطريات الأخرى. في كثير من الحالات ، لا تقدم البلازميدات أي ميزة للخلايا المضيفة ووظيفتها الوحيدة هي التكاثر بشكل مستقل. ومع ذلك ، تحمل بعض البلازميدات جينات مفيدة للمضيف. على سبيل المثال ، يمكن للجينات الموجودة في البلازميدات أن تمنح مقاومة للعوامل المضادة للبكتيريا في الخلايا البكتيرية. تمنح البلازميدات التي تحمل جين بيتا لاكتاماز مقاومة للمضادات الحيوية بيتا لاكتام مثل البنسلين والأموكسيسيلين. يمكن أن تنتقل البلازميدات من الخلايا المقاومة للمضادات الحيوية إلى خلايا أخرى من نفس النوع أو أنواع بكتيرية مختلفة ، مما يتسبب في أن تصبح هذه الخلايا مقاومة أيضًا. الاستخدام المكثف للمضادات الحيوية هو عامل انتقائي قوي يعزز انتشار البلازميدات التي تشفر مقاومة المضادات الحيوية (وكذلك الينقولات التي تشفر الجينات المماثلة) بين البكتيريا المسببة للأمراض ، ويؤدي إلى ظهور سلالات بكتيرية ذات مقاومة للعديد من المضادات الحيوية. بدأ الأطباء في فهم مخاطر استخدام المضادات الحيوية على نطاق واسع ووصفهم فقط عند الضرورة القصوى. لأسباب مماثلة ، فإن الاستخدام الواسع للمضادات الحيوية لعلاج حيوانات المزرعة محدود.
أنظر أيضا: Ravin N.V. ، شيستاكوف S.V. جينوم بدائيات النوى // مجلة فافيلوف للوراثة والتربية ، 2013. V. 17. No. 4/2. ص 972-984.
حقيقيات النواة.
الجدول 2. الحمض النووي والجينات والكروموسومات لبعض الكائنات الحية
|
الحمض النووي المشترك ، ب. |
عدد الكروموسومات * |
العدد التقريبي للجينات |
|
|
الإشريكية القولونية(بكتيريا) |
4 639 675 |
4 435 |
|
|
خميرة الخميرة(خميرة) |
12 080 000 |
16** |
5 860 |
|
أنواع معينة انيقة(دودة خيطية) |
90 269 800 |
12*** |
23 000 |
|
نبات الأرابيدوبسيس thaliana(مصنع) |
119 186 200 |
33 000 |
|
|
ذبابة الفاكهة سوداء البطن(ذبابة الفاكهة) |
120 367 260 |
20 000 |
|
|
أرز أسيوي(أرز) |
480 000 000 |
57 000 |
|
|
عضلة المصحف(الفأر) |
2 634 266 500 |
27 000 |
|
|
الانسان العاقل(بشر) |
3 070 128 600 |
29 000 |
ملحوظة.يتم تحديث المعلومات باستمرار ؛ لمزيد من المعلومات المحدثة ، راجع مواقع الويب الخاصة بالمشروع الجينومي الفردي.
* بالنسبة لجميع حقيقيات النوى ، باستثناء الخميرة ، يتم إعطاء المجموعة ثنائية الصبغيات من الكروموسومات. ثنائي الصيغة الصبغيةعدة الكروموسومات (من دبلومة اليونانية - مزدوج و eidos - عرض) - مجموعة مزدوجة من الكروموسومات(2 ن) ، كل منها له تماثل مع نفسه.
** مجموعة هابلويد. تحتوي سلالات الخميرة البرية عادةً على ثماني مجموعات (ثماني الصبغيات) أو أكثر من هذه الكروموسومات.
*** للإناث مع اثنين من الكروموسومات X. الذكور لديهم كروموسوم X ، ولكن ليس لديهم كروموسوم Y ، أي 11 كروموسوم فقط.
تحتوي خلية الخميرة ، وهي واحدة من أصغر حقيقيات النوى ، على حمض نووي 2.6 مرة أكثر من الخلية بكتريا قولونية(الجدول 2). خلايا ذبابة الفاكهة ذبابة الفاكهةوهو كائن تقليدي من الأبحاث الجينية ، ويحتوي على 35 مرة من الحمض النووي ، وتحتوي الخلايا البشرية على حوالي 700 مرة من الحمض النووي أكثر من الخلايا بكتريا قولونية.تحتوي العديد من النباتات والبرمائيات على المزيد من الحمض النووي. يتم تنظيم المادة الوراثية للخلايا حقيقية النواة في شكل كروموسومات. مجموعة ثنائية الصبغيات من الكروموسومات (2 ن) يعتمد على نوع الكائن الحي (الجدول 2).
على سبيل المثال ، يوجد في الخلية الجسدية البشرية 46 كروموسومًا ( أرز. 17). كل كروموسوم في خلية حقيقية النواة ، كما هو موضح في الشكل. 17 ، أ، يحتوي على جزيء DNA مزدوج الشريطة كبير جدًا. أربعة وعشرون كروموسومًا بشريًا (22 كروموسومًا مزدوجًا واثنان من الكروموسومات الجنسية X و Y) تختلف في الطول بأكثر من 25 مرة. يحتوي كل كروموسوم حقيقي النواة على مجموعة محددة من الجينات.

أرز. 17. الكروموسومات حقيقية النواة.أ- زوج من الكروماتيدات الشقيقة المتصلة والمكثفة من الكروموسوم البشري. في هذا الشكل ، تبقى الكروموسومات حقيقية النواة بعد التكرار وفي الطور الطوري أثناء الانقسام. ب- مجموعة كاملة من الكروموسومات من الكريات البيض لأحد مؤلفي الكتاب. تحتوي كل خلية جسدية بشرية طبيعية على 46 كروموسومًا.

يوضح حجم ووظيفة الحمض النووي كمصفوفة لتخزين ونقل المواد الوراثية وجود عناصر هيكلية خاصة في تنظيم هذا الجزيء. في الكائنات الحية الأعلى ، يتم توزيع الحمض النووي بين الكروموسومات.
تسمى مجموعة الحمض النووي (الكروموسومات) للكائن الحي الجينوم. توجد الكروموسومات في نواة الخلية وتشكل بنية تسمى الكروماتين. الكروماتين مركب من الحمض النووي والبروتينات الأساسية (الهستونات) بنسبة 1: 1. يُقاس طول الحمض النووي عادةً بعدد أزواج النيوكليوتيدات التكميلية (بي بي). على سبيل المثال ، ثالث كروموسوم بشريقرن هو جزيء DNA بحجم 160 مليون برميل. يبلغ طوله حوالي 1 مم ، لذلك ، سيكون طول الجزيء الخطي للكروموسوم البشري الثالث 5 مم ، والحمض النووي لجميع الكروموسومات 23 (~ 3 * 10 9 bp ، MR = 1.8 * 10 12) من الصبغيات الفردية الخلية - خلية البويضة أو الحيوانات المنوية - في شكل خطي تكون 1 م ، باستثناء الخلايا الجرثومية ، تحتوي جميع خلايا جسم الإنسان (يوجد حوالي 1013 منها) على مجموعة مزدوجة من الكروموسومات. أثناء انقسام الخلية ، تتكاثر جزيئات الحمض النووي الـ 46 جميعًا وتعيد تنظيمها في 46 كروموسومًا.
إذا قمت بتوصيل جزيئات الحمض النووي للجينوم البشري (22 كروموسوم وكروموسومات X و Y أو X و X) ببعضها البعض ، تحصل على تسلسل يبلغ طوله حوالي متر واحد. ملحوظة: في جميع الثدييات وغيرها من الكائنات الذكرية غير المتجانسة ، تمتلك الإناث صبغيين X (XX) وللذكور كروموسوم X واحد وكروموسوم Y واحد (XY).
معظم الخلايا البشرية ، لذلك يبلغ الطول الإجمالي للحمض النووي لهذه الخلايا حوالي 2 متر. يمتلك الإنسان البالغ حوالي 10 14 خلية ، وبالتالي فإن الطول الإجمالي لجميع جزيئات الحمض النووي هو 2 × 10 11 كم. للمقارنة ، يبلغ محيط الأرض 4 × 10 4 كم ، والمسافة من الأرض إلى الشمس هي 1.5 × 10 8 كم. هذا هو مدى وجود الحمض النووي المضغوط بشكل مدهش في خلايانا!
في الخلايا حقيقية النواة ، توجد عضيات أخرى تحتوي على الحمض النووي - وهي الميتوكوندريا والبلاستيدات الخضراء. تم طرح العديد من الفرضيات فيما يتعلق بأصل الحمض النووي للميتوكوندريا والبلاستيدات الخضراء. وجهة النظر المقبولة عمومًا اليوم هي أنها بدائل كروموسومات البكتيريا القديمة التي اخترقت سيتوبلازم الخلايا المضيفة وأصبحت سلائف هذه العضيات. أكواد الحمض النووي للميتوكوندريا لـ tRNA و rRNA للميتوكوندريا ، بالإضافة إلى العديد من بروتينات الميتوكوندريا. يتم ترميز أكثر من 95٪ من بروتينات الميتوكوندريا بواسطة DNA النووي.
هيكل الجينات
ضع في اعتبارك بنية الجين في بدائيات النوى وحقيقيات النوى ، وأوجه التشابه والاختلاف بينهما. على الرغم من حقيقة أن الجين هو جزء من DNA يشفر بروتينًا واحدًا فقط أو RNA ، بالإضافة إلى جزء الترميز المباشر ، فإنه يتضمن أيضًا عناصر تنظيمية وعناصر هيكلية أخرى لها بنية مختلفة في بدائيات النوى وحقيقيات النوى.
تسلسل الترميز- الوحدة الهيكلية والوظيفية الرئيسية للجين ، حيث يتم ترميز ثلاثة توائم من النيوكليوتيداتتسلسل الأحماض الأمينية. يبدأ برمز البداية وينتهي برمز التوقف.
قبل وبعد تسلسل الترميز متواليات 5 'و 3' غير مترجمة. يؤدون وظائف تنظيمية ومساعدة ، على سبيل المثال ، ضمان هبوط الريبوسوم على mRNA.
تشكل التسلسلات غير المترجمة والمشفرة وحدة النسخ - منطقة الحمض النووي المنسوخة ، أي منطقة الحمض النووي التي يتم تصنيع الرنا المرسال منها.
المنهيمنطقة غير منسوخة من الحمض النووي في نهاية الجين حيث يتوقف تخليق الحمض النووي الريبي.
في بداية الجين هو المنطقة التنظيمية، الذي يتضمن المروجينو المشغل أو العامل.
المروجين- التسلسل الذي يرتبط به البوليميراز أثناء بدء النسخ. المشغل أو العامل- هذه هي المنطقة التي يمكن أن ترتبط بها بروتينات خاصة - القامعين، والتي يمكن أن تقلل من نشاط تخليق الحمض النووي الريبي من هذا الجين - بمعنى آخر ، تقليله التعبير.
هيكل الجينات في بدائيات النوى
لا تختلف الخطة العامة لهيكل الجينات في بدائيات النوى وحقيقيات النوى - كلاهما يحتوي على منطقة تنظيمية مع مروج ومشغل ، ووحدة نسخ مع تسلسلات مشفرة وغير مترجمة ، ونهائي. ومع ذلك ، فإن تنظيم الجينات في بدائيات النوى وحقيقيات النوى مختلف.

أرز. 18. مخطط بنية الجين في بدائيات النوى (البكتيريا) -يتم تكبير الصورة
في بداية ونهاية المشغل ، هناك مناطق تنظيمية مشتركة للعديد من الجينات الهيكلية. من المنطقة المنسوخة من الأوبون ، يُقرأ جزيء mRNA واحد ، والذي يحتوي على العديد من سلاسل الترميز ، ولكل منها كودون البداية والتوقف الخاص به. من كل من هذه المجالاتيتم تصنيع بروتين واحد. في هذا الطريق، يتم تصنيع العديد من جزيئات البروتين من جزيء i-RNA واحد.
تتميز بدائيات النوى بمزيج من عدة جينات في وحدة وظيفية واحدة - أوبرون. يمكن تنظيم عمل الأوبرون بواسطة جينات أخرى ، والتي يمكن إزالتها بشكل ملحوظ من الأوبرون نفسه - المنظمين. يسمى البروتين المترجم من هذا الجين كاظمة. إنه ملزم بمشغل المشغل ، وينظم التعبير عن جميع الجينات الموجودة فيه مرة واحدة.
تتميز بدائيات النوى أيضًا بالظاهرة تصريفات النسخ والترجمة.
![]()
أرز. 19- ظاهرة اقتران النسخ والترجمة في بدائيات النوى- يتم تكبير الصورة
لا يحدث هذا الاقتران في حقيقيات النوى بسبب وجود غلاف نووي يفصل السيتوبلازم ، حيث تحدث الترجمة ، عن المادة الجينية التي يحدث النسخ عليها. في بدائيات النوى ، أثناء تخليق RNA على قالب DNA ، يمكن للريبوسوم أن يرتبط فورًا بجزيء RNA المركب. وهكذا ، تبدأ الترجمة حتى قبل اكتمال النسخ. علاوة على ذلك ، يمكن للعديد من الريبوسومات أن ترتبط في وقت واحد بجزيء واحد من الحمض النووي الريبي ، وتوليف عدة جزيئات من بروتين واحد في وقت واحد.
هيكل الجينات في حقيقيات النوى
جينات وكروموسومات حقيقيات النوى منظمة بشكل معقد للغاية.
تحتوي البكتيريا في العديد من الأنواع على كروموسوم واحد فقط ، وفي جميع الحالات تقريبًا توجد نسخة واحدة من كل جين على كل كروموسوم. فقط عدد قليل من الجينات ، مثل جينات الرنا الريباسي ، موجودة في نسخ متعددة. تشكل الجينات والتسلسلات التنظيمية تقريبًا جينوم بدائيات النوى بأكمله. علاوة على ذلك ، يتوافق كل جين تقريبًا بشكل صارم مع تسلسل الحمض الأميني (أو تسلسل الحمض النووي الريبي) الذي يشفره (الشكل 14).
التنظيم الهيكلي والوظيفي للجينات حقيقية النواة أكثر تعقيدًا. جلبت دراسة الكروموسومات حقيقية النواة ، وبعد ذلك التسلسل الكامل للجينوم حقيقيات النوى ، العديد من المفاجآت. تتمتع العديد من الجينات حقيقية النواة ، إن لم يكن معظمها ، بميزة مثيرة للاهتمام: تحتوي تسلسلات النيوكليوتيدات الخاصة بها على واحد أو أكثر من مناطق الحمض النووي التي لا تكوّد تسلسل الأحماض الأمينية لمنتج متعدد الببتيد. تؤدي هذه الإدخالات غير المترجمة إلى تعطيل المراسلات المباشرة بين تسلسل النيوكليوتيدات للجين وتسلسل الأحماض الأمينية لبولي ببتيد المشفر. تسمى هذه الأجزاء غير المترجمة في الجينات الإنترونات، أو مدمج تسلسل، ومقاطع الترميز exons. في بدائيات النوى ، فقط عدد قليل من الجينات تحتوي على إنترونات.
لذلك ، في حقيقيات النوى ، لا يوجد عمليًا أي مزيج من الجينات في أوبرا ، وغالبًا ما يتم تقسيم تسلسل تشفير الجين حقيقي النواة إلى مناطق مترجمة. - اكسونات، والأقسام غير المترجمة - الإنترونات.
في معظم الحالات ، لم يتم إنشاء وظيفة introns. بشكل عام ، حوالي 1.5٪ فقط من الحمض النووي البشري "مشفر" ، أي أنه يحمل معلومات عن البروتينات أو الحمض النووي الريبي. ومع ذلك ، مع الأخذ في الاعتبار الإنترونات الكبيرة ، اتضح أن 30 ٪ من الحمض النووي البشري يتكون من جينات. نظرًا لأن الجينات تشكل نسبة صغيرة نسبيًا من الجينوم البشري ، فلا يزال هناك قدر كبير من الحمض النووي في عداد المفقودين.

أرز. 16. مخطط بنية الجين في حقيقيات النوى - يتم تكبير الصورة
من كل جين ، يتم تصنيع الجين غير الناضج ، أو ما قبل RNA ، والذي يحتوي على كل من الإنترونات والإكسونات.
بعد ذلك ، تتم عملية التضفير ، ونتيجة لذلك يتم استئصال مناطق الإنترون ، ويتم تكوين mRNA الناضج ، والذي يمكن من خلاله تصنيع البروتين.

أرز. 20. عملية الربط البديلة - يتم تكبير الصورة
يسمح مثل هذا التنظيم للجينات ، على سبيل المثال ، عندما يمكن تصنيع أشكال مختلفة من البروتين من جين واحد ، نظرًا لحقيقة أنه يمكن دمج الإكسونات في تسلسلات مختلفة أثناء التضفير.

أرز. 21. الاختلافات في بنية جينات بدائيات النوى وحقيقيات النوى - يتم تكبير الصورة
الطفرات والطفرات
طفرهيسمى التغيير المستمر في التركيب الجيني ، أي تغيير في تسلسل النوكليوتيدات.
تسمى العملية التي تؤدي إلى الطفرة الطفرات، والكائن الحي الكلالتي تحمل خلاياها نفس الطفرة متحولة.
نظرية الطفرةصاغها هيو دي فريس لأول مرة في عام 1903. تتضمن نسخته الحديثة الأحكام التالية:
1. تحدث الطفرات فجأة وفجأة.
2. تنتقل الطفرات من جيل إلى جيل.
3. يمكن أن تكون الطفرات مفيدة أو ضارة أو محايدة أو مسيطرة أو متنحية.
4. تعتمد احتمالية اكتشاف الطفرات على عدد الأفراد المدروسة.
5. يمكن أن تحدث طفرات مماثلة بشكل متكرر.
6. لا يتم توجيه الطفرات.
يمكن أن تحدث الطفرات تحت تأثير عوامل مختلفة. يميز بين الطفرات التي تسببها مطفر التأثيرات: الفيزيائية (مثل الأشعة فوق البنفسجية أو الإشعاعية) ، والمواد الكيميائية (مثل الكولشيسين أو أنواع الأكسجين التفاعلية) والبيولوجية (مثل الفيروسات). يمكن أن تحدث الطفرات أيضًا أخطاء النسخ المتماثل.
اعتمادا على شروط ظهور الطفرات تنقسم إلى من تلقاء نفسها- أي الطفرات التي نشأت في ظل الظروف العادية ، و الناجم عن- أي الطفرات التي نشأت في ظل ظروف خاصة.
يمكن أن تحدث الطفرات ليس فقط في الحمض النووي النووي ، ولكن أيضًا ، على سبيل المثال ، في الحمض النووي للميتوكوندريا أو البلاستيدات. وفقًا لذلك ، يمكننا التمييز نوويو السيتوبلازمالطفرات.
نتيجة لحدوث الطفرات ، يمكن أن تظهر ألائل جديدة في كثير من الأحيان. إذا تجاوز الأليل الطافر الأليل الطبيعي ، تسمى الطفرة مهيمن. إذا قام الأليل الطبيعي بقمع الطفرة ، يتم استدعاء الطفرة الصفة الوراثية النادرة. معظم الطفرات التي تؤدي إلى ظهور أليلات جديدة تكون متنحية.
تتميز الطفرات بالتأثير تكيفية، مما يؤدي إلى زيادة قدرة الكائن الحي على التكيف مع البيئة ، حياديالتي لا تؤثر على البقاء ضارالتي تقلل من قدرة الكائنات الحية على التكيف مع الظروف البيئية و قاتلةمما يؤدي إلى موت الكائن الحي في المراحل الأولى من تطوره.
وفقا للنتائج ، تتميز الطفرات التي تؤدي إلى فقدان وظيفة البروتين، الطفرات التي تؤدي إلى ظهور البروتين له وظيفة جديدة، وكذلك الطفرات التي تغيير جرعة الجين، وبالتالي جرعة البروتين المصنعة منه.
يمكن أن تحدث طفرة في أي خلية من خلايا الجسم. إذا حدثت طفرة في خلية جرثومية ، يتم استدعاؤها جرثومية(جرثومي أو توليدي). لا تظهر مثل هذه الطفرات في الكائن الحي الذي ظهرت فيه ، ولكنها تؤدي إلى ظهور طفرات في النسل وتكون موروثة ، لذا فهي مهمة لعلم الوراثة والتطور. إذا حدثت الطفرة في أي خلية أخرى ، يتم استدعاؤها جسدي. يمكن أن تظهر هذه الطفرة إلى حد ما في الكائن الحي الذي نشأت فيه ، على سبيل المثال ، تؤدي إلى تكوين أورام سرطانية. ومع ذلك ، فإن هذه الطفرة ليست وراثية ولا تؤثر على النسل.
يمكن أن تؤثر الطفرات على أجزاء من الجينوم بأحجام مختلفة. تخصيص وراثي, الكروموسوماتو الجينومالطفرات.
الطفرات الجينية
تسمى الطفرات التي تحدث على مقياس أصغر من جين واحد وراثي، أو منقط (منقط). تؤدي هذه الطفرات إلى تغيير واحد أو أكثر من النيوكليوتيدات في التسلسل. تشمل الطفرات الجينيةبدائل، مما يؤدي إلى استبدال أحد النوكليوتيدات بآخر ،الحذفمما يؤدي إلى فقدان أحد النيوكليوتيدات ،الإدخالات، مما يؤدي إلى إضافة نيوكليوتيد إضافي إلى التسلسل.

أرز. 23. الطفرات الجينية (النقطية)
وفقًا لآلية العمل على البروتين ، تنقسم الطفرات الجينية إلى:مرادف، والتي (نتيجة لانحلال الكود الجيني) لا تؤدي إلى تغيير في تكوين الأحماض الأمينية لمنتج البروتين ،الطفرات المغلوطة، مما يؤدي إلى استبدال أحد الأحماض الأمينية بآخر ويمكن أن يؤثر على بنية البروتين المركب ، على الرغم من عدم أهميته في كثير من الأحيان ،طفرات لا معنى لها، مما أدى إلى استبدال كود الترميز برمز التوقف ،الطفرات التي تؤدي إلى اضطراب التضفير:

أرز. 24. مخططات الطفرة
أيضًا ، وفقًا لآلية العمل على البروتين ، يتم عزل الطفرات المؤدية إلى تحول الإطار قراءة٪ sمثل عمليات الإدراج والحذف. مثل هذه الطفرات ، مثل الطفرات غير المنطقية ، على الرغم من حدوثها في نقطة واحدة في الجين ، غالبًا ما تؤثر على البنية الكاملة للبروتين ، مما قد يؤدي إلى تغيير كامل في بنيته. عندما يدور جزء من الكروموسوم 180 درجة أرز. 28. النقل
عندما يدور جزء من الكروموسوم 180 درجة أرز. 28. النقل
أرز. 29. الكروموسوم قبل وبعد الازدواجية
الطفرات الجينومية
أخيرا، الطفرات الجينيةيؤثر على الجينوم بأكمله ، أي عدد الكروموسومات يتغير. يتميز تعدد الصبغيات - زيادة في تعدد الصبغيات في الخلية ، واختلال الصيغة الصبغية ، أي تغيير في عدد الكروموسومات ، على سبيل المثال ، التثلث الصبغي (وجود متماثل إضافي في أحد الكروموسومات) و monosomy (غياب متماثل في الكروموسوم).
فيديو متعلق بالحمض النووي
استنساخ الحمض النووي ، ترميز الحمض النووي الريبي ، تخليق البروتين
(إذا لم يتم عرض الفيديو ، فهو متاح في
حسب التركيب الكيميائي للحمض النووي ( حمض النووي الريبي منقوص الأكسجين) هو البوليمر الحيوي، التي تكون مونومراتها النيوكليوتيدات. وهذا هو الحمض النووي عديد النوكليوتيد. علاوة على ذلك ، يتكون جزيء الحمض النووي عادةً من سلسلتين ملتويتين بالنسبة لبعضهما البعض على طول خط حلزوني (يُطلق عليه غالبًا "ملتوية حلزونية") ومترابطة بواسطة روابط هيدروجينية.
يمكن ثني السلاسل إلى اليسار وإلى اليمين (في أغلب الأحيان).
تحتوي بعض الفيروسات على حمض نووي واحد.
يتكون كل نوكليوتيد DNA من 1) قاعدة نيتروجينية ، 2) ديوكسيريبوز ، 3) بقايا حمض الفوسفوريك.
حلزون الحمض النووي المزدوج الأيمن
يحتوي الحمض النووي على ما يلي: الأدينين, جوانين, الثايمينو السيتوزين. الأدينين والجوانين البيورينات، والثيمين والسيتوزين - ل بيريميدين. يحتوي الحمض النووي أحيانًا على مادة اليوراسيل ، والتي عادةً ما تكون من سمات الحمض النووي الريبي ، حيث تحل محل الثايمين.
ترتبط القواعد النيتروجينية لسلسلة واحدة من جزيء الحمض النووي بالقواعد النيتروجينية لسلسلة أخرى بدقة وفقًا لمبدأ التكامل: الأدينين فقط مع الثايمين (يشكلان رابطتين هيدروجينيتين فيما بينهما) ، والجوانين فقط مع السيتوزين (ثلاث روابط).
ترتبط القاعدة النيتروجينية في النيوكليوتيدات نفسها بأول ذرة كربون في الشكل الدوري ديوكسيريبوز، وهو بنتوز (كربوهيدرات بخمس ذرات كربون). الرابطة تساهمية ، جليكوسيدية (C-N). على عكس الريبوز ، يفتقر deoxyribose إلى إحدى مجموعات الهيدروكسيل الخاصة به. تتكون حلقة الديوكسيريبوز من أربع ذرات كربون وذرة أكسجين واحدة. تقع ذرة الكربون الخامسة خارج الحلقة وهي متصلة من خلال ذرة أكسجين ببقايا حمض الفوسفوريك. أيضًا ، من خلال ذرة الأكسجين عند ذرة الكربون الثالثة ، يتم إرفاق بقايا حمض الفوسفوريك للنيوكليوتيدات المجاورة.
وهكذا ، في خيط واحد من الحمض النووي ، تكون النيوكليوتيدات المتجاورة مترابطة روابط تساهميةبين deoxyribose وحمض الفوسفوريك (phosphodiester bond). يتكون العمود الفقري من الفوسفات-ديوكسيريبوز. عموديًا عليها ، باتجاه خيط آخر من الحمض النووي ، يتم توجيه القواعد النيتروجينية ، والتي ترتبط بقواعد الخيط الثاني بواسطة روابط هيدروجينية.
إن بنية الحمض النووي هي أن العمود الفقري للسلاسل المتصلة بواسطة روابط هيدروجينية يتم توجيهها في اتجاهات مختلفة (يقولون "متعدد الاتجاهات" ، "مضاد للتوازي"). على الجانب الذي ينتهي فيه أحدهما بحمض الفوسفوريك المتصل بذرة الكربون الخامسة من الديوكسيريبوز ، ينتهي الآخر بذرة كربون ثالثة "حرة". وهذا يعني أن الهيكل العظمي لسلسلة واحدة مقلوب رأسًا على عقب ، كما كان ، بالنسبة للسلسلة الأخرى. وهكذا ، في بنية سلاسل الحمض النووي ، يتم تمييز 5 "نهايات و 3" نهايات.
عند تكرار (مضاعفة) الحمض النووي ، يستمر تركيب السلاسل الجديدة دائمًا من الطرف الخامس إلى الثالث ، حيث لا يمكن ربط النيوكليوتيدات الجديدة إلا بالطرف الثالث الحر.
في النهاية (بشكل غير مباشر عن طريق الحمض النووي الريبي) ، كل ثلاثة نيوكليوتيدات متتالية في سلسلة الحمض النووي رمز لحمض أميني واحد من البروتين.
حدث اكتشاف بنية جزيء DNA في عام 1953 بفضل عمل F. Crick و D. Watson (والذي تم تسهيله أيضًا من خلال العمل المبكر لعلماء آخرين). على الرغم من كيف مادة كيميائيةعُرف الحمض النووي منذ القرن التاسع عشر. في الأربعينيات من القرن الماضي ، أصبح من الواضح أن الحمض النووي هو الناقل للمعلومات الجينية.
يعتبر اللولب المزدوج التركيب الثانوي لجزيء الحمض النووي. في الخلايا حقيقية النواة ، توجد الغالبية العظمى من الحمض النووي في الكروموسومات ، حيث يرتبط بالبروتينات والمواد الأخرى ، ويخضع أيضًا لتعبئة أكثر كثافة.
حمض النووي الريبي منقوص الأكسجين أو الحمض النووي هو الناقل للمعلومات الجينية. يقع معظم الحمض النووي في الخلايا في النواة. إنه المكون الرئيسي للكروموسومات. في حقيقيات النوى ، يوجد الحمض النووي أيضًا في الميتوكوندريا والبلاستيدات. يتكون الحمض النووي من أحاديات النيوكليوتيدات مرتبطة تساهميًا ببعضها البعض ، مما يمثل بوليمرًا طويلًا غير متفرع. تتكون أحاديات النيوكليوتيدات التي تتكون منها الحمض النووي من ديوكسيريبوز ، وهي إحدى القواعد النيتروجينية الأربعة (الأدينين ، والجوانين ، والسيتوزين ، والثايمين) ، وبقايا حمض الفوسفوريك. عدد هذه أحاديات النوكليوتيدات كبير جدًا. على سبيل المثال ، في الخلايا بدائية النواة التي تحتوي على كروموسوم واحد ، الحمض النووي عبارة عن جزيء ضخم واحد بوزن جزيئي أكبر من 2 × 10 9.
ترتبط أحاديات النيوكليوتيدات لشريط واحد من الحمض النووي في سلسلة مع بعضها البعض بسبب التكوين روابط الفوسفوديستر التساهمية بين مجموعة deoxyribose OH من أحادي النيوكليوتيد وبقايا حمض الفوسفوريك في مجموعة أخرى. على جانب واحد من العمود الفقري المتكون من خيط واحد من الحمض النووي توجد قواعد نيتروجينية. يمكن مقارنتها بأربع خرزات مختلفة موضوعة على خيط واحد ، لأن. إنها ، كما كانت ، معلقة في سلسلة فوسفات السكر.
السؤال الذي يطرح نفسه ، كيف يمكن لهذه السلسلة الطويلة من عديد النوكليوتيد ترميز البرنامج لتطوير خلية أو حتى كائن حي بأكمله؟ يمكن الحصول على إجابة هذا السؤال من خلال فهم كيفية تكوين التركيب المكاني للحمض النووي. تم فك شفرة تركيب هذا الجزيء ووصفه بواسطة J. Watson و F. Crick في عام 1953.
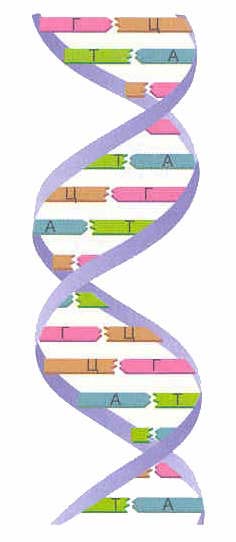
جزيئات الحمض النووي عبارة عن خيطين متوازيين ويتشكلان الحلزون الأيمن
. يبلغ عرض هذا اللولب حوالي 2 نانومتر ، لكن يمكن أن يصل طوله إلى مئات الآلاف من النانومترات. اقترح Watson and Crick نموذجًا للحمض النووي ، والذي بموجبه توجد جميع قواعد الحمض النووي داخل الحلزون ، والعمود الفقري للسكر والفوسفات في الخارج. وبالتالي ، فإن قواعد إحدى السلاسل أقرب ما يمكن إلى قواعد السلسلة الأخرى ،
لذلك تتشكل روابط هيدروجينية بينهما. إن بنية حلزون الحمض النووي هي بحيث لا يمكن فصل سلاسل عديد النوكليوتيد التي تتكون منها إلا بعد فكها.
نظرًا للقرب الأقصى بين خيطي الحمض النووي ، فإن تركيبته تحتوي على نفس الكمية من القواعد النيتروجينية من نوع واحد (الأدينين والجوانين) والقواعد النيتروجينية من نوع آخر (الثايمين والسيتوزين) ، أي أن الصيغة صالحة: أ + ج = تي + ج. هذا يرجع إلى حجم القواعد النيتروجينية ، أي طول الهياكل التي يتم تشكيلها بسبب حدوث رابطة هيدروجينية بين أزواج الأدينين - الثايمين والجوانين والسيتوزين حوالي 1.1 نانومتر. تتوافق الأبعاد الإجمالية لهذه الأزواج مع أبعاد الجزء الداخلي من حلزون الحمض النووي. لتشكيل دوامة زوجان سي تيسيكون صغيرا جدا زوجان A-G، على العكس من ذلك ، كبير جدًا. أي أن القاعدة النيتروجينية للشريط الأول من الحمض النووي تحدد القاعدة الموجودة في نفس مكان الشريط الآخر من الحمض النووي. تسمى المراسلات الصارمة للنيوكليوتيدات الموجودة في جزيء DNA في سلاسل مقترنة موازية لبعضها البعض التكامل (اختياري). الاستنساخ الدقيق أو تكرار المعلومات الجينية ممكنة على وجه التحديد بسبب هذه الميزة لجزيء الحمض النووي.
في الحمض النووي ، يتم تسجيل المعلومات البيولوجية بطريقة يمكن نسخها ونقلها إلى الخلايا السليلة. قبل انقسام الخلية تكرار (المضاعفة الذاتية ) الحمض النووي. نظرًا لأن كل سلسلة تحتوي على تسلسل نيوكليوتيد مكمل لتسلسل السلسلة الشريكة ، فإنها تحمل في الواقع نفس المعلومات الجينية. إذا قمت بفصل الخيوط واستخدمت كل منها كقالب (مصفوفة) لبناء خصلة ثانية ، فستحصل على خيطين جديدين متطابقين من الحمض النووي. هذه هي الطريقة التي يتم بها تكرار الحمض النووي في الخلية.


