DNA কি দিয়ে তৈরি। নিউক্লিওটাইড - এটা কি? ডিএনএ চেইনে নিউক্লিওটাইডের গঠন, গঠন, সংখ্যা এবং ক্রম ডিএনএ ডাবল হেলিক্স আয়ন দ্বারা স্থিতিশীল হয়
নিউক্লিক অ্যাসিড হল ম্যাক্রোমোলিকুলার পদার্থ যা মনোনিউক্লিওটাইড সমন্বিত, যা 3"5" - ফসফোডিস্টার বন্ড ব্যবহার করে একটি পলিমার চেইনে একে অপরের সাথে সংযুক্ত থাকে এবং একটি নির্দিষ্ট উপায়ে কোষে প্যাক করা হয়।
নিউক্লিক অ্যাসিড দুটি জাতের বায়োপলিমার: রাইবোনিউক্লিক অ্যাসিড (আরএনএ) এবং ডিঅক্সিরাইবোনিউক্লিক অ্যাসিড (ডিএনএ)। প্রতিটি বায়োপলিমারে নিউক্লিওটাইড থাকে যা কার্বোহাইড্রেটের অবশিষ্টাংশ (রাইবোজ, ডিঅক্সিরাইবোজ) এবং নাইট্রোজেনাস বেসগুলির মধ্যে একটি (ইউরাসিল, থাইমিন) এর মধ্যে পার্থক্য করে। তদনুসারে, নিউক্লিক অ্যাসিড তাদের নাম পেয়েছে।
ডিঅক্সিরাইবোনিউক্লিক অ্যাসিডের গঠন
নিউক্লিক অ্যাসিডের প্রাথমিক, মাধ্যমিক এবং তৃতীয় কাঠামো রয়েছে।
DNA এর প্রাথমিক গঠন
ডিএনএর প্রাথমিক গঠন হল একটি রৈখিক পলিনিউক্লিওটাইড চেইন যেখানে মনোনিউক্লিওটাইডগুলি 3", 5" ফসফোডিস্টার বন্ড দ্বারা সংযুক্ত থাকে। একটি কোষে একটি নিউক্লিক অ্যাসিড চেইন একত্রিত করার জন্য প্রাথমিক উপাদান হল নিউক্লিওসাইড 5'-ট্রাইফসফেট, যা ফসফরিক অ্যাসিডের β এবং γ অবশিষ্টাংশগুলি অপসারণের ফলে, অন্য নিউক্লিওসাইডের 3'-কার্বন পরমাণু সংযুক্ত করতে সক্ষম হয়। . এইভাবে, একটি ডিঅক্সিরাইবোজের 3" কার্বন পরমাণু একটি ফসফরিক অ্যাসিডের অবশিষ্টাংশের মাধ্যমে অন্য ডিঅক্সিরাইবোজের 5" কার্বন পরমাণুর সাথে সমন্বিতভাবে আবদ্ধ হয় এবং নিউক্লিক অ্যাসিডের একটি রৈখিক পলিনিউক্লিওটাইড চেইন তৈরি করে। তাই নাম: 3", 5"-ফসফোডিস্টার বন্ড। নাইট্রোজেনাস ঘাঁটিগুলি একটি শৃঙ্খলের নিউক্লিওটাইডের সংযোগে অংশ নেয় না (চিত্র 1।)।
একটি নিউক্লিওটাইডের ফসফরিক অ্যাসিড অণু এবং অন্যটির কার্বোহাইড্রেটের মধ্যে এই ধরনের সংযোগ, পলিনিউক্লিওটাইড অণুর একটি পেন্টোজ-ফসফেট ব্যাকবোন গঠনের দিকে পরিচালিত করে, যার উপর নাইট্রোজেনাস ঘাঁটিগুলি একের পর এক পাশ থেকে যুক্ত হয়। নিউক্লিক অ্যাসিড অণুর শৃঙ্খলে তাদের ক্রমটি বিভিন্ন জীবের কোষের জন্য কঠোরভাবে নির্দিষ্ট, যেমন একটি নির্দিষ্ট চরিত্র আছে (চারগাফের নিয়ম)।
একটি রৈখিক ডিএনএ চেইন, যার দৈর্ঘ্য চেইনের অন্তর্ভুক্ত নিউক্লিওটাইডের সংখ্যার উপর নির্ভর করে, এর দুটি প্রান্ত রয়েছে: একটিকে 3 "এন্ড বলা হয় এবং একটি মুক্ত হাইড্রক্সিল থাকে এবং অন্যটি, 5" প্রান্তে একটি ফসফরিক অ্যাসিড থাকে অবশিষ্টাংশ সার্কিটটি পোলার এবং 5"->3" এবং 3"->5" হতে পারে। একটি ব্যতিক্রম বৃত্তাকার ডিএনএ।
ডিএনএর জেনেটিক "টেক্সট" কোডন "শব্দ" দিয়ে গঠিত - নিউক্লিওটাইডের ট্রিপলেট যাকে কোডন বলা হয়। সব ধরনের আরএনএর প্রাথমিক গঠন সম্পর্কে তথ্য সম্বলিত ডিএনএ সেগমেন্টকে স্ট্রাকচারাল জিন বলে।
পলিনিউক্লিওডিটিক ডিএনএ চেইনগুলি বিশাল আকারে পৌঁছায়, তাই সেগুলি কোষে একটি নির্দিষ্ট উপায়ে প্যাক করা হয়।
DNA এর গঠন অধ্যয়ন করে, Chargaff (1949) পৃথক ডিএনএ বেসের বিষয়বস্তু সংক্রান্ত গুরুত্বপূর্ণ নিয়মিততা প্রতিষ্ঠা করে। তারা ডিএনএর গৌণ গঠন উন্মোচন করতে সাহায্য করেছিল। এই প্যাটার্নগুলোকে Chargaff এর নিয়ম বলা হয়। Chargaff নিয়ম
এই নিয়মগুলি বলে যে ডিএনএ তৈরি করার সময়, সাধারণভাবে পিউরিন এবং পাইরিমিডিন ঘাঁটির জন্য নয়, বিশেষত থাইমিনের সাথে অ্যাডেনিন এবং গুয়ানিনের সাথে সাইটোসিনের জন্য একটি বরং কঠোর চিঠিপত্র (জোড়া) অবশ্যই পালন করা উচিত। এই নিয়মগুলির উপর ভিত্তি করে, অন্যান্য বিষয়গুলির মধ্যে, 1953 সালে ওয়াটসন এবং ক্রিক ডিএনএর গৌণ কাঠামোর একটি মডেল প্রস্তাব করেছিলেন, যাকে বলা হয় ডাবল হেলিক্স (চিত্র)। |

DNA এর সেকেন্ডারি গঠন
ডিএনএর সেকেন্ডারি স্ট্রাকচার হল একটি ডাবল হেলিক্স, যার মডেলটি ডি. ওয়াটসন এবং এফ. ক্রিক 1953 সালে প্রস্তাব করেছিলেন।
একটি ডিএনএ মডেল তৈরির পূর্বশর্ত
প্রাথমিক বিশ্লেষণের ফলস্বরূপ, ধারণা ছিল যে কোনও উত্সের ডিএনএ সমান মোলার পরিমাণে চারটি নিউক্লিওটাইড ধারণ করে। যাইহোক, 1940-এর দশকে, E. Chargaff এবং তার সহকর্মীরা, বিভিন্ন জীব থেকে বিচ্ছিন্ন ডিএনএ বিশ্লেষণের ফলস্বরূপ, স্পষ্টভাবে দেখিয়েছিলেন যে তাদের মধ্যে বিভিন্ন পরিমাণগত অনুপাতের মধ্যে নাইট্রোজেনাস ঘাঁটি রয়েছে। Chargaff খুঁজে পেয়েছেন যে, যদিও এই অনুপাতগুলি একই প্রজাতির জীবের সমস্ত কোষের ডিএনএর জন্য একই, তবে বিভিন্ন প্রজাতির ডিএনএ নির্দিষ্ট নিউক্লিওটাইডের বিষয়বস্তুতে স্পষ্টভাবে আলাদা হতে পারে। এটি পরামর্শ দিয়েছে যে নাইট্রোজেনাস ঘাঁটির অনুপাতের পার্থক্যগুলি কিছু জৈবিক কোডের সাথে সম্পর্কিত হতে পারে। যদিও বিভিন্ন ডিএনএ নমুনায় স্বতন্ত্র পিউরিন এবং পাইরিমিডিন বেসের অনুপাত অসম বলে প্রমাণিত হয়েছিল, বিশ্লেষণের ফলাফলের তুলনা করার সময়, একটি নির্দিষ্ট প্যাটার্ন প্রকাশিত হয়েছিল: সমস্ত নমুনায়, পিউরিনের মোট পরিমাণ পাইরিমিডিনের মোট পরিমাণের সমান ছিল। (A + G = T + C), অ্যাডেনিনের পরিমাণ ছিল থাইমিনের পরিমাণের সমান (A = T), এবং গুয়ানিনের পরিমাণ - সাইটোসিনের পরিমাণ (G = C)। স্তন্যপায়ী কোষ থেকে বিচ্ছিন্ন ডিএনএ সাধারণত অ্যাডেনিন এবং থাইমিনে সমৃদ্ধ এবং গুয়ানিন এবং সাইটোসিনে তুলনামূলকভাবে দরিদ্র ছিল, যখন ব্যাকটেরিয়া থেকে পাওয়া ডিএনএ গুয়ানিন এবং সাইটোসিনে সমৃদ্ধ এবং অ্যাডেনিন এবং থাইমিনে অপেক্ষাকৃত দরিদ্র ছিল। এই তথ্যগুলি বাস্তব উপাদানের একটি গুরুত্বপূর্ণ অংশ গঠন করেছিল, যার ভিত্তিতে পরবর্তীতে ওয়াটসন-ক্রিক ডিএনএ কাঠামোর মডেল তৈরি করা হয়েছিল।
ডিএনএর সম্ভাব্য কাঠামোর আরেকটি গুরুত্বপূর্ণ পরোক্ষ ইঙ্গিত ছিল প্রোটিন অণুর গঠনের ওপর এল. পলিং-এর তথ্য। পলিং দেখিয়েছেন যে একটি প্রোটিন অণুতে অ্যামিনো অ্যাসিড চেইনের বিভিন্ন স্থিতিশীল কনফিগারেশন সম্ভব। পেপটাইড চেইনের একটি সাধারণ কনফিগারেশন - α-হেলিক্স - একটি নিয়মিত হেলিকাল গঠন। এই জাতীয় কাঠামোর সাথে, চেইনের সংলগ্ন বাঁকগুলিতে অবস্থিত অ্যামিনো অ্যাসিডগুলির মধ্যে হাইড্রোজেন বন্ধন তৈরি করা সম্ভব। পলিং 1950 সালে পলিপেপটাইড চেইনের α-হেলিকাল কনফিগারেশন বর্ণনা করেছিলেন এবং পরামর্শ দিয়েছিলেন যে ডিএনএ অণুগুলিরও সম্ভবত হাইড্রোজেন বন্ড দ্বারা স্থির একটি হেলিকাল গঠন রয়েছে।
যাইহোক, ডিএনএ অণুর গঠন সম্পর্কে সবচেয়ে মূল্যবান তথ্য এক্স-রে ডিফ্র্যাকশন বিশ্লেষণের ফলাফল দ্বারা সরবরাহ করা হয়েছিল। এক্স-রে, একটি ডিএনএ স্ফটিকের মধ্য দিয়ে যাওয়া, বিচ্ছুরণের মধ্য দিয়ে যায়, অর্থাৎ, তারা নির্দিষ্ট দিকগুলিতে বিচ্যুত হয়। রশ্মির বিক্ষেপণের মাত্রা এবং প্রকৃতি অণুগুলির গঠনের উপর নির্ভর করে। এক্স-রে ডিফ্র্যাকশন প্যাটার্ন (চিত্র 3) অভিজ্ঞ চোখকে অধ্যয়নের অধীনে থাকা পদার্থের অণুগুলির গঠন সম্পর্কিত পরোক্ষ ইঙ্গিত দেয়। ডিএনএ এক্স-রে ডিফ্র্যাকশন প্যাটার্নের বিশ্লেষণের ফলে এই সিদ্ধান্তে উপনীত হয় যে নাইট্রোজেনাস বেসগুলি (একটি সমতল আকৃতির) প্লেটের স্তুপের মতো স্তুপীকৃত। এক্স-রে প্যাটার্নগুলি স্ফটিক ডিএনএ গঠনের তিনটি প্রধান সময়কাল সনাক্ত করা সম্ভব করেছে: 0.34, 2 এবং 3.4 nm।

ওয়াটসন-ক্রিক ডিএনএ মডেল
Chargaff এর বিশ্লেষণাত্মক তথ্য থেকে শুরু করে, উইলকিন্সের এক্স-রে এবং রসায়নবিদ যারা একটি অণুতে পরমাণুর মধ্যে সঠিক দূরত্ব, একটি প্রদত্ত পরমাণুর বন্ধনের মধ্যবর্তী কোণ এবং পরমাণুর আকার সম্পর্কে তথ্য প্রদান করেছিলেন, ওয়াটসন এবং ক্রিক একটি নির্দিষ্ট স্কেলে ডিএনএ অণুর পৃথক উপাদানগুলির ভৌত মডেল তৈরি করুন এবং তাদের একে অপরের সাথে এমনভাবে "সামঞ্জস্য" করুন যাতে ফলাফল সিস্টেমটি বিভিন্ন পরীক্ষামূলক ডেটার সাথে মিলে যায় [দেখা] .
এমনকি আগেও, এটি জানা ছিল যে একটি ডিএনএ শৃঙ্খলে সংলগ্ন নিউক্লিওটাইডগুলি ফসফোডিস্টার সেতু দ্বারা সংযুক্ত থাকে যা একটি নিউক্লিওটাইডের ডিঅক্সিরাইবোজের 5'-কার্বন পরমাণুকে পরবর্তী নিউক্লিওটাইডের ডিঅক্সিরাইবোজের 3'-কার্বন পরমাণুর সাথে সংযুক্ত করে। ওয়াটসন এবং ক্রিকের কোন সন্দেহ ছিল না যে 0.34 এনএম সময়কাল একটি ডিএনএ স্ট্র্যান্ডের ধারাবাহিক নিউক্লিওটাইডের মধ্যে দূরত্বের সাথে মিলে যায়। আরও, এটি অনুমান করা যেতে পারে যে 2 এনএম সময়কাল চেইনের পুরুত্বের সাথে মিলে যায়। এবং 3.4 এনএম সময়কালের সাথে বাস্তব কাঠামোটি কী তা ব্যাখ্যা করার জন্য, ওয়াটসন এবং ক্রিক, সেইসাথে আগে পলিং, ধরে নিয়েছিলেন যে চেইনটি একটি সর্পিল আকারে পেঁচানো হয়েছে (বা, আরও স্পষ্টভাবে, একটি হেলিক্স গঠন করে, যেহেতু সর্পিল এর কঠোর অর্থে শব্দটি প্রাপ্ত হয় যখন বাঁকগুলি মহাকাশে একটি নলাকার পৃষ্ঠের পরিবর্তে একটি শঙ্কুময় গঠন করে)। তারপর 3.4 nm সময়কাল এই সর্পিলটির ধারাবাহিক বাঁকগুলির মধ্যে দূরত্বের সাথে মিলিত হবে। এই জাতীয় সর্পিল খুব ঘন বা কিছুটা প্রসারিত হতে পারে, অর্থাৎ, এর বাঁকগুলি সমতল বা খাড়া হতে পারে। যেহেতু 3.4 এনএম সময়কাল পরপর নিউক্লিওটাইডের (0.34 এনএম) মধ্যে দূরত্বের ঠিক 10 গুণ, এটি স্পষ্ট যে হেলিক্সের প্রতিটি সম্পূর্ণ বাঁকটিতে 10টি নিউক্লিওটাইড রয়েছে। এই তথ্যগুলি থেকে, ওয়াটসন এবং ক্রিক 2 এনএম ব্যাস সহ একটি হেলিক্সে পেঁচানো একটি পলিনিউক্লিওটাইড চেইনের ঘনত্ব গণনা করতে সক্ষম হয়েছিল, যার মধ্যে 3.4 এনএমের সমান দূরত্ব ছিল। দেখা গেল যে এই জাতীয় স্ট্র্যান্ডের ঘনত্ব ডিএনএর প্রকৃত ঘনত্বের অর্ধেক হবে, যা আগে থেকেই জানা ছিল। আমাকে অনুমান করতে হয়েছিল যে ডিএনএ অণু দুটি চেইন নিয়ে গঠিত - এটি নিউক্লিওটাইডের একটি ডাবল হেলিক্স।
পরবর্তী কাজটি ছিল, অবশ্যই, ডাবল হেলিক্স গঠনকারী দুটি স্ট্র্যান্ডের মধ্যে স্থানিক সম্পর্ককে ব্যাখ্যা করা। তাদের ভৌত মডেলে চেইনের বিন্যাসের বেশ কয়েকটি রূপ চেষ্টা করার পর, ওয়াটসন এবং ক্রিক আবিষ্কার করেছেন যে সমস্ত উপলভ্য ডেটার জন্য সবচেয়ে উপযুক্ত হল একটি যেখানে দুটি পলিনিউক্লিওটাইড হেলিস বিপরীত দিকে যায়; এই ক্ষেত্রে, চিনি এবং ফসফেটের অবশিষ্টাংশ সমন্বিত চেইনগুলি একটি ডাবল হেলিক্সের পৃষ্ঠ তৈরি করে এবং পিউরিন এবং পাইরিমিডিনগুলি ভিতরে অবস্থিত। দুটি চেইনের অন্তর্গত একে অপরের বিপরীতে অবস্থিত বেসগুলি হাইড্রোজেন বন্ড দ্বারা জোড়ায় জোড়ায় সংযুক্ত থাকে; এই হাইড্রোজেন বন্ডগুলিই চেইনগুলিকে একসাথে ধরে রাখে, এইভাবে অণুর সামগ্রিক কনফিগারেশন ঠিক করে।
ডিএনএ ডাবল হেলিক্সকে একটি হেলিকাল দড়ির মই হিসাবে ভাবা যেতে পারে, যার দন্ডগুলি অনুভূমিক থাকে। তারপরে দুটি অনুদৈর্ঘ্য দড়ি চিনি এবং ফসফেট অবশিষ্টাংশের চেইনগুলির সাথে মিলিত হবে এবং ক্রসবারগুলি হাইড্রোজেন বন্ড দ্বারা সংযুক্ত নাইট্রোজেনাস ঘাঁটির সাথে মিলিত হবে।
সম্ভাব্য মডেলগুলির আরও অধ্যয়নের ফলে, ওয়াটসন এবং ক্রিক এই সিদ্ধান্তে উপনীত হন যে প্রতিটি "ক্রসবার"-এ একটি পিউরিন এবং একটি পাইরিমিডিন থাকা উচিত; 2 nm সময়কালে (ডাবল হেলিক্সের ব্যাসের সাথে সম্পর্কিত), দুটি পিউরিনের জন্য পর্যাপ্ত জায়গা থাকবে না এবং দুটি পাইরিমিডিন সঠিক হাইড্রোজেন বন্ধন গঠনের জন্য পর্যাপ্ত কাছাকাছি থাকতে পারে না। বিশদ মডেলের একটি গভীর গবেষণায় দেখা গেছে যে অ্যাডেনিন এবং সাইটোসিন সঠিক আকারের সংমিশ্রণ তৈরি করে, এখনও এমনভাবে সাজানো যায়নি যে তাদের মধ্যে হাইড্রোজেন বন্ধন তৈরি হয়। অনুরূপ রিপোর্ট গুয়ানাইন-থাইমিন সংমিশ্রণকে বাদ দিতে বাধ্য করেছিল, যখন অ্যাডেনিন-থাইমিন এবং গুয়ানিন-সাইটোসিনের সংমিশ্রণগুলি বেশ গ্রহণযোগ্য বলে মনে হয়েছিল। হাইড্রোজেন বন্ধনের প্রকৃতি এমন যে থাইমিনের সাথে অ্যাডেনিন জোড়া এবং সাইটোসিনের সাথে গুয়ানিনের জোড়া। নির্দিষ্ট বেস পেয়ারিংয়ের এই ধারণাটি "চারগাফ নিয়ম" ব্যাখ্যা করা সম্ভব করেছে, যে অনুসারে যে কোনও ডিএনএ অণুতে অ্যাডেনিনের পরিমাণ সর্বদা থাইমিনের পরিমাণের সমান এবং গুয়ানিনের পরিমাণ সর্বদা সাইটোসিনের পরিমাণের সমান। . অ্যাডেনিন এবং থাইমিনের মধ্যে দুটি হাইড্রোজেন বন্ধন এবং তিনটি গুয়ানিন এবং সাইটোসিনের মধ্যে তৈরি হয়। এক শৃঙ্খলে প্রতিটি অ্যাডেনিনের বিরুদ্ধে হাইড্রোজেন বন্ধন গঠনে এই বিশেষত্বের কারণে, থাইমিন অন্যটিতে থাকে; একইভাবে, প্রতিটি গুয়ানিনের বিপরীতে শুধুমাত্র সাইটোসিন স্থাপন করা যেতে পারে। এইভাবে, চেইনগুলি একে অপরের পরিপূরক, অর্থাৎ, একটি শৃঙ্খলে নিউক্লিওটাইডগুলির ক্রম অনন্যভাবে অন্যটিতে তাদের ক্রম নির্ধারণ করে। দুটি চেইন বিপরীত দিকে চলে এবং তাদের ফসফেট শেষ গ্রুপগুলি ডাবল হেলিক্সের বিপরীত প্রান্তে থাকে।
তাদের গবেষণার ফলস্বরূপ, 1953 সালে ওয়াটসন এবং ক্রিক ডিএনএ অণুর গঠনের জন্য একটি মডেল প্রস্তাব করেছিলেন (চিত্র 3), যা বর্তমানের সাথে প্রাসঙ্গিক। মডেল অনুসারে, একটি ডিএনএ অণু দুটি পরিপূরক পলিনিউক্লিওটাইড চেইন নিয়ে গঠিত। প্রতিটি ডিএনএ স্ট্র্যান্ড একটি পলিনিউক্লিওটাইড যা কয়েক হাজার নিউক্লিওটাইড নিয়ে গঠিত। এটিতে, প্রতিবেশী নিউক্লিওটাইডগুলি একটি শক্তিশালী সমযোজী বন্ধনের দ্বারা ফসফরিক অ্যাসিডের অবশিষ্টাংশ এবং ডিঅক্সিরিবোজের সংমিশ্রণের কারণে একটি নিয়মিত পেন্টোজ-ফসফেট ব্যাকবোন গঠন করে। একটি পলিনিউক্লিওটাইড চেইনের নাইট্রোজেনাস বেসগুলি অন্যটির নাইট্রোজেনাস বেসের বিরুদ্ধে কঠোরভাবে সংজ্ঞায়িত ক্রমে সাজানো হয়। পলিনিউক্লিওটাইড শৃঙ্খলে নাইট্রোজেনাস ঘাঁটির পরিবর্তন অনিয়মিত।
ডিএনএ শৃঙ্খলে নাইট্রোজেনাস ঘাঁটিগুলির বিন্যাস পরিপূরক (গ্রীক "পরিপূরক" থেকে - যোগ), অর্থাৎ অ্যাডেনিন (A) এর বিরুদ্ধে সর্বদা থাইমিন (টি), এবং গুয়ানিনের বিরুদ্ধে (জি) - শুধুমাত্র সাইটোসিন (সি)। এটি এই সত্য দ্বারা ব্যাখ্যা করা হয়েছে যে A এবং T, পাশাপাশি G এবং C, একে অপরের সাথে কঠোরভাবে সঙ্গতিপূর্ণ, অর্থাৎ একে অপরের পরিপূরক. এই চিঠিপত্র ঘাঁটিগুলির রাসায়নিক কাঠামো দ্বারা দেওয়া হয়, যা পিউরিন এবং পাইরিমিডিনের জোড়ায় হাইড্রোজেন বন্ধন গঠনের অনুমতি দেয়। A এবং T এর মধ্যে দুটি বন্ধন রয়েছে, G এবং C এর মধ্যে - তিনটি। এই বন্ধনগুলি মহাকাশে ডিএনএ অণুর আংশিক স্থিতিশীলতা প্রদান করে। ডাবল হেলিক্সের স্থায়িত্ব সরাসরি G≡C বন্ডের সংখ্যার সমানুপাতিক, যা A=T বন্ডের চেয়ে বেশি স্থিতিশীল।
ডিএনএ-র এক স্ট্র্যান্ডে নিউক্লিওটাইডের পরিচিত ক্রম, পরিপূরকতার নীতির দ্বারা, অন্য স্ট্র্যান্ডের নিউক্লিওটাইডগুলি স্থাপন করা সম্ভব করে।
উপরন্তু, এটা পাওয়া গেছে যে নাইট্রোজেনাস ঘাঁটি একটি সুগন্ধযুক্ত গঠন, মধ্যে জলীয় দ্রবণএকে অপরের উপরে সাজানো হয়, গঠন করা হয়, যেমন ছিল, মুদ্রার স্তুপ। এর স্ট্যাক গঠনের এই প্রক্রিয়া জৈব অণুস্ট্যাকিং বলা হয়। বিবেচিত ওয়াটসন-ক্রিক মডেলের ডিএনএ অণুর পলিনিউক্লিওটাইড চেইনগুলির একটি অনুরূপ ভৌত রাসায়নিক অবস্থা রয়েছে, তাদের নাইট্রোজেনাস ঘাঁটিগুলি মুদ্রার স্তুপের আকারে সাজানো হয়েছে, যার সমতলগুলির মধ্যে ভ্যান ডার ওয়ালস মিথস্ক্রিয়া (স্ট্যাকিং মিথস্ক্রিয়া) ঘটে।
ভ্যান ডার ওয়ালসের কারণে (উল্লম্বভাবে) পরিপূরক ঘাঁটি (অনুভূমিকভাবে) এবং পলিনিউক্লিওটাইড শৃঙ্খলে বেস প্লেনের মধ্যে স্ট্যাকিং মিথস্ক্রিয়া ডিএনএ অণুকে মহাকাশে অতিরিক্ত স্থিতিশীলতা প্রদান করে।
উভয় চেইনের সুগার-ফসফেট ব্যাকবোনগুলি বাইরের দিকে পরিণত হয় এবং ভিত্তিগুলি একে অপরের দিকে ভিতরের দিকে থাকে। ডিএনএ-তে স্ট্র্যান্ডগুলির দিক সমান্তরাল (এগুলির একটির দিক 5"->3", অন্যটির - 3"->5", অর্থাৎ একটি স্ট্র্যান্ডের 3"-প্রান্ত 5"-এন্ডের বিপরীতে অবস্থিত অন্যের।) চেইনগুলি একটি সাধারণ অক্ষের সাথে ডান হেলিক্স গঠন করে। হেলিক্সের একটি বাঁক হল 10টি নিউক্লিওটাইড, টার্নের আকার 3.4 এনএম, প্রতিটি নিউক্লিওটাইডের উচ্চতা 0.34 এনএম, হেলিক্সের ব্যাস 2.0 এনএম। একটি স্ট্র্যান্ড অন্যটির চারপাশে ঘোরানোর ফলে, ডিএনএ ডাবল হেলিক্সে একটি বড় খাঁজ (প্রায় 20 Å ব্যাস) এবং একটি ছোট খাঁজ (প্রায় 12 Å) তৈরি হয়। ওয়াটসন-ক্রিক ডাবল হেলিক্সের এই ফর্মটিকে পরে বি-ফর্ম বলা হয়। কোষে, ডিএনএ সাধারণত বি আকারে বিদ্যমান থাকে, যা সবচেয়ে স্থিতিশীল।
ডিএনএর কার্যাবলী
প্রস্তাবিত মডেলটি ডিঅক্সিরাইবোনিউক্লিক অ্যাসিডের অনেক জৈবিক বৈশিষ্ট্য ব্যাখ্যা করেছে, যার মধ্যে রয়েছে জেনেটিক তথ্যের সঞ্চয়স্থান এবং জিনের বৈচিত্র্য, যা 4টি নিউক্লিওটাইডের পরপর সংমিশ্রণ এবং একটি জেনেটিক কোডের অস্তিত্বের সত্যতা দ্বারা সরবরাহিত জিনের বৈচিত্র্য। স্ব-পুনরুৎপাদন এবং জেনেটিক তথ্য প্রেরণ করে, যা প্রতিলিপি প্রক্রিয়া দ্বারা প্রদত্ত, এবং প্রোটিন আকারে জেনেটিক তথ্যের বাস্তবায়ন, সেইসাথে এনজাইম প্রোটিনের সাহায্যে গঠিত অন্য কোনো যৌগ।
DNA এর মৌলিক কাজ।
- ডিএনএ হল জেনেটিক তথ্যের বাহক, যা জেনেটিক কোডের অস্তিত্বের সত্যতা দ্বারা নিশ্চিত করা হয়।
- কোষ এবং জীবের প্রজন্মের মধ্যে বংশবৃদ্ধি এবং জেনেটিক তথ্য প্রেরণ। এই ফাংশন প্রতিলিপি প্রক্রিয়া দ্বারা উপলব্ধ করা হয়.
- প্রোটিন আকারে জেনেটিক তথ্য বাস্তবায়ন, সেইসাথে এনজাইম প্রোটিনের সাহায্যে গঠিত অন্য কোনো যৌগ। এই ফাংশন ট্রান্সক্রিপশন এবং অনুবাদ প্রক্রিয়া দ্বারা উপলব্ধ করা হয়.

ডাবল-স্ট্র্যান্ডেড ডিএনএর সংগঠনের ফর্ম
DNA বিভিন্ন ধরনের ডাবল হেলিক্স গঠন করতে পারে (চিত্র 4)। বর্তমানে, ছয়টি ফর্ম ইতিমধ্যেই পরিচিত (এ থেকে ই এবং জেড ফর্ম)।
রোজালিন্ড ফ্র্যাঙ্কলিন দ্বারা প্রতিষ্ঠিত ডিএনএর কাঠামোগত রূপগুলি জলের সাথে নিউক্লিক অ্যাসিড অণুর সম্পৃক্ততার উপর নির্ভর করে। এক্স-রে বিবর্তন বিশ্লেষণ ব্যবহার করে ডিএনএ তন্তুগুলির গবেষণায়, এটি দেখানো হয়েছিল যে এক্স-রে বিচ্ছুরণ প্যাটার্ন আমূলভাবে নির্ভর করে কোন আপেক্ষিক আর্দ্রতার উপর, এই ফাইবারের জলের স্যাচুরেশন কত ডিগ্রিতে পরীক্ষাটি হয়। যদি ফাইবার পর্যাপ্ত পরিমাণে জলে পরিপূর্ণ হয় তবে একটি রেডিওগ্রাফ পাওয়া যায়। শুকিয়ে গেলে, একটি সম্পূর্ণ ভিন্ন এক্স-রে প্যাটার্ন দেখা যায়, যা উচ্চ-আদ্রতাযুক্ত ফাইবারের এক্স-রে প্যাটার্ন থেকে খুব আলাদা।
উচ্চ আর্দ্রতার ডিএনএর অণুকে বি-শেপ বলে. শারীরবৃত্তীয় অবস্থার অধীনে (নিম্ন লবণের ঘনত্ব, হাইড্রেশনের উচ্চ মাত্রা), ডিএনএ-এর প্রভাবশালী কাঠামোগত ধরন হল বি-ফর্ম (ডবল-স্ট্র্যান্ডেড ডিএনএর প্রধান রূপ হল ওয়াটসন-ক্রিক মডেল)। এই ধরনের একটি অণুর হেলিক্স পিচ হল 3.4 nm। "মুদ্রা" - নাইট্রোজেনাস ঘাঁটিগুলির পেঁচানো স্ট্যাকের আকারে প্রতি টার্নে 10টি পরিপূরক জোড়া রয়েছে। স্ট্যাকের দুটি বিপরীত "মুদ্রা" এর মধ্যে হাইড্রোজেন বন্ড দ্বারা স্ট্যাকগুলিকে একত্রে রাখা হয় এবং ফসফোডিস্টার ব্যাকবোনের দুটি ফিতা দিয়ে একটি ডান হাতের হেলিক্সে পেঁচানো "কুণ্ডলীকৃত" হয়। নাইট্রোজেনাস ঘাঁটিগুলির সমতলগুলি হেলিক্সের অক্ষের সাথে লম্ব। প্রতিবেশী পরিপূরক জোড়া একে অপরের সাপেক্ষে 36° দ্বারা ঘোরানো হয়। হেলিক্সের ব্যাস 20Å, পিউরিন নিউক্লিওটাইড 12Å এবং পাইরিমিডিন নিউক্লিওটাইড 8Å দখল করে।
নিম্ন আর্দ্রতার ডিএনএ অণুকে এ-ফর্ম বলা হয়. A-ফর্মটি কম উচ্চ হাইড্রেশনের অবস্থার অধীনে এবং Na + বা K + আয়নগুলির উচ্চ পরিমাণে গঠিত হয়। এই বৃহত্তর ডান হাতের কনফর্মেশনে প্রতি টার্নে 11টি বেস জোড়া রয়েছে। নাইট্রোজেনাস ঘাঁটিগুলির সমতলগুলি হেলিক্সের অক্ষের দিকে একটি শক্তিশালী প্রবণতা রয়েছে, তারা স্বাভাবিক থেকে হেলিক্সের অক্ষে 20° দ্বারা বিচ্যুত হয়। এটি 5 Å ব্যাস সহ একটি অভ্যন্তরীণ শূন্যতার উপস্থিতি বোঝায়। সংলগ্ন নিউক্লিওটাইডগুলির মধ্যে দূরত্ব হল 0.23 nm, কুণ্ডলীর দৈর্ঘ্য 2.5 nm এবং হেলিক্সের ব্যাস হল 2.3 nm।
প্রাথমিকভাবে, DNA-এর A-ফর্ম কম গুরুত্বপূর্ণ বলে মনে করা হয়েছিল। যাইহোক, পরে দেখা গেল যে DNA-এর A-ফর্ম, সেইসাথে B-ফর্মের জৈবিক গুরুত্ব অনেক বেশি। টেমপ্লেট-সিড কমপ্লেক্সের আরএনএ-ডিএনএ হেলিক্সের এ-ফর্ম রয়েছে, সেইসাথে আরএনএ-আরএনএ হেলিক্স এবং আরএনএ হেয়ারপিন গঠন রয়েছে (রাইবোজের 2'-হাইড্রক্সিল গ্রুপ আরএনএ অণুকে বি-ফর্ম গঠন করতে দেয় না) . ডিএনএর এ-ফর্ম স্পোরগুলিতে পাওয়া যায়। এটি প্রতিষ্ঠিত হয়েছে যে ডিএনএর A-ফর্ম B-ফর্মের চেয়ে 10 গুণ বেশি UV রশ্মির প্রতিরোধী।
A-ফর্ম এবং B-ফর্ম বলা হয় ক্যানোনিকাল ফর্মডিএনএ।
ফর্ম সি-ইএছাড়াও ডান-হাতি, তাদের গঠন শুধুমাত্র বিশেষ পরীক্ষায় লক্ষ্য করা যায়, এবং দৃশ্যত, তারা ভিভোতে বিদ্যমান নেই। ডিএনএর সি-ফর্মের গঠন বি-ডিএনএর অনুরূপ। প্রতি টার্নে বেস জোড়ার সংখ্যা 9.33, এবং হেলিক্সের দৈর্ঘ্য 3.1 nm। বেস জোড়াগুলি অক্ষের লম্ব অবস্থানের সাপেক্ষে 8 ডিগ্রি কোণে ঝুঁকে থাকে। খাঁজগুলো B-DNA এর খাঁজের কাছাকাছি। এই ক্ষেত্রে, প্রধান খাঁজটি কিছুটা ছোট, এবং ছোট খাঁজটি আরও গভীর। প্রাকৃতিক এবং সিন্থেটিক ডিএনএ পলিনিউক্লিওটাইডগুলি সি-ফর্মে যেতে পারে।
| সারণী 1. কিছু ধরণের ডিএনএ কাঠামোর বৈশিষ্ট্য | |||
| সর্পিল প্রকার | ক | খ | জেড |
| সর্পিল পিচ | 0.32 এনএম | 3.38 এনএম | 4.46 এনএম |
| সর্পিল মোচড় | ঠিক | ঠিক | বাম |
| প্রতি টার্ন বেস জোড়া সংখ্যা | 11 | 10 | 12 |
| বেস প্লেন মধ্যে দূরত্ব | 0.256 এনএম | 0.338 এনএম | 0.371 এনএম |
| গ্লাইকোসিডিক বন্ড গঠন | বিরোধী | বিরোধী | বিরোধী সি syn-G |
| Furanose রিং গঠন | C3 "-এন্ডো | C2 "-এন্ডো | C3 "-এন্ডো-জি C2 "-এন্ডো-সি |
| খাঁজ প্রস্থ, ছোট/বড় | 1.11/0.22 এনএম | 0.57/1.17 এনএম | 0.2/0.88 এনএম |
| খাঁজ গভীরতা, ছোট/বড় | 0.26/1.30 এনএম | 0.82/0.85 এনএম | 1.38/0.37 এনএম |
| সর্পিল ব্যাস | 2.3 এনএম | 2.0 এনএম | 1.8 এনএম |
DNA এর কাঠামোগত উপাদান
(অ-প্রামাণিক ডিএনএ কাঠামো)
ডিএনএর কাঠামোগত উপাদানগুলির মধ্যে রয়েছে কিছু বিশেষ ক্রম দ্বারা সীমাবদ্ধ অস্বাভাবিক কাঠামো:
|

DNA এর Z-রূপ 1979 সালে হেক্সানিউক্লিওটাইড ডি(সিজি)3 - অধ্যয়নের সময় আবিষ্কৃত হয়েছিল। এটি MIT এর অধ্যাপক আলেকজান্ডার রিচ এবং তার কর্মীরা খোলেন। জেড-ফর্মটি ডিএনএ-এর সবচেয়ে গুরুত্বপূর্ণ কাঠামোগত উপাদানগুলির মধ্যে একটি হয়ে উঠেছে কারণ এটির গঠন ডিএনএ অঞ্চলে পরিলক্ষিত হয়েছিল যেখানে পিউরিনগুলি পাইরিমিডিনগুলির সাথে বিকল্প হয় (উদাহরণস্বরূপ, 5'-HCHCHC-3'), বা পুনরাবৃত্তি 5' -CHCHCH-3' মিথিলেটেড সাইটোসিন ধারণকারী। জেড-ডিএনএ গঠন ও স্থিতিশীলতার একটি অপরিহার্য শর্ত ছিল এতে সিন-কনফরমেশনে পিউরিন নিউক্লিওটাইডের উপস্থিতি, অ্যান্টি-কনফরমেশনে পাইরিমিডিন বেসগুলির সাথে পর্যায়ক্রমে।
প্রাকৃতিক ডিএনএ অণুগুলি বেশিরভাগই সঠিক B আকারে বিদ্যমান যদি না তাদের মধ্যে (CG)n এর মতো অনুক্রম থাকে। যাইহোক, যদি এই ধরনের ক্রমগুলি ডিএনএর অংশ হয়, তবে এই অঞ্চলগুলি, যখন দ্রবণের আয়নিক শক্তি বা ক্যাটেশনগুলি যা ফসফোডিস্টার ব্যাকবোনে নেতিবাচক চার্জকে নিরপেক্ষ করে, জেড ফর্মে পরিবর্তিত হতে পারে, যখন শৃঙ্খলের অন্যান্য ডিএনএ অঞ্চলগুলি থাকে শাস্ত্রীয় বি-ফর্ম। এই ধরনের পরিবর্তনের সম্ভাবনা নির্দেশ করে যে ডিএনএ ডাবল হেলিক্সের দুটি স্ট্র্যান্ড একটি গতিশীল অবস্থায় রয়েছে এবং একে অপরের সাপেক্ষে মুক্ত হতে পারে, ডান ফর্ম থেকে বাম দিকে চলে যায় এবং এর বিপরীতে। এই স্থিতিশীলতার জৈবিক পরিণতি, যা ডিএনএ কাঠামোর গঠনমূলক রূপান্তরকে অনুমতি দেয়, এখনও সম্পূর্ণরূপে বোঝা যায় নি। এটা বিশ্বাস করা হয় যে জেড-ডিএনএ অঞ্চলগুলি নির্দিষ্ট জিনের অভিব্যক্তি নিয়ন্ত্রণে ভূমিকা পালন করে এবং জেনেটিক পুনর্মিলনে অংশ নেয়।
ডিএনএর জেড-ফর্ম হল একটি বাম-হাতের ডাবল হেলিক্স, যেখানে ফসফোডিস্টার ব্যাকবোনটি অণুর অক্ষ বরাবর জিগজ্যাগ। তাই অণুর নাম (জিগজ্যাগ)-ডিএনএ। Z-DNA হল সবচেয়ে কম পাকানো (প্রতি টার্নে 12 বেস জোড়া) এবং প্রকৃতিতে সবচেয়ে পাতলা। সংলগ্ন নিউক্লিওটাইডগুলির মধ্যে দূরত্ব হল 0.38 nm, কুণ্ডলীর দৈর্ঘ্য 4.56 nm এবং Z-DNA ব্যাস হল 1.8 nm। এছাড়া, চেহারাএই ডিএনএ অণুটি একটি একক খাঁজের উপস্থিতি দ্বারা আলাদা করা হয়।
ডিএনএর জেড-ফর্ম প্রোক্যারিওটিক এবং ইউক্যারিওটিক কোষে পাওয়া গেছে। আজ অবধি, অ্যান্টিবডি পাওয়া গেছে যা DNA-এর Z-ফর্ম এবং B-ফর্মের মধ্যে পার্থক্য করতে পারে। এই অ্যান্টিবডিগুলি ড্রোসোফিলা (ড. মেলানোগাস্টার) লালা গ্রন্থি কোষের দৈত্য ক্রোমোজোমের নির্দিষ্ট অঞ্চলে আবদ্ধ হয়। এই ক্রোমোজোমগুলির অস্বাভাবিক গঠনের কারণে বাঁধাই প্রতিক্রিয়া অনুসরণ করা সহজ, যেখানে ঘন অঞ্চল (ডিস্ক) কম ঘন অঞ্চলের (ইন্টারডিস্ক) সাথে বৈসাদৃশ্য। জেড-ডিএনএ অঞ্চলগুলি ইন্টারডিস্কে অবস্থিত। এটি থেকে এটি অনুসরণ করে যে Z-ফর্মটি আসলে প্রাকৃতিক পরিস্থিতিতে বিদ্যমান, যদিও Z-ফর্মের পৃথক বিভাগগুলির আকার এখনও জানা যায়নি।

(শিফটার) - ডিএনএ-তে সবচেয়ে বিখ্যাত এবং ঘন ঘন ঘটতে থাকা বেস সিকোয়েন্স। একটি প্যালিনড্রোম হল একটি শব্দ বা বাক্যাংশ যা বাম থেকে ডানে এবং একইভাবে বিপরীতে পড়ে। এই ধরনের শব্দ বা বাক্যাংশের উদাহরণ হল: HUT, COSSACK, FLOOD, AND A ROSE FALLED ON AZOR'S PAWS. ডিএনএ-এর অংশগুলিতে প্রয়োগ করা হলে, এই শব্দটি (প্যালিন্ড্রোম) মানে ডান থেকে বাম এবং বাম থেকে ডানে (যেমন "হুট" শব্দের অক্ষরগুলির মতো) চেইন বরাবর নিউক্লিওটাইডগুলির একই পরিবর্তন।
একটি প্যালিনড্রোম দুটি ডিএনএ স্ট্র্যান্ডের সাথে একটি দ্বিতীয়-ক্রম প্রতিসাম্যযুক্ত বেস সিকোয়েন্সের উল্টানো পুনরাবৃত্তির উপস্থিতি দ্বারা চিহ্নিত করা হয়। এই ধরনের ক্রমগুলি, সুস্পষ্ট কারণে, স্ব-পরিপূরক এবং হেয়ারপিন বা ক্রুসিফর্ম কাঠামো গঠনের প্রবণতা (চিত্র)। হেয়ারপিনগুলি নিয়ন্ত্রক প্রোটিনগুলিকে সেই জায়গাটি চিনতে সাহায্য করে যেখানে ক্রোমোজোম ডিএনএর জেনেটিক পাঠ্য অনুলিপি করা হয়।
যে ক্ষেত্রে একই ডিএনএ স্ট্র্যান্ডে একটি উল্টানো পুনরাবৃত্তি উপস্থিত থাকে, এই ধরনের ক্রমকে মিরর পুনরাবৃত্তি বলা হয়। মিরর পুনরাবৃত্তির স্ব-পরিপূরক বৈশিষ্ট্য নেই এবং তাই হেয়ারপিন বা ক্রুসিফর্ম গঠন গঠনে সক্ষম নয়। এই ধরণের সিকোয়েন্সগুলি প্রায় সমস্ত বড় ডিএনএ অণুতে পাওয়া যায় এবং মাত্র কয়েকটি বেস জোড়া থেকে কয়েক হাজার বেস জোড়া পর্যন্ত হতে পারে।
ইউক্যারিওটিক কোষে ক্রুসিফর্ম কাঠামোর আকারে প্যালিনড্রোমের উপস্থিতি প্রমাণিত হয়নি, যদিও ই. কোলি কোষে ভিভোতে বেশ কয়েকটি ক্রুসিফর্ম কাঠামো পাওয়া গেছে। RNA বা সিঙ্গেল-স্ট্র্যান্ডেড ডিএনএ-তে স্ব-পরিপূরক ক্রমগুলির উপস্থিতি হল একটি নির্দিষ্ট স্থানিক কাঠামোতে সমাধানে নিউক্লিক চেইন ভাঁজ করার প্রধান কারণ, যা অনেকগুলি "হেয়ারপিন" গঠনের দ্বারা চিহ্নিত করা হয়।

DNA এর H-রূপ- এটি একটি হেলিক্স যা ডিএনএর তিনটি স্ট্র্যান্ড দ্বারা গঠিত - ডিএনএর ট্রিপল হেলিক্স। এটি ওয়াটসন-ক্রিক ডাবল হেলিক্সের একটি কমপ্লেক্স যার মধ্যে তৃতীয় একক-স্ট্র্যান্ডেড ডিএনএ স্ট্র্যান্ড রয়েছে, যা তার বৃহৎ খাঁজে ফিট করে, তথাকথিত হুগস্টিন জোড়ার গঠনের সাথে।
ডিএনএ ডাবল হেলিক্স যুক্ত করার ফলে এই ধরনের ট্রিপ্লেক্সের গঠন এমনভাবে ঘটে যে এর অর্ধেক অংশ ডাবল হেলিক্সের আকারে থাকে এবং দ্বিতীয় অর্ধেক সংযোগ বিচ্ছিন্ন হয়। এই ক্ষেত্রে, সংযোগ বিচ্ছিন্ন সর্পিলগুলির মধ্যে একটি ডাবল হেলিক্সের প্রথমার্ধের সাথে একটি নতুন কাঠামো তৈরি করে - একটি ট্রিপল হেলিক্স, এবং দ্বিতীয়টি একটি একক-ফিলামেন্ট বিভাগের আকারে অসংগঠিত হতে দেখা যায়। এই কাঠামোগত পরিবর্তনের একটি বৈশিষ্ট্য হল মাধ্যমের pH-এর উপর একটি তীক্ষ্ণ নির্ভরতা, যার প্রোটনগুলি নতুন কাঠামোকে স্থিতিশীল করে। এই বৈশিষ্ট্যের কারণে, নতুন কাঠামোটিকে ডিএনএর এইচ-ফর্ম বলা হয়, যার গঠন হোমোপিউরিন-হোমোপাইরিমিডিন অঞ্চল ধারণকারী সুপারকোয়েলড প্লাজমিডে পাওয়া যায়, যা একটি মিরর পুনরাবৃত্তি।
আরও গবেষণায়, কিছু হোমোপিউরিন-হোমোপাইরিমিডিন ডাবল-স্ট্র্যান্ডেড পলিনিউক্লিওটাইডের কাঠামোগত রূপান্তরের সম্ভাবনা একটি তিন-বিঘ্নিত কাঠামো গঠনের সাথে প্রতিষ্ঠিত হয়েছিল:
- একটি হোমোপিউরিন এবং দুটি হোমোপাইরিমিডিন স্ট্র্যান্ড ( পাই-পু-পাই ট্রিপলেক্স) [হুগস্টিনের মিথস্ক্রিয়া]।
Py-Pu-Py ট্রিপ্লেক্সের উপাদান ব্লকগুলি হল ক্যানোনিকাল আইসোমরফিক CGC+ এবং TAT ট্রায়াড। ট্রিপ্লেক্সের স্থিতিশীলতার জন্য CGC+ ট্রায়াডের প্রোটোনেশন প্রয়োজন, তাই এই ট্রিপ্লেক্সগুলি দ্রবণের pH-এর উপর নির্ভরশীল।
- একটি হোমোপাইরিমিডিন এবং দুটি হোমোপিউরিন স্ট্র্যান্ড ( পাই-পু-পু ট্রিপলেক্স) [উল্টো হুগস্টিন মিথস্ক্রিয়া]।
পাই-পু-পু ট্রিপ্লেক্সের উপাদান ব্লকগুলি হল ক্যানোনিকাল আইসোমরফিক সিজিজি এবং টিএএ ট্রায়াড। Py-Pu-Pu ট্রিপ্লেক্সগুলির একটি অপরিহার্য বৈশিষ্ট্য হল দ্বিগুণ চার্জযুক্ত আয়নগুলির উপস্থিতির উপর তাদের স্থায়িত্বের নির্ভরতা এবং বিভিন্ন অনুক্রমের ট্রিপ্লেক্সগুলিকে স্থিতিশীল করার জন্য বিভিন্ন আয়ন প্রয়োজন। যেহেতু Py-Pu-Pu ট্রিপ্লেক্স গঠনের জন্য তাদের উপাদান নিউক্লিওটাইডগুলির প্রোটোনেশনের প্রয়োজন হয় না, তাই এই জাতীয় ট্রিপ্লেক্স নিরপেক্ষ pH এ থাকতে পারে।
দ্রষ্টব্য: প্রত্যক্ষ এবং বিপরীত Hoogsteen মিথস্ক্রিয়া 1-মিথাইলথাইমিনের প্রতিসাম্য দ্বারা ব্যাখ্যা করা হয়েছে: একটি 180 ° ঘূর্ণন এই সত্যের দিকে পরিচালিত করে যে O4 পরমাণুর স্থানটি O2 পরমাণু দ্বারা দখল করা হয়, যখন হাইড্রোজেন বন্ধনের সিস্টেমটি সংরক্ষিত থাকে।
দুই ধরনের ট্রিপল হেলিক্স আছে:
- সমান্তরাল ট্রিপল হেলিক্স যেখানে তৃতীয় স্ট্র্যান্ডের পোলারিটি ওয়াটসন-ক্রিক ডুপ্লেক্সের হোমোপিউরিন চেইনের সমান
- অ্যান্টিপ্যারালাল ট্রিপল হেলিক্স, যেখানে তৃতীয় এবং হোমোপিউরিন চেইনের মেরুতা বিপরীত।

জি-কোয়াড্রুপ্লেক্স- 4-স্ট্র্যান্ডেড ডিএনএ। এই ধরনের একটি গঠন গঠিত হয় যদি চারটি গুয়ানিন থাকে, যা তথাকথিত জি-কোয়াড্রুপ্লেক্স গঠন করে - চারটি গুয়ানিনের একটি বৃত্তাকার নৃত্য।
এই ধরনের কাঠামো গঠনের সম্ভাবনার প্রথম ইঙ্গিত ওয়াটসন এবং ক্রিক-এর যুগান্তকারী কাজের অনেক আগে প্রাপ্ত হয়েছিল - 1910 সালের প্রথম দিকে। তারপরে জার্মান রসায়নবিদ ইভার ব্যাং আবিষ্কার করেছিলেন যে ডিএনএর একটি উপাদান - গুয়ানোসিক অ্যাসিড - উচ্চ ঘনত্বে জেল তৈরি করে, যখন ডিএনএর অন্যান্য উপাদানগুলিতে এই সম্পত্তি নেই।
1962 সালে, এক্স-রে ডিফ্র্যাকশন পদ্ধতি ব্যবহার করে, এই জেলের কোষ গঠন স্থাপন করা সম্ভব হয়েছিল। এটি চারটি গুয়ানিনের অবশিষ্টাংশের সমন্বয়ে পরিণত হয়েছিল, একে অপরকে একটি বৃত্তে সংযুক্ত করে এবং একটি বৈশিষ্ট্যযুক্ত বর্গক্ষেত্র তৈরি করে। কেন্দ্রে, বন্ধনটি একটি ধাতব আয়ন (Na, K, Mg) দ্বারা সমর্থিত। একই কাঠামো ডিএনএতে তৈরি হতে পারে যদি এতে প্রচুর গুয়ানিন থাকে। এই সমতল স্কোয়ারগুলি (G-quartets) মোটামুটি স্থিতিশীল, ঘন কাঠামো (G-quadruplexes) গঠনের জন্য স্ট্যাক করা হয়।
ডিএনএ-এর চারটি পৃথক স্ট্র্যান্ডকে চার-অসন্তস্থ কমপ্লেক্সে বোনা যেতে পারে, তবে এটি একটি ব্যতিক্রম। প্রায়শই, নিউক্লিক অ্যাসিডের একটি একক স্ট্র্যান্ডকে কেবল একটি গিঁটে বেঁধে রাখা হয়, যা বৈশিষ্ট্যযুক্ত ঘনত্ব তৈরি করে (উদাহরণস্বরূপ, ক্রোমোজোমের প্রান্তে), বা ডাবল-স্ট্র্যান্ডেড ডিএনএ কিছু গুয়ানিন-সমৃদ্ধ স্থানে একটি স্থানীয় চতুর্ভুজ গঠন করে।
সর্বাধিক অধ্যয়ন করা হয় ক্রোমোজোমের প্রান্তে কোয়াড্রুপ্লেক্সের অস্তিত্ব - টেলোমেয়ারে এবং অনকোপ্রোমোটারগুলিতে। যাইহোক, মানুষের ক্রোমোজোমে এই জাতীয় ডিএনএর স্থানীয়করণের সম্পূর্ণ বোঝা এখনও জানা যায়নি।
রৈখিক আকারে ডিএনএর এই সমস্ত অস্বাভাবিক কাঠামো ডিএনএর বি-ফর্মের তুলনায় অস্থির। যাইহোক, ডিএনএ প্রায়শই টপোলজিক্যাল টেনশনের একটি রিং আকারে বিদ্যমান থাকে যখন এটি সুপারকয়লিং নামে পরিচিত। এই অবস্থার অধীনে, নন-ক্যাননিকাল ডিএনএ গঠনগুলি সহজেই গঠিত হয়: জেড-ফর্ম, "ক্রস" এবং "হেয়ারপিনস", এইচ-ফর্ম, গুয়ানিন কোয়াড্রুপ্লেক্স এবং আই-মোটিফ।
- সুপারকোয়েলড ফর্ম - পেন্টোজ-ফসফেট ব্যাকবোনের ক্ষতি ছাড়াই কোষের নিউক্লিয়াস থেকে মুক্তির সময় উল্লেখ করা হয়। এটা supertwisted বন্ধ রিং ফর্ম আছে. সুপারটুইস্টেড অবস্থায়, ডিএনএ ডাবল হেলিক্স অন্তত একবার "নিজেই পেঁচানো" হয়, অর্থাৎ এতে অন্তত একটি সুপারকয়েল থাকে (এটি চিত্র আটের আকার নেয়)।
- ডিএনএ-এর শিথিল অবস্থা - একটি একক বিরতি (এক স্ট্র্যান্ডের বিরতি) দিয়ে পর্যবেক্ষণ করা হয়েছে। এই ক্ষেত্রে, সুপারকয়েলগুলি অদৃশ্য হয়ে যায় এবং ডিএনএ একটি বদ্ধ বলয়ের আকার নেয়।
- ডাবল হেলিক্সের দুটি স্ট্র্যান্ড ভেঙে গেলে ডিএনএর রৈখিক রূপ পরিলক্ষিত হয়।
DNA এর তৃতীয় কাঠামো
DNA এর তৃতীয় কাঠামোএকটি ডাবল-স্ট্র্যান্ডেড অণুর স্থান অতিরিক্ত মোচড়ের ফলে গঠিত হয় - এর সুপারকোলিং। ইউক্যারিওটিক কোষে ডিএনএ অণুর সুপারকয়েলিং প্রোক্যারিওটসের বিপরীতে প্রোটিন সহ কমপ্লেক্স আকারে সঞ্চালিত হয়।
প্রায় সমস্ত ইউক্যারিওটিক ডিএনএ নিউক্লিয়াসের ক্রোমোজোমে অবস্থিত, এর সামান্য পরিমাণ মাইটোকন্ড্রিয়া এবং উদ্ভিদ এবং প্লাস্টিডে পাওয়া যায়। ইউক্যারিওটিক কোষের (মানুষের ক্রোমোজোম সহ) ক্রোমোজোমের প্রধান পদার্থ হল ক্রোমাটিন, এতে ডবল-স্ট্র্যান্ডেড ডিএনএ, হিস্টোন এবং নন-হিস্টোন প্রোটিন থাকে।
ক্রোমাটিনের হিস্টোন প্রোটিন
হিস্টোনগুলি হল সাধারণ প্রোটিন যা 50% পর্যন্ত ক্রোমাটিন তৈরি করে। প্রাণী ও উদ্ভিদের সমস্ত অধ্যয়ন করা কোষে, হিস্টোনের পাঁচটি প্রধান শ্রেণি পাওয়া গেছে: H1, H2A, H2B, H3, H4, আকারে ভিন্নতা, অ্যামিনো অ্যাসিড গঠন এবং চার্জ (সর্বদা ইতিবাচক)।
স্তন্যপায়ী হিস্টোন H1 একটি একক পলিপেপটাইড চেইন নিয়ে গঠিত যাতে প্রায় 215টি অ্যামিনো অ্যাসিড থাকে; অন্যান্য হিস্টোনের আকার 100 থেকে 135 অ্যামিনো অ্যাসিডের মধ্যে পরিবর্তিত হয়। এগুলির সবগুলিই প্রায় 2.5 এনএম ব্যাস সহ একটি গ্লোবুলে সর্পিল এবং পেঁচানো হয়, এতে অস্বাভাবিকভাবে প্রচুর পরিমাণে ইতিবাচক চার্জযুক্ত অ্যামিনো অ্যাসিড লাইসিন এবং আরজিনাইন থাকে। হিস্টোনগুলি অ্যাসিটাইলেটেড, মিথাইলেটেড, ফসফরিলেটেড, পলি(ADP)-রাইবোসিলেট হতে পারে এবং হিস্টোনগুলি H2A এবং H2B সহযোগে ইউবিকুইটিনের সাথে যুক্ত হতে পারে। হিস্টোনগুলির দ্বারা ফাংশনগুলির গঠন এবং কার্যকারিতা গঠনে এই জাতীয় পরিবর্তনগুলির ভূমিকা কী তা এখনও সম্পূর্ণরূপে ব্যাখ্যা করা যায়নি। এটা ধরে নেওয়া হয় যে এটি তাদের ডিএনএর সাথে যোগাযোগ করার এবং জিনের ক্রিয়া নিয়ন্ত্রণের জন্য একটি প্রক্রিয়া সরবরাহ করার ক্ষমতা।
হিস্টোন প্রধানত এর মাধ্যমে ডিএনএর সাথে যোগাযোগ করে আয়নিক বন্ধন(লবণ সেতু) ডিএনএর নেতিবাচক চার্জযুক্ত ফসফেট গ্রুপ এবং ইতিবাচক চার্জযুক্ত লাইসিন এবং আর্জিনাইন হিস্টোন অবশিষ্টাংশের মধ্যে গঠিত।
ক্রোমাটিনের অ-হিস্টোন প্রোটিন
নন-হিস্টোন প্রোটিন, হিস্টোনের বিপরীতে, খুব বৈচিত্র্যময়। ডিএনএ-বাইন্ডিং ননহিস্টোন প্রোটিনের 590টি পর্যন্ত বিভিন্ন ভগ্নাংশ বিচ্ছিন্ন করা হয়েছে। এগুলিকে অ্যাসিডিক প্রোটিনও বলা হয়, যেহেতু অম্লীয় অ্যামিনো অ্যাসিডগুলি তাদের গঠনে প্রাধান্য পায় (তারা পলিয়নিয়ান)। ক্রোমাটিন কার্যকলাপের নির্দিষ্ট নিয়ন্ত্রণ বিভিন্ন অ-হিস্টোন প্রোটিনের সাথে যুক্ত। উদাহরণস্বরূপ, ডিএনএ প্রতিলিপি এবং প্রকাশের জন্য প্রয়োজনীয় এনজাইমগুলি ক্ষণস্থায়ীভাবে ক্রোমাটিনের সাথে আবদ্ধ হতে পারে। অন্যান্য প্রোটিন, যা বিভিন্ন নিয়ন্ত্রক প্রক্রিয়ার সাথে জড়িত বলে, শুধুমাত্র নির্দিষ্ট টিস্যুতে বা পার্থক্যের নির্দিষ্ট পর্যায়ে ডিএনএর সাথে আবদ্ধ হয়। প্রতিটি প্রোটিন ডিএনএ নিউক্লিওটাইডের (ডিএনএ সাইট) একটি নির্দিষ্ট অনুক্রমের পরিপূরক। এই গ্রুপ অন্তর্ভুক্ত:
- সাইট-নির্দিষ্ট জিঙ্ক ফিঙ্গার প্রোটিনের একটি পরিবার। প্রতিটি "জিঙ্ক আঙুল" 5 টি নিউক্লিওটাইড জোড়া নিয়ে গঠিত একটি নির্দিষ্ট সাইটকে স্বীকৃতি দেয়।
- সাইট-নির্দিষ্ট প্রোটিনের একটি পরিবার - হোমোডিমার। ডিএনএর সংস্পর্শে এই জাতীয় প্রোটিনের একটি খণ্ডের একটি "হেলিক্স-টার্ন-হেলিক্স" গঠন রয়েছে।
- উচ্চ গতিশীলতা প্রোটিন (এইচএমজি প্রোটিন - ইংরেজি থেকে, উচ্চ গতিশীলতা জেল প্রোটিন) হল কাঠামোগত এবং নিয়ন্ত্রক প্রোটিনের একটি গ্রুপ যা ক্রমাগত ক্রোমাটিনের সাথে যুক্ত থাকে। তাদের আণবিক ওজন 30 kD এর কম এবং চার্জযুক্ত অ্যামিনো অ্যাসিডের উচ্চ উপাদান দ্বারা চিহ্নিত করা হয়। তাদের কম আণবিক ওজনের কারণে, পলিঅ্যাক্রিলামাইড জেল ইলেক্ট্রোফোরসিসের সময় এইচএমজি প্রোটিনগুলি অত্যন্ত মোবাইল।
- প্রতিলিপি, প্রতিলিপি এবং মেরামতের এনজাইম।
ডিএনএ এবং আরএনএ সংশ্লেষণে জড়িত কাঠামোগত, নিয়ন্ত্রক প্রোটিন এবং এনজাইমগুলির অংশগ্রহণের সাথে, নিউক্লিওসোম থ্রেড প্রোটিন এবং ডিএনএর একটি উচ্চ ঘনীভূত কমপ্লেক্সে রূপান্তরিত হয়। ফলস্বরূপ গঠনটি মূল ডিএনএ অণুর চেয়ে 10,000 গুণ ছোট।

ক্রোমাটিন
ক্রোমাটিন হল পারমাণবিক ডিএনএ সহ প্রোটিনের একটি জটিল এবং অজৈব পদার্থ. বেশিরভাগ ক্রোমাটিন নিষ্ক্রিয়। এতে রয়েছে ঘনবসতিপূর্ণ, ঘনীভূত ডিএনএ। এটি হেটেরোক্রোমাটিন। গঠনমূলক, জেনেটিক্যালি নিষ্ক্রিয় ক্রোমাটিন (উপগ্রহ ডিএনএ) রয়েছে যা প্রকাশ না করা অঞ্চল নিয়ে গঠিত এবং ফ্যাকাল্টেটিভ - বেশ কয়েকটি প্রজন্মের মধ্যে নিষ্ক্রিয়, তবে কিছু নির্দিষ্ট পরিস্থিতিতে প্রকাশ করতে সক্ষম।
সক্রিয় ক্রোমাটিন (ইউক্রোমাটিন) অসংকীর্ণ, অর্থাৎ কম শক্তভাবে প্যাক করা। বিভিন্ন কক্ষে, এর বিষয়বস্তু 2 থেকে 11% পর্যন্ত। মস্তিষ্কের কোষে, এটি সবচেয়ে বেশি - 10-11%, লিভারের কোষে - 3-4 এবং কিডনি - 2-3%। ইউক্রোমাটিনের একটি সক্রিয় প্রতিলিপি আছে। একই সময়ে, এর কাঠামোগত সংস্থা একটি নির্দিষ্ট ধরণের জীবের অন্তর্নিহিত একই ডিএনএ জেনেটিক তথ্যকে বিশেষায়িত কোষে বিভিন্ন উপায়ে ব্যবহার করা সম্ভব করে তোলে।
একটি ইলেক্ট্রন মাইক্রোস্কোপে, ক্রোমাটিনের চিত্রটি পুঁতির অনুরূপ: গোলাকার ঘনত্ব প্রায় 10 এনএম আকারে, ফিলামেন্টাস সেতু দ্বারা পৃথক করা হয়। এই গোলাকার ঘনত্বকে নিউক্লিওসোম বলে। নিউক্লিওসোম হল ক্রোমাটিনের কাঠামোগত একক। প্রতিটি নিউক্লিওসোমে একটি 146 bp দীর্ঘ সুপারকোয়েলড ডিএনএ সেগমেন্টের ক্ষত থাকে যা প্রতি নিউক্লিওসোম কোরে 1.75টি বাম বাঁক তৈরি করে। নিউক্লিওসোমাল কোর হল একটি হিস্টোন অক্টামার যা হিস্টোন H2A, H2B, H3 এবং H4 নিয়ে গঠিত, প্রতিটি ধরণের দুটি অণু (চিত্র 9), যা দেখতে 11 এনএম ব্যাস এবং 5.7 এনএম পুরুত্ব সহ একটি ডিস্কের মতো। পঞ্চম হিস্টোন, H1, নিউক্লিওসোমাল কোরের অংশ নয় এবং হিস্টোন অক্টেমারের চারপাশে ডিএনএ ঘুরানোর প্রক্রিয়ার সাথে জড়িত নয়। এটি ডিএনএ-র সাথে সেই বিন্দুতে যোগাযোগ করে যেখানে ডাবল হেলিক্স নিউক্লিওসোমাল কোরে প্রবেশ করে এবং প্রস্থান করে। এগুলি হল ডিএনএর ইন্টারকোর (লিঙ্কার) বিভাগ, যার দৈর্ঘ্য 40 থেকে 50 নিউক্লিওটাইড জোড়া কোষের ধরণের উপর নির্ভর করে পরিবর্তিত হয়। ফলস্বরূপ, নিউক্লিওসোমের অংশ ডিএনএ খণ্ডের দৈর্ঘ্যও পরিবর্তিত হয় (186 থেকে 196 নিউক্লিওটাইড জোড়া পর্যন্ত)।
নিউক্লিওসোমে ডিএনএর প্রায় 90% থাকে, বাকি অংশ লিঙ্কার। এটি বিশ্বাস করা হয় যে নিউক্লিওসোমগুলি "নীরব" ক্রোমাটিনের টুকরা, যখন লিঙ্কার সক্রিয় থাকে। যাইহোক, নিউক্লিওসোমগুলি উন্মোচিত হতে পারে এবং রৈখিক হতে পারে। উন্মোচিত নিউক্লিওসোমগুলি ইতিমধ্যে সক্রিয় ক্রোমাটিন। এটি কাঠামোর উপর ফাংশনের নির্ভরতা স্পষ্টভাবে দেখায়। এটি অনুমান করা যেতে পারে যে গ্লোবুলার নিউক্লিওসোমগুলির সংমিশ্রণে ক্রোমাটিন যত বেশি, এটি তত কম সক্রিয়। স্পষ্টতই, বিভিন্ন কোষে বিশ্রামের ক্রোমাটিনের অসম অনুপাত এই জাতীয় নিউক্লিওসোমের সংখ্যার সাথে যুক্ত।
ইলেক্ট্রন মাইক্রোস্কোপিক ফটোগ্রাফগুলিতে, বিচ্ছিন্নতার অবস্থা এবং প্রসারিত হওয়ার মাত্রার উপর নির্ভর করে, ক্রোমাটিন কেবল ঘন হওয়া - নিউক্লিওসোমের "জপমালা" সহ একটি দীর্ঘ থ্রেড হিসাবে নয়, বরং একটি খাটো এবং ঘন ফাইব্রিল (ফাইবার) হিসাবেও দেখা যেতে পারে যার ব্যাস রয়েছে। 30 এনএম, যার গঠনটি ডিএনএ এবং হিস্টোন এইচ 3 এর লিঙ্কার অঞ্চলের সাথে সম্পর্কিত হিস্টোন এইচ 1 এর মিথস্ক্রিয়া চলাকালীন পরিলক্ষিত হয়, যা 30 এনএম ব্যাস সহ একটি সোলেনয়েড গঠনের সাথে প্রতি টার্নে ছয়টি নিউক্লিওসোমের হেলিক্সের অতিরিক্ত মোচড়ের দিকে পরিচালিত করে। . এই ক্ষেত্রে, হিস্টোন প্রোটিন বেশ কয়েকটি জিনের প্রতিলিপিতে হস্তক্ষেপ করতে পারে এবং এইভাবে তাদের কার্যকলাপ নিয়ন্ত্রণ করতে পারে।
উপরে বর্ণিত হিস্টোনগুলির সাথে ডিএনএ-এর মিথস্ক্রিয়াগুলির ফলস্বরূপ, 2 এনএম এবং 57 এনএম দৈর্ঘ্যের গড় ব্যাস সহ 186টি বেস জোড়ার ডিএনএ ডাবল হেলিক্সের একটি অংশ 10 এনএম এবং দৈর্ঘ্যের ব্যাস সহ একটি হেলিক্সে পরিণত হয়। 5 এনএম এর। 30 এনএম ব্যাসের একটি ফাইবারে এই হেলিক্সের পরবর্তী সংকোচনের সাথে, ঘনীভবনের মাত্রা আরও ছয় গুণ বৃদ্ধি পায়।
শেষ পর্যন্ত, পাঁচটি হিস্টোন সহ ডিএনএ ডুপ্লেক্সের প্যাকেজিং 50-গুণ ডিএনএ ঘনীভূত হয়। যাইহোক, এমনকি এত উচ্চ মাত্রার ঘনীভবনও মেটাফেজ ক্রোমোসোমে প্রায় 50,000-100,000-গুণ ডিএনএ কম্প্যাকশন ব্যাখ্যা করতে পারে না। দুর্ভাগ্যবশত, মেটাফেজ ক্রোমোজোম পর্যন্ত ক্রোমাটিনের আরও প্যাকিংয়ের বিশদ বিবরণ এখনও জানা যায়নি; অতএব, এই প্রক্রিয়াটির শুধুমাত্র সাধারণ বৈশিষ্ট্যগুলি বিবেচনা করা যেতে পারে।
ক্রোমোজোমে ডিএনএ কমপ্যাকশনের মাত্রা
প্রতিটি ডিএনএ অণু একটি পৃথক ক্রোমোজোমে প্যাকেজ করা হয়। ডিপ্লোয়েড মানব কোষে 46টি ক্রোমোজোম থাকে, যা কোষের নিউক্লিয়াসে অবস্থিত। একটি কোষের সমস্ত ক্রোমোজোমের ডিএনএর মোট দৈর্ঘ্য 1.74 মিটার, তবে যে নিউক্লিয়াসে ক্রোমোজোমগুলি প্যাক করা হয় তার ব্যাস লক্ষ লক্ষ গুণ ছোট। কোষের নিউক্লিয়াসে ক্রোমোসোম এবং ক্রোমোজোমে ডিএনএর এই ধরনের কম্প্যাক্ট প্যাকিং বিভিন্ন হিস্টোন এবং নন-হিস্টোন প্রোটিন ডিএনএর সাথে একটি নির্দিষ্ট ক্রমানুসারে মিথস্ক্রিয়া করে (উপরে দেখুন)। ক্রোমোজোমে ডিএনএ-এর কম্প্যাকশন এর রৈখিক মাত্রা প্রায় 10,000 গুণ কমানো সম্ভব করে - শর্তসাপেক্ষে 5 সেমি থেকে 5 মাইক্রন পর্যন্ত। কম্প্যাক্টাইজেশনের বিভিন্ন স্তর রয়েছে (চিত্র 10)।


- DNA ডাবল হেলিক্স হল একটি নেতিবাচক চার্জযুক্ত অণু যার ব্যাস 2 nm এবং দৈর্ঘ্য কয়েক সেমি।
- নিউক্লিওসোমাল স্তর- ক্রোমাটিন একটি ইলেক্ট্রন মাইক্রোস্কোপে "পুঁতি" - নিউক্লিওসোম - "একটি সুতার উপর" এর চেইন হিসাবে দেখায়। নিউক্লিওসোম হল একটি সার্বজনীন কাঠামোগত একক যা ইউক্রোমাটিন এবং হেটেরোক্রোমাটিন উভয় ক্ষেত্রেই পাওয়া যায়, ইন্টারফেজ নিউক্লিয়াস এবং মেটাফেজ ক্রোমোজোমে।
কম্প্যাকশনের নিউক্লিওসোমাল স্তর বিশেষ প্রোটিন - হিস্টোন দ্বারা সরবরাহ করা হয়। আটটি ইতিবাচক চার্জযুক্ত হিস্টোন ডোমেন নিউক্লিওসোমের মূল (কোর) গঠন করে যার চারপাশে নেতিবাচক চার্জযুক্ত ডিএনএ অণুটি ক্ষতবিক্ষত থাকে। এটি 7 এর একটি ফ্যাক্টর দ্বারা একটি সংক্ষিপ্তকরণ দেয়, যখন ব্যাস 2 থেকে 11 nm পর্যন্ত বৃদ্ধি পায়।
- সোলেনয়েড স্তর
ক্রোমোজোম সংস্থার সোলেনয়েড স্তরটি নিউক্লিওসোমাল ফিলামেন্টের মোচড়ের দ্বারা চিহ্নিত করা হয় এবং এটি থেকে 20-35 এনএম ব্যাসের ঘন ফাইব্রিলগুলির গঠন - সোলেনয়েড বা সুপারবিডস। সোলেনয়েড পিচ 11 এনএম, এবং প্রতি টার্নে প্রায় 6-10 নিউক্লিওসোম রয়েছে। সোলেনয়েড প্যাকিংকে সুপারবিড প্যাকিংয়ের চেয়ে বেশি সম্ভাব্য বলে মনে করা হয়, যার মতে 20-35 এনএম ব্যাস সহ একটি ক্রোমাটিন ফাইব্রিল হল গ্রানুলের একটি চেইন, বা সুপারবিড, যার প্রতিটিতে আটটি নিউক্লিওসোম থাকে। সোলেনয়েড স্তরে, ডিএনএর রৈখিক আকার 6-10 বার হ্রাস পায়, ব্যাস 30 এনএম পর্যন্ত বৃদ্ধি পায়।
- লুপ স্তর
লুপ স্তরটি অ-হিস্টোন সাইট-নির্দিষ্ট ডিএনএ-বাইন্ডিং প্রোটিন দ্বারা সরবরাহ করা হয় যা নির্দিষ্ট ডিএনএ ক্রমগুলিকে চিনতে এবং আবদ্ধ করে, প্রায় 30-300 kb এর লুপ তৈরি করে। লুপ জিনের অভিব্যক্তি নিশ্চিত করে, যেমন লুপটি কেবল একটি কাঠামোগত নয়, একটি কার্যকরী গঠনও। এই স্তরে সংক্ষিপ্তকরণ 20-30 বার ঘটে। ব্যাস 300 এনএম পর্যন্ত বৃদ্ধি পায়। উভচর oocytes মধ্যে লুপ-সদৃশ "ল্যাম্পব্রাশ" কাঠামো সাইটোলজিক্যাল প্রস্তুতিতে দেখা যায়। এই লুপগুলি সুপারকোয়েল করা এবং ডিএনএ ডোমেনের প্রতিনিধিত্ব করে, সম্ভবত ক্রোমাটিন ট্রান্সক্রিপশন এবং প্রতিলিপির ইউনিটগুলির সাথে সম্পর্কিত। নির্দিষ্ট প্রোটিনগুলি লুপগুলির ভিত্তিগুলি এবং সম্ভবত, তাদের কিছু অভ্যন্তরীণ অঞ্চলগুলিকে ঠিক করে। লুপ-সদৃশ ডোমেন সংস্থাটি মেটাফেজ ক্রোমোজোমের ক্রোমাটিনকে উচ্চতর অর্ডারের হেলিকাল কাঠামোতে ভাঁজ করার সুবিধা দেয়।
- ডোমেইন স্তর
ক্রোমোজোম সংস্থার ডোমেন স্তর যথেষ্ট অধ্যয়ন করা হয়নি। এই স্তরে, লুপ ডোমেনগুলির গঠন লক্ষ করা যায় - 25-30 এনএম পুরু ফিলামেন্টের কাঠামো, যাতে 60% প্রোটিন, 35% ডিএনএ এবং 5% আরএনএ থাকে, কোষ চক্রের সমস্ত ধাপে কার্যত অদৃশ্য থাকে। মাইটোসিসের ব্যতিক্রম এবং কিছুটা এলোমেলোভাবে কোষের নিউক্লিয়াসে বিতরণ করা হয়। উভচর oocytes মধ্যে লুপ-সদৃশ "ল্যাম্পব্রাশ" কাঠামো সাইটোলজিক্যাল প্রস্তুতিতে দেখা যায়।
লুপ ডোমেনগুলি তথাকথিত অন্তর্নির্মিত সংযুক্তি সাইটগুলিতে ইন্ট্রানিউক্লিয়ার প্রোটিন ম্যাট্রিক্সের সাথে তাদের বেসের সাথে সংযুক্ত থাকে, প্রায়শই MAR / SAR সিকোয়েন্স হিসাবে উল্লেখ করা হয় (MAR, ইংরেজি ম্যাট্রিক্স সম্পর্কিত অঞ্চল থেকে; SAR, ইংরেজি স্ক্যাফোল্ড সংযুক্তি অঞ্চল থেকে) - ডিএনএ কয়েকশ লম্বা বেস জোড়াকে খণ্ডিত করে যা A/T বেস জোড়ার উচ্চ বিষয়বস্তু (>65%) দ্বারা চিহ্নিত করা হয়। প্রতিটি ডোমেইনের প্রতিলিপির একটি একক উত্স আছে বলে মনে হয় এবং একটি স্বায়ত্তশাসিত সুপারকোল্ড ইউনিট হিসাবে কাজ করে। যেকোনো লুপ ডোমেনে অনেক ট্রান্সক্রিপশন ইউনিট থাকে, যার কার্যকারিতা সমন্বিত হওয়ার সম্ভাবনা থাকে - পুরো ডোমেনটি হয় সক্রিয় বা নিষ্ক্রিয় অবস্থায় থাকে।
ডোমেন স্তরে, ক্রোমাটিনের অনুক্রমিক প্যাকিংয়ের ফলে, ডিএনএর রৈখিক মাত্রা প্রায় 200 গুণ (700 এনএম) হ্রাস পায়।
- ক্রোমোজোম স্তর
উপরে ক্রোমোজোম স্তরনন-হিস্টোন প্রোটিনের অক্ষীয় কাঠামোর চারপাশে লুপ ডোমেনের কম্প্যাকশনের সাথে মেটাফেজ ক্রোমোজোমে প্রোফেস ক্রোমোজোমের ঘনীভবন ঘটে। এই সুপারকোলিং কোষের সমস্ত H1 অণুর ফসফোরিলেশনের সাথে থাকে। ফলস্বরূপ, মেটাফেজ ক্রোমোজোমকে একটি আঁটসাঁট সর্পিল মধ্যে কুণ্ডলীকৃত ঘন বস্তাবন্দী সোলেনয়েড লুপ হিসাবে চিত্রিত করা যেতে পারে। একটি সাধারণ মানব ক্রোমোজোমে 2600টি পর্যন্ত লুপ থাকতে পারে। এই ধরনের কাঠামোর পুরুত্ব 1400 এনএম (দুটি ক্রোমাটিড) এ পৌঁছায়, যখন ডিএনএ অণু 104 বার ছোট হয়, অর্থাৎ 5 সেমি প্রসারিত DNA থেকে 5 µm পর্যন্ত।
ক্রোমোজোমের কাজ
এক্সট্রা ক্রোমোসোমাল মেকানিজমের সাথে মিথস্ক্রিয়ায়, ক্রোমোজোম প্রদান করে
- বংশগত তথ্য সংরক্ষণ
- সেলুলার সংগঠন তৈরি এবং বজায় রাখতে এই তথ্য ব্যবহার করে
- বংশগত তথ্য পড়ার নিয়ন্ত্রণ
- জেনেটিক উপাদানের স্ব-প্রতিলিপি
- মাতৃকোষ থেকে কন্যা কোষে জেনেটিক উপাদান স্থানান্তর।
প্রমাণ আছে যে একটি ক্রোমাটিন অঞ্চল সক্রিয় করার উপর, যেমন ট্রান্সক্রিপশনের সময়, হিস্টোন H1 বিপরীতভাবে এটি থেকে প্রথমে সরানো হয় এবং তারপরে হিস্টোন অক্টেট। এর ফলে ক্রোমাটিনের ঘনীভবন ঘটে, একটি 30-nm ক্রোমাটিন ফাইব্রিলের ধারাবাহিকভাবে 10-nm ফিলামেন্টে রূপান্তরিত হয় এবং এটি আরও মুক্ত ডিএনএ অঞ্চলে উদ্ভাসিত হয়, যেমন নিউক্লিওসোমাল কাঠামোর ক্ষতি।
আমরা সবাই জানি যে একজন ব্যক্তির চেহারা, কিছু অভ্যাস এমনকি রোগও উত্তরাধিকারসূত্রে প্রাপ্ত হয়। একটি জীব সম্পর্কে এই সমস্ত তথ্য জিনে এনকোড করা হয়। তাহলে এই কুখ্যাত জিনগুলি দেখতে কেমন, তারা কীভাবে কাজ করে এবং তারা কোথায় অবস্থিত?
সুতরাং, যে কোনও ব্যক্তি বা প্রাণীর সমস্ত জিনের বাহক হল ডিএনএ। এই যৌগটি 1869 সালে জোহান ফ্রেডরিখ মিশার আবিষ্কার করেছিলেন।রাসায়নিকভাবে, ডিএনএ হল ডিঅক্সিরাইবোনিউক্লিক অ্যাসিড। এটার মানে কি? কিভাবে এই অ্যাসিড আমাদের গ্রহের সমস্ত জীবনের জেনেটিক কোড বহন করে?
ডিএনএ কোথায় অবস্থিত তা দেখে শুরু করা যাক। মানব কোষে অনেক অর্গানেল রয়েছে যা বিভিন্ন কার্য সম্পাদন করে। ডিএনএ নিউক্লিয়াসে অবস্থিত। নিউক্লিয়াস হল একটি ছোট অর্গানেল যা একটি বিশেষ ঝিল্লি দ্বারা বেষ্টিত যা সমস্ত জেনেটিক উপাদান - ডিএনএ সঞ্চয় করে।
ডিএনএ অণুর গঠন কী?
প্রথমে দেখা যাক ডিএনএ কি? ডিএনএ একটি খুব দীর্ঘ অণু যা গঠনগত উপাদান - নিউক্লিওটাইড নিয়ে গঠিত। নিউক্লিওটাইডের 4 প্রকার রয়েছে - অ্যাডেনিন (এ), থাইমিন (টি), গুয়ানিন (জি) এবং সাইটোসিন (সি)। নিউক্লিওটাইডের চেইন পরিকল্পিতভাবে এইরকম দেখায়: GGAATTSTAAG.... নিউক্লিওটাইডের এই ক্রমটি হল DNA চেইন।ডিএনএর গঠন প্রথম 1953 সালে জেমস ওয়াটসন এবং ফ্রান্সিস ক্রিক দ্বারা পাঠোদ্ধার করা হয়েছিল।
একটি ডিএনএ অণুতে, নিউক্লিওটাইডের দুটি শৃঙ্খল রয়েছে যা একে অপরের চারপাশে হেলালিভাবে পেঁচানো থাকে। কীভাবে এই নিউক্লিওটাইড চেইনগুলি একসাথে লেগে থাকে এবং একটি সর্পিল হয়ে যায়? এই ঘটনাটি পরিপূরকতার সম্পত্তির কারণে। পরিপূরকতা মানে শুধুমাত্র নির্দিষ্ট নিউক্লিওটাইড (পরিপূরক) দুটি চেইনে একে অপরের বিপরীত হতে পারে। সুতরাং, বিপরীত অ্যাডেনিন সবসময় থাইমিন হয়, এবং বিপরীত গুয়ানিন সবসময় শুধুমাত্র সাইটোসিন হয়। এইভাবে, গুয়ানিন সাইটোসিনের সাথে পরিপূরক, এবং থাইমিনের সাথে অ্যাডেনিন। এই ধরনের জোড়া নিউক্লিওটাইডগুলি একে অপরের বিপরীতে বিভিন্ন শৃঙ্খলে পরিপূরক বলা হয়।
 এটি পরিকল্পনাগতভাবে নিম্নরূপ উপস্থাপন করা যেতে পারে:
এটি পরিকল্পনাগতভাবে নিম্নরূপ উপস্থাপন করা যেতে পারে:
জি - সি
টি - এ
টি - এ
গ - জি
এই পরিপূরক জোড়া A - T এবং G - C ফর্ম রাসায়নিক বন্ধনজোড়ার নিউক্লিওটাইডের মধ্যে, এবং G এবং C-এর মধ্যে বন্ধন A এবং T-এর মধ্যে থেকে শক্তিশালী। বন্ধনটি পরিপূরক ঘাঁটির মধ্যে কঠোরভাবে গঠিত হয়, অর্থাৎ, অ-পরিপূরক G এবং A-এর মধ্যে একটি বন্ধন তৈরি করা অসম্ভব।
ডিএনএর "প্যাকেজিং", ডিএনএর একটি স্ট্র্যান্ড কীভাবে ক্রোমোজোমে পরিণত হয়?
কেন ডিএনএর এই নিউক্লিওটাইড চেইনগুলি একে অপরের চারপাশে মোচড় দেয়? কেন এই প্রয়োজন? আসল বিষয়টি হল নিউক্লিওটাইডের সংখ্যা বিশাল এবং এই ধরনের দীর্ঘ চেইনগুলিকে মিটমাট করার জন্য আপনার অনেক জায়গার প্রয়োজন। এই কারণে, ডিএনএর দুটি স্ট্র্যান্ডের চারপাশে একটি সর্পিল মোচড় রয়েছে। এই ঘটনাসর্পিলকরণ বলা হয়। সর্পিলকরণের ফলে, ডিএনএ চেইন 5-6 বার ছোট হয়।কিছু ডিএনএ অণু সক্রিয়ভাবে শরীর দ্বারা ব্যবহৃত হয়, অন্যগুলি খুব কমই ব্যবহৃত হয়। এই ধরনের খুব কমই ব্যবহৃত ডিএনএ অণুগুলি, হেলিকালাইজেশন ছাড়াও, আরও কমপ্যাক্ট "প্যাকেজিং" এর মধ্য দিয়ে যায়। এই ধরনের একটি কমপ্যাক্ট প্যাকেজকে সুপারকোলিং বলা হয় এবং ডিএনএ স্ট্র্যান্ডকে 25-30 বার ছোট করে!
ডিএনএ হেলিক্স কিভাবে প্যাকেজ করা হয়?
 সুপারকোলিংয়ের জন্য, হিস্টোন প্রোটিন ব্যবহার করা হয়, যার চেহারা এবং গঠন একটি রড বা থ্রেডের স্পুল। ডিএনএর সর্পিল স্ট্র্যান্ডগুলি এই "কয়েল" - হিস্টোন প্রোটিনের উপর ক্ষতবিক্ষত হয়। এইভাবে, লম্বা ফিলামেন্ট খুব কম্প্যাক্টভাবে প্যাক হয়ে যায় এবং খুব কম জায়গা নেয়।
সুপারকোলিংয়ের জন্য, হিস্টোন প্রোটিন ব্যবহার করা হয়, যার চেহারা এবং গঠন একটি রড বা থ্রেডের স্পুল। ডিএনএর সর্পিল স্ট্র্যান্ডগুলি এই "কয়েল" - হিস্টোন প্রোটিনের উপর ক্ষতবিক্ষত হয়। এইভাবে, লম্বা ফিলামেন্ট খুব কম্প্যাক্টভাবে প্যাক হয়ে যায় এবং খুব কম জায়গা নেয়। যদি এক বা অন্য ডিএনএ অণু ব্যবহার করার প্রয়োজন হয় তবে "আনওয়াইন্ডিং" প্রক্রিয়াটি ঘটে, অর্থাৎ, ডিএনএ থ্রেডটি "কুণ্ডলী" থেকে "আনওয়াউন্ড" হয় - হিস্টোন প্রোটিন (যদি এটিতে ক্ষত থাকে) এবং এটি থেকে খুলে যায়। হেলিক্স দুটি সমান্তরাল শৃঙ্খলে। এবং যখন ডিএনএ অণুটি এমন একটি অপরিবর্তিত অবস্থায় থাকে, তখন এটি থেকে প্রয়োজনীয় জেনেটিক তথ্য পড়া যায়। তাছাড়া, জেনেটিক তথ্যের পঠন শুধুমাত্র অপরিবর্তিত ডিএনএ স্ট্র্যান্ড থেকে ঘটে!
সুপারকোয়েলড ক্রোমোজোমের সমষ্টিকে বলা হয় heterochromatin, এবং তথ্য পড়ার জন্য উপলব্ধ ক্রোমোজোম - ইউক্রোমাটিন.
জিন কি, ডিএনএর সাথে তাদের সম্পর্ক কি?
এখন দেখা যাক জিন কি। জানা যায়, রক্তের গ্রুপ, চোখের রঙ, চুল, ত্বক এবং আমাদের শরীরের আরও অনেক বৈশিষ্ট্য নির্ধারণ করে এমন জিন রয়েছে। একটি জিন হল ডিএনএর একটি কঠোরভাবে সংজ্ঞায়িত বিভাগ, যা একটি কঠোরভাবে সংজ্ঞায়িত সংমিশ্রণে সাজানো নির্দিষ্ট সংখ্যক নিউক্লিওটাইড নিয়ে গঠিত। ডিএনএ-র একটি কঠোরভাবে সংজ্ঞায়িত বিভাগে অবস্থানের অর্থ হল একটি নির্দিষ্ট জিনের স্থান রয়েছে এবং এই স্থানটি পরিবর্তন করা অসম্ভব। এই জাতীয় তুলনা করা উপযুক্ত: একজন ব্যক্তি একটি নির্দিষ্ট রাস্তায়, একটি নির্দিষ্ট বাড়ি এবং অ্যাপার্টমেন্টে থাকেন এবং একজন ব্যক্তি নির্বিচারে অন্য বাড়ি, অ্যাপার্টমেন্ট বা অন্য রাস্তায় যেতে পারে না। একটি জিনে একটি নির্দিষ্ট সংখ্যক নিউক্লিওটাইড মানে প্রতিটি জিনের একটি নির্দিষ্ট সংখ্যক নিউক্লিওটাইড থাকে এবং কম বা বেশি হতে পারে না। উদাহরণস্বরূপ, জিন এনকোডিং ইনসুলিন উত্পাদন 60 বেস জোড়া দীর্ঘ; হরমোন অক্সিটোসিনের উৎপাদন এনকোডিং জিন 370 bp. একটি কঠোর নিউক্লিওটাইড ক্রম প্রতিটি জিনের জন্য অনন্য এবং কঠোরভাবে সংজ্ঞায়িত। উদাহরণস্বরূপ, AATAATA ক্রম হল একটি জিনের একটি খণ্ড যা ইনসুলিন উৎপাদনের জন্য কোড করে। ইনসুলিন প্রাপ্ত করার জন্য, ঠিক এই জাতীয় ক্রম ব্যবহার করা হয়; প্রাপ্ত করার জন্য, উদাহরণস্বরূপ, অ্যাড্রেনালিন, নিউক্লিওটাইডগুলির একটি ভিন্ন সংমিশ্রণ ব্যবহার করা হয়। এটা বোঝা গুরুত্বপূর্ণ যে শুধুমাত্র নিউক্লিওটাইডের একটি নির্দিষ্ট সংমিশ্রণ একটি নির্দিষ্ট "পণ্য" (অ্যাড্রেনালিন, ইনসুলিন, ইত্যাদি) এনকোড করে। নির্দিষ্ট সংখ্যক নিউক্লিওটাইডের এমন একটি অনন্য সংমিশ্রণ, "তার জায়গায়" দাঁড়িয়ে - এটি হল জিন.
জিন ছাড়াও, তথাকথিত "নন-কোডিং সিকোয়েন্স" ডিএনএ চেইনে অবস্থিত। এই ধরনের নন-কোডিং নিউক্লিওটাইড ক্রমগুলি জিনের কার্যকারিতা নিয়ন্ত্রণ করে, ক্রোমোজোম সর্পিলকরণে সহায়তা করে এবং একটি জিনের শুরু এবং শেষ বিন্দু চিহ্নিত করে। যাইহোক, আজ অবধি, বেশিরভাগ নন-কোডিং সিকোয়েন্সের ভূমিকা অস্পষ্ট রয়ে গেছে।
ক্রোমোজোম কি? যৌন ক্রোমোজোম
একজন ব্যক্তির জিনের সামগ্রিকতাকে জিনোম বলা হয়। স্বাভাবিকভাবেই, পুরো জিনোম একটি একক ডিএনএতে প্যাক করা যায় না। জিনোমটি 46 জোড়া ডিএনএ অণুতে বিভক্ত। এক জোড়া ডিএনএ অণুকে ক্রোমোজোম বলে। সুতরাং এই ক্রোমোজোমগুলিই একজন ব্যক্তির 46 টি টুকরা থাকে। প্রতিটি ক্রোমোজোম জিনের একটি কঠোরভাবে সংজ্ঞায়িত সেট বহন করে, উদাহরণস্বরূপ, 18 তম ক্রোমোজোমে চোখের রঙের এনকোডিং জিন রয়েছে, ইত্যাদি। ক্রোমোজোমগুলি একে অপরের থেকে দৈর্ঘ্য এবং আকারে আলাদা। সবচেয়ে সাধারণ ফর্মগুলি X বা Y আকারে, তবে অন্যান্যগুলিও রয়েছে। একজন ব্যক্তির একই আকৃতির দুটি ক্রোমোজোম থাকে, যাকে জোড়া (জোড়া) বলা হয়। এই ধরনের পার্থক্যের সাথে সংযোগে, সমস্ত জোড়াযুক্ত ক্রোমোজোম সংখ্যাযুক্ত - 23 জোড়া রয়েছে। এর মানে হল এক জোড়া ক্রোমোজোম #1, জোড়া #2, #3, ইত্যাদি। একটি নির্দিষ্ট বৈশিষ্ট্যের জন্য দায়ী প্রতিটি জিন একই ক্রোমোজোমে অবস্থিত। বিশেষজ্ঞদের জন্য আধুনিক ম্যানুয়ালগুলিতে, জিনের স্থানীয়করণ নির্দেশিত হতে পারে, উদাহরণস্বরূপ, নিম্নরূপ: ক্রোমোজোম 22, দীর্ঘ বাহু।ক্রোমোজোমের মধ্যে পার্থক্য কি?
 আর কিভাবে ক্রোমোজোম একে অপরের থেকে আলাদা? দীর্ঘ বাহু শব্দটির অর্থ কী? X-আকৃতির ক্রোমোজোম ধরা যাক। ডিএনএ স্ট্র্যান্ডের ক্রসিং মাঝখানে (X) কঠোরভাবে ঘটতে পারে, বা এটি কেন্দ্রীয়ভাবে ঘটতে পারে না। যখন ডিএনএ স্ট্র্যান্ডের এমন একটি ছেদ কেন্দ্রীয়ভাবে ঘটে না, তখন ছেদ বিন্দুর সাপেক্ষে, কিছু প্রান্তগুলি যথাক্রমে লম্বা হয়, অন্যগুলি ছোট হয়। এই ধরনের দীর্ঘ প্রান্তগুলিকে সাধারণত ক্রোমোজোমের দীর্ঘ বাহু এবং ছোট প্রান্তকে যথাক্রমে ছোট বাহু বলা হয়। ওয়াই-আকৃতির ক্রোমোজোমগুলি বেশিরভাগই লম্বা বাহু দ্বারা দখল করা হয় এবং ছোটগুলি খুব ছোট (এগুলি এমনকি পরিকল্পিত ছবিতেও নির্দেশিত নয়)।
আর কিভাবে ক্রোমোজোম একে অপরের থেকে আলাদা? দীর্ঘ বাহু শব্দটির অর্থ কী? X-আকৃতির ক্রোমোজোম ধরা যাক। ডিএনএ স্ট্র্যান্ডের ক্রসিং মাঝখানে (X) কঠোরভাবে ঘটতে পারে, বা এটি কেন্দ্রীয়ভাবে ঘটতে পারে না। যখন ডিএনএ স্ট্র্যান্ডের এমন একটি ছেদ কেন্দ্রীয়ভাবে ঘটে না, তখন ছেদ বিন্দুর সাপেক্ষে, কিছু প্রান্তগুলি যথাক্রমে লম্বা হয়, অন্যগুলি ছোট হয়। এই ধরনের দীর্ঘ প্রান্তগুলিকে সাধারণত ক্রোমোজোমের দীর্ঘ বাহু এবং ছোট প্রান্তকে যথাক্রমে ছোট বাহু বলা হয়। ওয়াই-আকৃতির ক্রোমোজোমগুলি বেশিরভাগই লম্বা বাহু দ্বারা দখল করা হয় এবং ছোটগুলি খুব ছোট (এগুলি এমনকি পরিকল্পিত ছবিতেও নির্দেশিত নয়)। ক্রোমোজোমের আকার ওঠানামা করে: সবচেয়ে বড় হল জোড়া নং 1 এবং নং 3 এর ক্রোমোজোম, 17 নং, 19 নং জোড়ার সবচেয়ে ছোট ক্রোমোজোম।
আকৃতি এবং আকার ছাড়াও, ক্রোমোজোমগুলি তাদের কার্যকারিতায় ভিন্ন। 23 জোড়ার মধ্যে 22 জোড়া সোমাটিক এবং 1 জোড়া যৌন। এর মানে কী? সোম্যাটিক ক্রোমোজোমগুলি একজন ব্যক্তির সমস্ত বাহ্যিক লক্ষণ, তার বৈশিষ্ট্যগুলি নির্ধারণ করে আচরণগত প্রতিক্রিয়া, বংশগত সাইকোটাইপ, অর্থাৎ, প্রতিটি পৃথক ব্যক্তির সমস্ত বৈশিষ্ট্য এবং বৈশিষ্ট্য। এক জোড়া যৌন ক্রোমোজোম একজন ব্যক্তির লিঙ্গ নির্ধারণ করে: পুরুষ বা মহিলা। মানুষের সেক্স ক্রোমোজোম দুই ধরনের - X (X) এবং Y (Y)। যদি তারা XX (x - x) হিসাবে মিলিত হয় - এটি একজন মহিলা, এবং যদি XY (x - y) - আমাদের সামনে একজন পুরুষ আছে।
বংশগত রোগ এবং ক্রোমোজোমের ক্ষতি
যাইহোক, জিনোমের "ভাঙ্গন" আছে, তারপর জিনগত রোগ মানুষের মধ্যে সনাক্ত করা হয়। উদাহরণস্বরূপ, যখন দুটির পরিবর্তে 21 জোড়া ক্রোমোজোমে তিনটি ক্রোমোজোম থাকে, তখন একজন ব্যক্তি ডাউন সিনড্রোম নিয়ে জন্মগ্রহণ করেন।জেনেটিক উপাদানগুলির অনেকগুলি ছোট "ভাঙ্গন" রয়েছে যা রোগের সূত্রপাতের দিকে পরিচালিত করে না, তবে, বিপরীতভাবে, ভাল বৈশিষ্ট্য দেয়। জেনেটিক উপাদানের সমস্ত "ভাঙ্গন" কে মিউটেশন বলা হয়। যে মিউটেশনগুলি রোগ বা জীবের বৈশিষ্ট্যগুলির অবনতির দিকে পরিচালিত করে তা নেতিবাচক বলে বিবেচিত হয় এবং নতুন উপকারী বৈশিষ্ট্যগুলির গঠনের দিকে পরিচালিত করে এমন মিউটেশনগুলিকে ইতিবাচক হিসাবে বিবেচনা করা হয়।
যাইহোক, বেশিরভাগ রোগের সাথে সম্পর্কযুক্ত যা মানুষ আজ ভোগে, এটি এমন একটি রোগ নয় যা উত্তরাধিকারসূত্রে পাওয়া যায়, তবে শুধুমাত্র একটি প্রবণতা। উদাহরণস্বরূপ, একটি সন্তানের পিতার মধ্যে, চিনি ধীরে ধীরে শোষিত হয়। এর অর্থ এই নয় যে শিশুটি ডায়াবেটিস নিয়ে জন্মগ্রহণ করবে, তবে শিশুর একটি প্রবণতা থাকবে। এর অর্থ হ'ল যদি কোনও শিশু মিষ্টি এবং আটার পণ্যের অপব্যবহার করে তবে তার ডায়াবেটিস হবে।
আজ তথাকথিত ভবিষ্যদ্বাণীমূলকঔষধ এই চিকিৎসা অনুশীলনের অংশ হিসাবে, একজন ব্যক্তির মধ্যে প্রবণতা সনাক্ত করা হয় (সংশ্লিষ্ট জিনগুলির সনাক্তকরণের উপর ভিত্তি করে), এবং তারপরে তাকে সুপারিশ করা হয় - কোন ডায়েট অনুসরণ করতে হবে, কীভাবে সঠিকভাবে কাজের শাসন এবং বিশ্রামের বিকল্প পরিবর্তন করতে হবে যাতে না হয়। অসুস্থ পেতে.
ডিএনএ-তে এনকোড করা তথ্য কীভাবে পড়তে হয়?
কিন্তু ডিএনএ-তে থাকা তথ্যগুলো কীভাবে পড়বেন? কিভাবে তার নিজের শরীর এটি ব্যবহার করে? ডিএনএ নিজেই এক ধরণের ম্যাট্রিক্স, তবে সহজ নয়, তবে এনকোডেড। ডিএনএ ম্যাট্রিক্স থেকে তথ্য পড়ার জন্য, এটি প্রথমে একটি বিশেষ ক্যারিয়ারে স্থানান্তরিত হয় - আরএনএ। RNA রাসায়নিকভাবে রাইবোনিউক্লিক অ্যাসিড। এটি ডিএনএ থেকে আলাদা যে এটি নিউক্লিয়ার ঝিল্লির মধ্য দিয়ে কোষে যেতে পারে, যখন ডিএনএ-তে এই ক্ষমতার অভাব রয়েছে (এটি শুধুমাত্র নিউক্লিয়াসে পাওয়া যায়)। এনকোড করা তথ্য ঘরেই ব্যবহৃত হয়। সুতরাং, আরএনএ হল নিউক্লিয়াস থেকে কোষে কোডেড তথ্যের বাহক।কীভাবে আরএনএ সংশ্লেষণ ঘটে, কীভাবে আরএনএর সাহায্যে প্রোটিন সংশ্লেষিত হয়?
 যে ডিএনএ স্ট্র্যান্ডগুলি থেকে তথ্য "পড়তে হবে" সেগুলি অবিকৃত, একটি বিশেষ এনজাইম, "নির্মাতা", তাদের কাছে যায় এবং ডিএনএ স্ট্র্যান্ডের সমান্তরালে একটি পরিপূরক আরএনএ চেইন সংশ্লেষ করে। আরএনএ অণুতে 4 ধরনের নিউক্লিওটাইড রয়েছে - অ্যাডেনিন (এ), ইউরাসিল (ইউ), গুয়ানিন (জি) এবং সাইটোসিন (সি)। এই ক্ষেত্রে, নিম্নলিখিত জোড়াগুলি পরিপূরক: অ্যাডেনিন - ইউরাসিল, গুয়ানিন - সাইটোসিন। আপনি দেখতে পাচ্ছেন, ডিএনএর বিপরীতে, আরএনএ থাইমিনের পরিবর্তে ইউরাসিল ব্যবহার করে। অর্থাৎ, "বিল্ডার" এনজাইমটি নিম্নরূপ কাজ করে: যদি এটি ডিএনএ স্ট্র্যান্ডে A দেখে, তবে এটি আরএনএ স্ট্র্যান্ডের সাথে Y সংযুক্ত করে, যদি G, তাহলে এটি C, ইত্যাদি সংযুক্ত করে। এইভাবে, প্রতিলিপির সময় প্রতিটি সক্রিয় জিন থেকে একটি টেমপ্লেট তৈরি হয় - আরএনএর একটি অনুলিপি যা পারমাণবিক ঝিল্লির মধ্য দিয়ে যেতে পারে।
যে ডিএনএ স্ট্র্যান্ডগুলি থেকে তথ্য "পড়তে হবে" সেগুলি অবিকৃত, একটি বিশেষ এনজাইম, "নির্মাতা", তাদের কাছে যায় এবং ডিএনএ স্ট্র্যান্ডের সমান্তরালে একটি পরিপূরক আরএনএ চেইন সংশ্লেষ করে। আরএনএ অণুতে 4 ধরনের নিউক্লিওটাইড রয়েছে - অ্যাডেনিন (এ), ইউরাসিল (ইউ), গুয়ানিন (জি) এবং সাইটোসিন (সি)। এই ক্ষেত্রে, নিম্নলিখিত জোড়াগুলি পরিপূরক: অ্যাডেনিন - ইউরাসিল, গুয়ানিন - সাইটোসিন। আপনি দেখতে পাচ্ছেন, ডিএনএর বিপরীতে, আরএনএ থাইমিনের পরিবর্তে ইউরাসিল ব্যবহার করে। অর্থাৎ, "বিল্ডার" এনজাইমটি নিম্নরূপ কাজ করে: যদি এটি ডিএনএ স্ট্র্যান্ডে A দেখে, তবে এটি আরএনএ স্ট্র্যান্ডের সাথে Y সংযুক্ত করে, যদি G, তাহলে এটি C, ইত্যাদি সংযুক্ত করে। এইভাবে, প্রতিলিপির সময় প্রতিটি সক্রিয় জিন থেকে একটি টেমপ্লেট তৈরি হয় - আরএনএর একটি অনুলিপি যা পারমাণবিক ঝিল্লির মধ্য দিয়ে যেতে পারে। কিভাবে একটি প্রোটিনের সংশ্লেষণ একটি নির্দিষ্ট জিন দ্বারা এনকোড করা হয়?
নিউক্লিয়াস ত্যাগ করার পর, আরএনএ সাইটোপ্লাজমে প্রবেশ করে। ইতিমধ্যে সাইটোপ্লাজমে, আরএনএ একটি ম্যাট্রিক্স হিসাবে হতে পারে, বিশেষ এনজাইম সিস্টেমে (রাইবোসোম) তৈরি, যা সংশ্লেষণ করতে পারে, আরএনএর তথ্য দ্বারা পরিচালিত, প্রোটিনের সংশ্লিষ্ট অ্যামিনো অ্যাসিড ক্রম। আপনি জানেন যে, একটি প্রোটিন অণু অ্যামিনো অ্যাসিড দিয়ে তৈরি। রাইবোসোম কীভাবে ক্রমবর্ধমান প্রোটিন শৃঙ্খলে কোন অ্যামিনো অ্যাসিড সংযুক্ত করবে তা জানতে পারে? এটি একটি ট্রিপলেট কোডের ভিত্তিতে করা হয়। ট্রিপলেট কোডের অর্থ হল RNA চেইনের তিনটি নিউক্লিওটাইডের ক্রম ( ত্রিপল,উদাহরণস্বরূপ, একটি অ্যামিনো অ্যাসিডের জন্য GGU) কোড (এই ক্ষেত্রে, গ্লাইসিন)। প্রতিটি অ্যামিনো অ্যাসিড একটি নির্দিষ্ট ট্রিপলেট দ্বারা এনকোড করা হয়। আর তাই, রাইবোসোম ত্রিপলকে "পড়ে", আরএনএ-তে তথ্য পড়ার সাথে সাথে পরবর্তীতে কোন অ্যামিনো অ্যাসিড যোগ করা উচিত তা নির্ধারণ করে। যখন অ্যামিনো অ্যাসিডের একটি শৃঙ্খল গঠিত হয়, তখন এটি একটি নির্দিষ্ট স্থানিক রূপ ধারণ করে এবং একটি প্রোটিনে পরিণত হয় যা এনজাইমেটিক, বিল্ডিং, হরমোন এবং এটির জন্য নির্ধারিত অন্যান্য কার্য সম্পাদন করতে সক্ষম।যে কোনো জীবের জন্য প্রোটিন একটি জিন পণ্য। এটি প্রোটিন যা জিনের সমস্ত বিভিন্ন বৈশিষ্ট্য, গুণাবলী এবং বাহ্যিক প্রকাশ নির্ধারণ করে।
ডানদিকে ভার্না (বুলগেরিয়া) সমুদ্র সৈকতে মানুষের দ্বারা নির্মিত বৃহত্তম মানব ডিএনএ হেলিক্স, যা 23 এপ্রিল, 2016-এ গিনেস বুক অফ রেকর্ডসে অন্তর্ভুক্ত হয়েছিল
ডিঅক্সিরাইবোনিউক্লিয়িক এসিড. সাধারণ জ্ঞাতব্য
ডিএনএ (ডিঅক্সিরাইবোনিউক্লিক অ্যাসিড) হল এক ধরনের জীবনের ব্লুপ্রিন্ট, একটি জটিল কোড যাতে বংশগত তথ্যের ডেটা থাকে। এই জটিল ম্যাক্রোমোলিকুল বংশগত জেনেটিক তথ্য প্রজন্ম থেকে প্রজন্মে সঞ্চয় ও প্রেরণ করতে সক্ষম। ডিএনএ বংশগত এবং পরিবর্তনশীলতার মতো যে কোনও জীবের বৈশিষ্ট্য নির্ধারণ করে। এতে এনকোড করা তথ্য যে কোনো জীবন্ত প্রাণীর সমগ্র উন্নয়ন কর্মসূচী নির্ধারণ করে। জেনেটিকালি এমবেডেড ফ্যাক্টরগুলি একজন ব্যক্তি এবং অন্য যেকোন জীব উভয়েরই জীবনের সমগ্র গতিপথ পূর্বনির্ধারিত করে। বাহ্যিক পরিবেশের কৃত্রিম বা প্রাকৃতিক প্রভাব পৃথক জেনেটিক বৈশিষ্ট্যগুলির সামগ্রিক তীব্রতাকে সামান্য প্রভাবিত করতে পারে বা প্রোগ্রাম করা প্রক্রিয়াগুলির বিকাশকে প্রভাবিত করতে পারে।
ডিঅক্সিরাইবোনিউক্লিয়িক এসিড(DNA) হল একটি ম্যাক্রোমোলিকিউল (তিনটি প্রধানের মধ্যে একটি, বাকি দুটি হল আরএনএ এবং প্রোটিন), যা সঞ্চয়, প্রজন্ম থেকে প্রজন্মে সংক্রমণ এবং জীবন্ত প্রাণীর বিকাশ ও কার্যকারিতার জন্য জেনেটিক প্রোগ্রাম বাস্তবায়ন প্রদান করে। ডিএনএ বিভিন্ন ধরনের আরএনএ এবং প্রোটিনের গঠন সম্পর্কে তথ্য ধারণ করে।
ইউক্যারিওটিক কোষে (প্রাণী, উদ্ভিদ এবং ছত্রাক), ডিএনএ কোষের নিউক্লিয়াসে ক্রোমোজোমের অংশ হিসেবে পাওয়া যায়, সেইসাথে কিছু কোষের অর্গানেলে (মাইটোকন্ড্রিয়া এবং প্লাস্টিড)। প্রোক্যারিওটিক জীবের কোষে (ব্যাকটেরিয়া এবং আর্কিয়া), একটি বৃত্তাকার বা রৈখিক ডিএনএ অণু, তথাকথিত নিউক্লিয়েড, কোষের ঝিল্লির ভেতর থেকে সংযুক্ত থাকে। তারা এবং নিম্ন ইউক্যারিওটস (উদাহরণস্বরূপ, খামির) এছাড়াও ছোট স্বায়ত্তশাসিত, বেশিরভাগ বৃত্তাকার ডিএনএ অণু রয়েছে যাকে প্লাজমিড বলা হয়।
রাসায়নিক দৃষ্টিকোণ থেকে, ডিএনএ একটি দীর্ঘ পলিমেরিক অণু যা পুনরাবৃত্তিমূলক ব্লকগুলি - নিউক্লিওটাইডগুলি নিয়ে গঠিত। প্রতিটি নিউক্লিওটাইড একটি নাইট্রোজেনাস বেস, একটি চিনি (ডিঅক্সিরিবোজ) এবং একটি ফসফেট গ্রুপ দিয়ে গঠিত। একটি শৃঙ্খলে নিউক্লিওটাইডগুলির মধ্যে বন্ধনগুলি ডিঅক্সিরিবোজ দ্বারা গঠিত হয় ( থেকে) এবং ফসফেট ( চ) গ্রুপ (ফসফোডিস্টার বন্ড)।

ভাত। 2. নিউক্লার্টাইড একটি নাইট্রোজেনাস বেস, চিনি (ডিঅক্সিরিবোজ) এবং একটি ফসফেট গ্রুপ নিয়ে গঠিত
অপ্রতিরোধ্য সংখ্যাগরিষ্ঠ ক্ষেত্রে (একক-স্ট্রেন্ডেড ডিএনএ ধারণকারী কিছু ভাইরাস ব্যতীত), ডিএনএ ম্যাক্রোমোলিকিউলে দুটি চেইন থাকে যা একে অপরের সাথে নাইট্রোজেনাস বেস দিয়ে থাকে। এই ডাবল-স্ট্র্যান্ডেড অণুটি হেলিক্সে পেঁচানো হয়।
ডিএনএতে চার ধরনের নাইট্রোজেনাস বেস পাওয়া যায় (অ্যাডেনাইন, গুয়ানিন, থাইমিন এবং সাইটোসিন)। একটি চেইনের নাইট্রোজেনাস বেসগুলি হাইড্রোজেন বন্ড দ্বারা অন্য চেইনের নাইট্রোজেনাস বেসগুলির সাথে সম্পূরকতার নীতি অনুসারে সংযুক্ত থাকে: অ্যাডেনিন শুধুমাত্র থাইমিনের সাথে একত্রিত হয় ( এ-টি), গুয়ানিন - শুধুমাত্র সাইটোসিনের সাথে ( জি-সি) এই জোড়াগুলিই ডিএনএর হেলিকাল "মই" এর "দন্ড" তৈরি করে (দেখুন: চিত্র 2, 3 এবং 4)।

ভাত। 2. নাইট্রোজেন ঘাঁটি
নিউক্লিওটাইডের ক্রম আপনাকে বিভিন্ন ধরণের RNA সম্পর্কে তথ্য "এনকোড" করতে দেয়, যার মধ্যে সবচেয়ে গুরুত্বপূর্ণ হল তথ্যগত বা টেমপ্লেট (mRNA), রাইবোসোমাল (rRNA) এবং পরিবহন (tRNA)। এই সমস্ত ধরণের আরএনএ ট্রান্সক্রিপশনের সময় সংশ্লেষিত আরএনএ সিকোয়েন্সে ডিএনএ ক্রম অনুলিপি করে ডিএনএ টেমপ্লেটে সংশ্লেষিত হয় এবং প্রোটিন জৈব সংশ্লেষণে (অনুবাদ প্রক্রিয়া) অংশ নেয়। কোডিং সিকোয়েন্স ছাড়াও, কোষের ডিএনএ-তে এমন ক্রম থাকে যা নিয়ন্ত্রক এবং কাঠামোগত কার্য সম্পাদন করে।

ভাত। 3. ডিএনএ প্রতিলিপি
ডিএনএ রাসায়নিক যৌগের মৌলিক সমন্বয়ের অবস্থান এবং এই সংমিশ্রণের মধ্যে পরিমাণগত অনুপাত বংশগত তথ্যের এনকোডিং প্রদান করে।
শিক্ষা নতুন ডিএনএ (প্রতিলিপি)
- প্রতিলিপির প্রক্রিয়া: ডিএনএ ডাবল হেলিক্সের মুক্ত হওয়া - ডিএনএ পলিমারেজ দ্বারা পরিপূরক স্ট্র্যান্ডের সংশ্লেষণ - একটি থেকে দুটি ডিএনএ অণুর গঠন।
- ডাবল হেলিক্স দুটি শাখায় "আনজিপ" করে যখন এনজাইম রাসায়নিক যৌগের ভিত্তি জোড়ার মধ্যে বন্ধন ভেঙ্গে দেয়।
- প্রতিটি শাখা একটি নতুন ডিএনএ উপাদান। নতুন বেস জোড়াগুলি মূল শাখার মতো একই ক্রমানুসারে সংযুক্ত থাকে।
ডুপ্লিকেশন সম্পন্ন হলে, দুটি স্বাধীন হেলিস গঠিত হয়, যা মূল ডিএনএর রাসায়নিক যৌগ থেকে তৈরি হয় এবং এর সাথে একই জেনেটিক কোড থাকে। এইভাবে, ডিএনএ কোষ থেকে কোষে তথ্যের মাধ্যমে ছিঁড়ে ফেলতে সক্ষম।
আরো বিস্তারিত তথ্য:
নিউক্লিক এসিডের গঠন

ভাত। চার নাইট্রোজেনাস বেস: অ্যাডেনিন, গুয়ানিন, সাইটোসিন, থাইমিন
ডিঅক্সিরাইবোনিউক্লিয়িক এসিড(ডিএনএ) নিউক্লিক অ্যাসিড বোঝায়। নিউক্লিক অ্যাসিডঅনিয়মিত বায়োপলিমারগুলির একটি শ্রেণি যার মনোমারগুলি নিউক্লিওটাইড।
নিউক্লিওটাইডসগঠিত নাইট্রোজেনাস বেস, একটি পাঁচ-কার্বন কার্বোহাইড্রেট (পেন্টোজ) এর সাথে সংযুক্ত - ডিঅক্সিরিবোজ(ডিএনএর ক্ষেত্রে) বা রাইবোস(RNA এর ক্ষেত্রে), যা একটি ফসফরিক অ্যাসিড অবশিষ্টাংশের সাথে একত্রিত হয় (H 2 PO 3 -)।
নাইট্রোজেন ঘাঁটিদুটি প্রকার রয়েছে: পাইরিমিডিন ঘাঁটি - ইউরাসিল (শুধুমাত্র আরএনএতে), সাইটোসিন এবং থাইমিন, পিউরিন বেস - অ্যাডেনিন এবং গুয়ানিন।

ভাত। চিত্র 5. নিউক্লিওটাইডের গঠন (বাম), ডিএনএ-তে নিউক্লিওটাইডের অবস্থান (নীচে) এবং নাইট্রোজেনাস ঘাঁটির ধরন (ডানে): পাইরিমিডিন এবং পিউরিন

একটি পেন্টোজ অণুর কার্বন পরমাণুর সংখ্যা 1 থেকে 5 পর্যন্ত। ফসফেট তৃতীয় এবং পঞ্চম কার্বন পরমাণুর সাথে মিলিত হয়। এইভাবে নিউক্লিক অ্যাসিডগুলি একসাথে যুক্ত হয়ে নিউক্লিক অ্যাসিডের একটি চেইন তৈরি করে। এইভাবে, আমরা ডিএনএ স্ট্র্যান্ডের 3' এবং 5' প্রান্তকে বিচ্ছিন্ন করতে পারি:

ভাত। 6. ডিএনএ স্ট্র্যান্ডের 3' এবং 5' প্রান্তের বিচ্ছিন্নতা
ডিএনএ গঠনের দুটি স্ট্র্যান্ড ডবল হেলিক্স. একটি সর্পিল এই চেইন বিপরীত দিকে ভিত্তিক হয়. ডিএনএর বিভিন্ন স্ট্র্যান্ডে, নাইট্রোজেনাস ঘাঁটিগুলি একে অপরের সাথে সংযুক্ত থাকে হাইড্রোজেন বন্ধন. অ্যাডেনিন সর্বদা থাইমিনের সাথে একত্রিত হয়, এবং সাইটোসিন সর্বদা গুয়ানিনের সাথে একত্রিত হয়। এটা কে বলে পরিপূরক নিয়ম(সেমি. পরিপূরকতার নীতি).
পরিপূরক নিয়ম:
| A-T G-C |
উদাহরণস্বরূপ, যদি আমাদের একটি ডিএনএ স্ট্র্যান্ড দেওয়া হয় যার ক্রম রয়েছে
3'-ATGTCCTAGCTGCTCG - 5',
তাহলে দ্বিতীয় চেইনটি এর পরিপূরক হবে এবং বিপরীত দিকে পরিচালিত হবে - 5'-এন্ড থেকে 3'-এন্ড পর্যন্ত:
5'- TACAGGATCGACGAGC- 3'।

ভাত। 7. ডিএনএ অণুর চেইনের দিক এবং হাইড্রোজেন বন্ড ব্যবহার করে নাইট্রোজেনাস বেসের সংযোগ
ডিএনএ রেপ্লিকেশন
ডিএনএ রেপ্লিকেশনটেমপ্লেট সংশ্লেষণের মাধ্যমে একটি ডিএনএ অণুকে দ্বিগুণ করার প্রক্রিয়া। প্রাকৃতিক ডিএনএ প্রতিলিপির বেশিরভাগ ক্ষেত্রেপ্রাইমারডিএনএ সংশ্লেষণের জন্য সংক্ষিপ্ত স্নিপেট (আবার তৈরি করা হয়েছে)। এই ধরনের একটি রাইবোনিউক্লিওটাইড প্রাইমার এনজাইম প্রাইমেস (প্রোক্যারিওটে ডিএনএ প্রাইমেজ, ইউক্যারিওটে ডিএনএ পলিমারেজ) দ্বারা তৈরি করা হয় এবং পরবর্তীতে ডিঅক্সিরাইবোনিউক্লিওটাইড পলিমারেজ দ্বারা প্রতিস্থাপিত হয়, যা সাধারণত মেরামতের কাজ করে (ডিএনএ অণুতে রাসায়নিক ক্ষতি এবং বিরতি সংশোধন)।
প্রতিলিপি একটি আধা-রক্ষণশীল পদ্ধতিতে ঘটে। এর মানে হল ডিএনএ-এর ডাবল হেলিক্স খুলে যায় এবং পরিপূরকতার নীতি অনুসারে এর প্রতিটি চেইনে একটি নতুন চেইন সম্পূর্ণ হয়। কন্যা ডিএনএ অণুতে এইভাবে প্যারেন্ট অণু থেকে একটি স্ট্র্যান্ড থাকে এবং একটি নতুন সংশ্লেষিত হয়। প্রতিলিপিটি প্যারেন্ট স্ট্র্যান্ডের 3' থেকে 5' দিকে ঘটে।

ভাত। 8. ডিএনএ অণুর প্রতিলিপি (দ্বিগুণ)
ডিএনএ সংশ্লেষণ- এটি এমন একটি জটিল প্রক্রিয়া নয় যতটা প্রথম নজরে মনে হতে পারে। আপনি যদি এটি সম্পর্কে চিন্তা করেন তবে প্রথমে আপনাকে সংশ্লেষণ কী তা খুঁজে বের করতে হবে। এটি কিছু একত্রিত করার প্রক্রিয়া। একটি নতুন ডিএনএ অণু গঠন বিভিন্ন পর্যায়ে সঞ্চালিত হয়:
1) ডিএনএ টোপোইসোমারেজ, প্রতিলিপি কাঁটাচামচের সামনে অবস্থিত, ডিএনএকে কেটে দেয় যাতে এটিকে মুক্ত করা এবং খোলার সুবিধা দেওয়া যায়।
2) ডিএনএ হেলিকেস, টপোইসোমারেজ অনুসরণ করে, ডিএনএ হেলিক্সকে "আনওয়াইন্ডিং" করার প্রক্রিয়াকে প্রভাবিত করে।
3) ডিএনএ-বাইন্ডিং প্রোটিনগুলি ডিএনএ স্ট্র্যান্ডগুলির বাঁধন সম্পাদন করে এবং তাদের স্থিতিশীলতাও সম্পাদন করে, তাদের একে অপরের সাথে আটকে থাকতে বাধা দেয়।
4) ডিএনএ পলিমারেজ δ(বদ্বীপ) , প্রতিলিপি কাঁটাচামচের গতির সাথে সমন্বিত, সংশ্লেষণ সম্পাদন করেনেতৃস্থানীয়চেইনসহায়ক ম্যাট্রিক্সে ডিএনএ 5" → 3" দিকেমাতৃ ডিএনএর স্ট্র্যান্ড এর 3" প্রান্ত থেকে 5" প্রান্ত পর্যন্ত (প্রতি সেকেন্ডে 100 বেস জোড়া পর্যন্ত গতি)। এ নিয়ে এসব ঘটনা মাতৃডিএনএর স্ট্র্যান্ড সীমিত।

ভাত। 9. ডিএনএ প্রতিলিপি প্রক্রিয়ার পরিকল্পিত উপস্থাপনা: (1) ল্যাগিং স্ট্র্যান্ড (ল্যাগ স্ট্র্যান্ড), (2) লিডিং স্ট্র্যান্ড (লিডিং স্ট্র্যান্ড), (3) ডিএনএ পলিমারেজ α (Polα), (4) ডিএনএ লিগেজ, (5) আরএনএ -প্রাইমার, (6) প্রাইমাস, (7) ওকাজাকি টুকরো, (8) ডিএনএ পলিমারেজ δ (পোলδ), (9) হেলিকেস, (10) একক-স্ট্র্যান্ডেড ডিএনএ-বাইন্ডিং প্রোটিন, (11) টপোইসোমারেজ।
ল্যাগিং কন্যা ডিএনএ স্ট্র্যান্ডের সংশ্লেষণ নীচে বর্ণিত হয়েছে (নীচে দেখুন)। পরিকল্পনাপ্রতিলিপি কাঁটা এবং প্রতিলিপি এনজাইমের কার্যকারিতা)
ডিএনএ প্রতিলিপি সম্পর্কে আরও তথ্যের জন্য, দেখুন
5) প্যারেন্ট অণুর অন্য স্ট্র্যান্ডের স্থিরকরণ এবং স্থিরকরণের পরপরই, এটি যোগ দেয়ডিএনএ পলিমারেজ α(আলফা)এবং দিক 5 "→3" একটি প্রাইমার (RNA প্রাইমার) সংশ্লেষিত করে - 10 থেকে 200 নিউক্লিওটাইডের দৈর্ঘ্য সহ একটি DNA টেমপ্লেটে একটি RNA ক্রম। এর পরে, এনজাইমডিএনএ স্ট্র্যান্ড থেকে সরানো হয়েছে।
পরিবর্তে ডিএনএ পলিমারেজα
প্রাইমারের 3" প্রান্তের সাথে সংযুক্তডিএনএ পলিমারেজε
.
6)
ডিএনএ পলিমারেজε
(এপিসিলন) যেন প্রাইমার লম্বা করতে থাকে, কিন্তু সাবস্ট্রেট এম্বেড করেডিঅক্সিরাইবোনিউক্লিওটাইডস(150-200 নিউক্লিওটাইডের পরিমাণে)। ফলস্বরূপ, দুটি অংশ থেকে একটি শক্ত সুতো তৈরি হয় -আরএনএ(যেমন প্রাইমার) এবং ডিএনএ.
ডিএনএ পলিমারেজ εএটি আগের প্রাইমারের মুখোমুখি না হওয়া পর্যন্ত কাজ করেটুকরো ওকাজাকি(একটু আগে সংশ্লেষিত)। এই এনজাইম তারপর চেইন থেকে সরানো হয়।
7) ডিএনএ পলিমারেজ β(বিটা) এর জায়গায় দাঁড়িয়েছেডিএনএ পলিমারেজ ε,একই দিকে সরে যায় (5" → 3") এবং তাদের জায়গায় ডিঅক্সিরাইবোনিউক্লিওটাইড ঢোকানোর সময় প্রাইমার রাইবোনিউক্লিওটাইডগুলি সরিয়ে দেয়। প্রাইমার সম্পূর্ণ অপসারণ না হওয়া পর্যন্ত এনজাইম কাজ করে, যেমন একটি ডিঅক্সিরাইবোনিউক্লিওটাইড পর্যন্ত (এমনকি আরও আগে সংশ্লেষিতডিএনএ পলিমারেজ ε). এনজাইম তার কাজের ফলাফল এবং সামনের ডিএনএ লিঙ্ক করতে সক্ষম হয় না, তাই এটি চেইন ছেড়ে যায়।
ফলস্বরূপ, কন্যা ডিএনএর একটি খণ্ড মা থ্রেডের ম্যাট্রিক্সে "মিথ্যা"। এটা কে বলেওকাজাকির টুকরো.
8) ডিএনএ লিগেস দুটি সংলগ্ন লিগেট টুকরো ওকাজাকি , অর্থাৎ 5 "-সেগমেন্টের শেষ, সংশ্লেষিতডিএনএ পলিমারেজ ε,এবং 3" চেইন এন্ড বিল্ট-ইনডিএনএ পলিমারেজβ .
RNA এর গঠন
রাইবোনিউক্লিক এসিড(আরএনএ) হল তিনটি প্রধান ম্যাক্রোমোলিকুলের একটি (অন্য দুটি হল ডিএনএ এবং প্রোটিন) যা সমস্ত জীবন্ত প্রাণীর কোষে পাওয়া যায়।
ঠিক ডিএনএর মতো, আরএনএ একটি দীর্ঘ শৃঙ্খল দ্বারা গঠিত যার প্রতিটি লিঙ্ককে বলা হয় নিউক্লিওটাইড. প্রতিটি নিউক্লিওটাইড একটি নাইট্রোজেনাস বেস, একটি রাইবোজ চিনি এবং একটি ফসফেট গ্রুপ দিয়ে গঠিত। যাইহোক, ডিএনএর বিপরীতে, আরএনএ সাধারণত দুটি স্ট্র্যান্ডের পরিবর্তে একটি থাকে। আরএনএ-তে পেন্টোজ রাইবোজ দ্বারা প্রতিনিধিত্ব করে, ডিঅক্সিরাইবোজ নয় (রাইবোজের দ্বিতীয় কার্বোহাইড্রেট পরমাণুর উপর একটি অতিরিক্ত হাইড্রক্সিল গ্রুপ রয়েছে)। অবশেষে, নাইট্রোজেনাস ঘাঁটিগুলির গঠনে ডিএনএ আরএনএ থেকে পৃথক: থাইমিনের পরিবর্তে ( টি) ইউরাসিল আরএনএতে উপস্থিত থাকে ( উ) , যা অ্যাডেনিনের পরিপূরক।
নিউক্লিওটাইডের ক্রম RNA কে জেনেটিক তথ্য এনকোড করতে দেয়। সমস্ত সেলুলার জীব প্রোটিন সংশ্লেষণ প্রোগ্রাম করতে RNA (mRNA) ব্যবহার করে।
সেলুলার আরএনএ একটি প্রক্রিয়ায় গঠিত হয় যাকে বলা হয় প্রতিলিপি , অর্থাৎ, একটি ডিএনএ টেমপ্লেটে আরএনএর সংশ্লেষণ, বিশেষ এনজাইম দ্বারা পরিচালিত - আরএনএ পলিমারেজ.
মেসেঞ্জার RNAs (mRNAs) তারপর নামক একটি প্রক্রিয়ায় অংশ নেয় সম্প্রচার সেগুলো. রাইবোসোমের অংশগ্রহণের সাথে mRNA টেমপ্লেটে প্রোটিন সংশ্লেষণ। অন্যান্য আরএনএ ট্রান্সক্রিপশনের পরে রাসায়নিক পরিবর্তনের মধ্য দিয়ে যায় এবং সেকেন্ডারি এবং টারশিয়ারি স্ট্রাকচার গঠনের পরে, তারা ফাংশনগুলি সম্পাদন করে যা আরএনএর ধরণের উপর নির্ভর করে।

ভাত। 10. নাইট্রোজেনাস বেস পরিপ্রেক্ষিতে ডিএনএ এবং আরএনএর মধ্যে পার্থক্য: থাইমিন (টি) এর পরিবর্তে, আরএনএতে ইউরাসিল (ইউ), যা অ্যাডেনিনের পরিপূরক।
ট্রান্সক্রিপশন
এটি একটি ডিএনএ টেমপ্লেটে আরএনএ সংশ্লেষণের প্রক্রিয়া। DNA সাইটের একটিতে unwinds. চেইনগুলির মধ্যে একটিতে এমন তথ্য রয়েছে যা RNA অণুতে অনুলিপি করা প্রয়োজন - এই চেইনটিকে কোডিং বলা হয়। ডিএনএর দ্বিতীয় স্ট্র্যান্ড, যা কোডিং স্ট্র্যান্ডের পরিপূরক, তাকে টেমপ্লেট স্ট্র্যান্ড বলা হয়। টেমপ্লেট চেইনে 3'-5' দিকে (ডিএনএ চেইন বরাবর) ট্রান্সক্রিপশনের প্রক্রিয়ায়, এটির পরিপূরক একটি আরএনএ চেইন সংশ্লেষিত হয়। এইভাবে, কোডিং স্ট্র্যান্ডের একটি RNA কপি তৈরি করা হয়।
![]()
ভাত। 11. প্রতিলিপির পরিকল্পিত উপস্থাপনা
উদাহরণস্বরূপ, যদি আমাদের কোডিং স্ট্র্যান্ডের ক্রম দেওয়া হয়
3'-ATGTCCTAGCTGCTCG - 5',
তারপর, পরিপূরকতার নিয়ম অনুসারে, ম্যাট্রিক্স চেইন ক্রমটি বহন করবে
5'- TACAGGATCGACGAGC- 3',
এবং এটি থেকে সংশ্লেষিত RNA হল ক্রম
ব্রডকাস্ট
মেকানিজম বিবেচনা করুন প্রোটিন সংশ্লেষণআরএনএ ম্যাট্রিক্সে, সেইসাথে জেনেটিক কোড এবং এর বৈশিষ্ট্য। এছাড়াও, স্পষ্টতার জন্য, নীচের লিঙ্কে, আমরা একটি জীবন্ত কোষে সংঘটিত প্রতিলিপি এবং অনুবাদের প্রক্রিয়াগুলি সম্পর্কে একটি ছোট ভিডিও দেখার পরামর্শ দিই:

ভাত। 12. প্রোটিন সংশ্লেষণের প্রক্রিয়া: আরএনএর জন্য ডিএনএ কোড, প্রোটিনের জন্য আরএনএ কোড
জিনগত সংকেত
জিনগত সংকেত- নিউক্লিওটাইডের ক্রম ব্যবহার করে প্রোটিনের অ্যামিনো অ্যাসিড ক্রম এনকোড করার একটি পদ্ধতি। প্রতিটি অ্যামিনো অ্যাসিড তিনটি নিউক্লিওটাইডের একটি ক্রম দ্বারা এনকোড করা হয় - একটি কোডন বা একটি ট্রিপলেট।
জেনেটিক কোড বেশিরভাগ প্রো- এবং ইউক্যারিওটে সাধারণ। টেবিলটি সমস্ত 64টি কোডন তালিকাভুক্ত করে এবং সংশ্লিষ্ট অ্যামিনো অ্যাসিড তালিকাভুক্ত করে। বেস অর্ডারটি mRNA এর 5" থেকে 3" শেষ পর্যন্ত।
সারণি 1. স্ট্যান্ডার্ড জেনেটিক কোড
|
১ম nie |
২য় বেস |
৩য় nie |
|||||||
|
উ |
গ |
ক |
জি |
||||||
|
উ |
উ উ উ |
(Phe/F) |
উ সি উ |
(Ser/S) |
উ আ উ |
(Tyr/Y) |
U G U |
(Cys/C) |
উ |
|
উ উ সি |
ইউ সি সি |
ইউ এ সি |
ইউ জি সি |
গ |
|||||
|
উ উ আ |
(লিউ/এল) |
তুমি ক |
U A A |
কোডন বন্ধ করুন** |
ইউ জি এ |
কোডন বন্ধ করুন** |
ক |
||
|
উ উ জি |
ইউ সি জি |
ইউ এ জি |
কোডন বন্ধ করুন** |
উ জি জি |
(Trp/W) |
জি |
|||
|
গ |
গ উ উ |
C C U |
(প্রো/পি) |
C A U |
(তার/এইচ) |
সি জি ইউ |
(আর্গ/আর) |
উ |
|
|
C U C |
গ গ গ |
C A C |
সি জি সি |
গ |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
সিজিএ |
ক |
||||
|
সি ইউ জি |
সি সি জি |
সি এ জি |
সি জি জি |
জি |
|||||
|
ক |
ক উ উ |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
এ জি ইউ |
(Ser/S) |
উ |
|
A U C |
ক সি গ |
A A C |
এ জি সি |
গ |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
এ জি এ |
ক |
||||
|
এ ইউ জি |
(মেট/এম) |
এ সি জি |
এ এ জি |
এ জি জি |
জি |
||||
|
জি |
G U U |
(ভাল/ভি) |
জি সি ইউ |
(আলা/আ) |
জি এ ইউ |
(Asp/D) |
G G U |
(গ্লাই/জি) |
উ |
|
জি ইউ সি |
জি সি সি |
জি এ সি |
জি জি সি |
গ |
|||||
|
জি ইউ এ |
জি সি এ |
জি এ এ |
(গ্লু/ই) |
জি জি এ |
ক |
||||
|
জি ইউ জি |
জি সি জি |
জি এ জি |
G G G |
জি |
|||||
ট্রিপলেটগুলির মধ্যে, 4টি বিশেষ ক্রম রয়েছে যা "বিরাম চিহ্ন" হিসাবে কাজ করে:
- * ট্রিপলেট AUG, এছাড়াও এনকোডিং methionine বলা হয় কোডন শুরু করুন. এই কোডন একটি প্রোটিন অণুর সংশ্লেষণ শুরু করে। সুতরাং, প্রোটিন সংশ্লেষণের সময়, অনুক্রমের প্রথম অ্যামিনো অ্যাসিড সর্বদা মেথিওনিন হবে।
- ** ট্রিপলেট UAA, UAGএবং UGAডাকা কোডন বন্ধ করুনএবং কোনো অ্যামিনো অ্যাসিডের জন্য কোড করবেন না। এই ক্রমগুলিতে, প্রোটিন সংশ্লেষণ বন্ধ হয়ে যায়।
জেনেটিক কোডের বৈশিষ্ট্য
1. ত্রিগুণ. প্রতিটি অ্যামিনো অ্যাসিড তিনটি নিউক্লিওটাইডের একটি ক্রম দ্বারা এনকোড করা হয় - একটি ট্রিপলেট বা কোডন।
2. ধারাবাহিকতা. ট্রিপলেটগুলির মধ্যে কোনও অতিরিক্ত নিউক্লিওটাইড নেই, তথ্য ক্রমাগত পড়া হয়।
3. অ-ওভারল্যাপিং. একটি নিউক্লিওটাইড একই সময়ে দুটি ট্রিপলেটের অংশ হতে পারে না।
4. অনন্যতা. একটি কোডন শুধুমাত্র একটি অ্যামিনো অ্যাসিডের জন্য কোড করতে পারে।

5. অধঃপতন. একটি অ্যামিনো অ্যাসিড বিভিন্ন কোডন দ্বারা এনকোড করা যেতে পারে।

6. বহুমুখিতা. জেনেটিক কোড সব জীবন্ত প্রাণীর জন্য একই।
উদাহরণ। আমাদের কোডিং স্ট্র্যান্ডের ক্রম দেওয়া হয়েছে:
3’- CCGATTGCACGTCGATCGTATA- 5’.
ম্যাট্রিক্স চেইনের ক্রম থাকবে:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
এখন আমরা এই চেইন থেকে তথ্যগত RNA "সংশ্লেষিত" করি:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
প্রোটিন সংশ্লেষণ 5' → 3' দিকে যায়, তাই জেনেটিক কোড "পড়তে" আমাদের ক্রমটি উল্টাতে হবে:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
এখন শুরু কোডন AUG খুঁজুন:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
ক্রমটিকে ত্রিপলে ভাগ করুন:
এইরকম শোনাচ্ছে: ডিএনএ থেকে তথ্য আরএনএ (ট্রান্সক্রিপশন), আরএনএ থেকে প্রোটিনে (অনুবাদ) স্থানান্তরিত হয়। ডিএনএ প্রতিলিপির মাধ্যমেও নকল করা যেতে পারে, এবং বিপরীত প্রতিলিপির প্রক্রিয়াও সম্ভব, যখন একটি আরএনএ টেমপ্লেট থেকে ডিএনএ সংশ্লেষিত হয়, তবে এই ধরনের প্রক্রিয়া মূলত ভাইরাসের বৈশিষ্ট্য।

ভাত। 13. কেন্দ্রীয় মতবাদ আণবিক জীববিজ্ঞান
জিনোম: জিন এবং ক্রোমোজোম
(সাধারণ ধারণা)
জিনোম - একটি জীবের সমস্ত জিনের সামগ্রিকতা; এর সম্পূর্ণ ক্রোমোজোম সেট।
জিনোম শব্দটি 1920 সালে জি. উইঙ্কলার দ্বারা প্রস্তাবিত হয়েছিল একই জৈবিক প্রজাতির জীবের ক্রোমোজোমের হ্যাপ্লয়েড সেটে থাকা জিনের সম্পূর্ণতা বর্ণনা করার জন্য। এই শব্দটির মূল অর্থ নির্দেশ করে যে জিনোমের ধারণা, জিনোটাইপের বিপরীতে, সমগ্র প্রজাতির একটি জেনেটিক বৈশিষ্ট্য, এবং কোনও ব্যক্তির নয়। আণবিক জেনেটিক্সের বিকাশের সাথে, এই শব্দটির অর্থ পরিবর্তিত হয়েছে। এটা জানা যায় যে ডিএনএ, যা বেশিরভাগ জীবের জিনগত তথ্যের বাহক এবং তাই, জিনোমের ভিত্তি তৈরি করে, শব্দের আধুনিক অর্থে শুধুমাত্র জিনই অন্তর্ভুক্ত করে না। অধিকাংশইউক্যারিওটিক কোষের ডিএনএ নন-কোডিং ("অপ্রয়োজনীয়") নিউক্লিওটাইড ক্রম দ্বারা প্রতিনিধিত্ব করা হয় যাতে প্রোটিন এবং নিউক্লিক অ্যাসিড সম্পর্কে তথ্য থাকে না। সুতরাং, যেকোন জীবের জিনোমের প্রধান অংশ হল তার ক্রোমোজোমের হ্যাপ্লয়েড সেটের সম্পূর্ণ ডিএনএ।
জিন হল ডিএনএ অণুর সেগমেন্ট যা পলিপেপটাইড এবং আরএনএ অণুর জন্য কোড করে।
গত শতাব্দীতে, জিন সম্পর্কে আমাদের বোঝার উল্লেখযোগ্যভাবে পরিবর্তিত হয়েছে। পূর্বে, একটি জিনোম ছিল একটি ক্রোমোজোমের একটি অঞ্চল যা একটি বৈশিষ্ট্যকে এনকোড বা নির্ধারণ করে বা ফেনোটাইপিক(দৃশ্যমান) সম্পত্তি, যেমন চোখের রঙ।
1940 সালে, জর্জ বিডল এবং এডওয়ার্ড টাথাম একটি জিনের একটি আণবিক সংজ্ঞা প্রস্তাব করেছিলেন। বিজ্ঞানীরা ছত্রাকের বীজ প্রক্রিয়াকরণ করেছেন নিউরোস্পোরা ক্র্যাসাএক্স-রে এবং অন্যান্য এজেন্ট যা ডিএনএ সিকোয়েন্সে পরিবর্তন ঘটায় ( মিউটেশন), এবং ছত্রাকের মিউট্যান্ট স্ট্রেন পাওয়া গেছে যা কিছু নির্দিষ্ট এনজাইম হারিয়েছে, যা কিছু ক্ষেত্রে পুরো বিপাকীয় পথের ব্যাঘাত ঘটায়। Beadle এবং Tatham এই সিদ্ধান্তে উপনীত হন যে একটি জিন হল জেনেটিক উপাদানের একটি অংশ যা একটি একক এনজাইমের জন্য সংজ্ঞায়িত বা কোড করে। এভাবেই অনুমান "একটি জিন, একটি এনজাইম". এই ধারণাটি পরে সংজ্ঞা পর্যন্ত প্রসারিত হয় "একটি জিন - একটি পলিপেপটাইড", যেহেতু অনেক জিন প্রোটিনকে এনকোড করে যা এনজাইম নয়, এবং একটি পলিপেপটাইড একটি জটিল প্রোটিন কমপ্লেক্সের একটি সাবুনিট হতে পারে।
ডুমুর উপর. 14 ডিএনএ ট্রিপলেটগুলি কীভাবে একটি পলিপেপটাইড নির্ধারণ করে, একটি প্রোটিনের অ্যামিনো অ্যাসিড ক্রম, এমআরএনএ দ্বারা মধ্যস্থতা করে তার একটি চিত্র দেখায়। ডিএনএ স্ট্র্যান্ডগুলির মধ্যে একটি এমআরএনএ সংশ্লেষণের জন্য একটি টেমপ্লেটের ভূমিকা পালন করে, যার মধ্যে নিউক্লিওটাইড ট্রিপলেট (কোডন) ডিএনএ ট্রিপলেটের পরিপূরক। কিছু ব্যাকটেরিয়া এবং অনেক ইউক্যারিওটে, কোডিং ক্রমগুলি নন-কোডিং অঞ্চল দ্বারা বাধাপ্রাপ্ত হয় (যাকে বলা হয় introns).
একটি জিনের আধুনিক জৈব রাসায়নিক সংজ্ঞা এমনকি আরো নির্দিষ্টভাবে। জিন হল ডিএনএ-র সমস্ত বিভাগ যা শেষ পণ্যগুলির প্রাথমিক ক্রম এনকোড করে, যার মধ্যে পলিপেপটাইড বা আরএনএ রয়েছে যার গঠনগত বা অনুঘটক ফাংশন রয়েছে।
জিনের পাশাপাশি, ডিএনএ-তে অন্যান্য ক্রমও রয়েছে যা একচেটিয়াভাবে নিয়ন্ত্রক কার্য সম্পাদন করে। নিয়ন্ত্রক ক্রমজিনের শুরু বা শেষ চিহ্নিত করতে পারে, ট্রান্সক্রিপশনকে প্রভাবিত করতে পারে বা প্রতিলিপি বা পুনঃসংযোগের সূচনার স্থান নির্দেশ করতে পারে। কিছু জিন বিভিন্ন উপায়ে প্রকাশ করা যেতে পারে, একই টুকরো ডিএনএ বিভিন্ন পণ্য গঠনের জন্য একটি টেমপ্লেট হিসাবে পরিবেশন করে।
আমরা মোটামুটি হিসাব করতে পারি ন্যূনতম জিনের আকারমধ্যবর্তী প্রোটিনের জন্য কোডিং। পলিপেপটাইড চেইনের প্রতিটি অ্যামিনো অ্যাসিড তিনটি নিউক্লিওটাইডের ক্রম দ্বারা এনকোড করা হয়; এই ট্রিপলেটগুলির (কোডন) ক্রমগুলি প্রদত্ত জিন দ্বারা এনকোড করা পলিপেপটাইডের অ্যামিনো অ্যাসিডের চেইনের সাথে মিলে যায়। 350 অ্যামিনো অ্যাসিডের অবশিষ্টাংশের একটি পলিপেপটাইড চেইন (মাঝারি দৈর্ঘ্যের চেইন) 1050 bp এর ক্রম অনুসারে। ( bp) যাইহোক, অনেক ইউক্যারিওটিক জিন এবং কিছু প্রোক্যারিওটিক জিন ডিএনএ সেগমেন্ট দ্বারা বাধাগ্রস্ত হয় যা করে না তথ্যের বাহকপ্রোটিন সম্পর্কে, এবং সেইজন্য একটি সাধারণ গণনা শো থেকে অনেক দীর্ঘ হতে চালু আউট.
একটি ক্রোমোজোমে কয়টি জিন থাকে?
 ভাত। 15. প্রোক্যারিওটিক (বাম) এবং ইউক্যারিওটিক কোষে ক্রোমোজোমের দৃশ্য। হিস্টোন হল পারমাণবিক প্রোটিনের একটি বিস্তৃত শ্রেণী যা দুটি প্রধান কাজ সম্পাদন করে: তারা নিউক্লিয়াসে ডিএনএ স্ট্র্যান্ডের প্যাকেজিং এবং ট্রান্সক্রিপশন, প্রতিলিপিকরণ এবং মেরামতের মতো পারমাণবিক প্রক্রিয়াগুলির এপিজেনেটিক নিয়ন্ত্রণে জড়িত।
ভাত। 15. প্রোক্যারিওটিক (বাম) এবং ইউক্যারিওটিক কোষে ক্রোমোজোমের দৃশ্য। হিস্টোন হল পারমাণবিক প্রোটিনের একটি বিস্তৃত শ্রেণী যা দুটি প্রধান কাজ সম্পাদন করে: তারা নিউক্লিয়াসে ডিএনএ স্ট্র্যান্ডের প্যাকেজিং এবং ট্রান্সক্রিপশন, প্রতিলিপিকরণ এবং মেরামতের মতো পারমাণবিক প্রক্রিয়াগুলির এপিজেনেটিক নিয়ন্ত্রণে জড়িত।
প্রোক্যারিওটগুলির ডিএনএ আরও সহজ: তাদের কোষগুলির একটি নিউক্লিয়াস নেই, তাই ডিএনএ একটি নিউক্লিওড আকারে সরাসরি সাইটোপ্লাজমে অবস্থিত।
 আপনি জানেন যে, ব্যাকটেরিয়া কোষগুলির একটি ডিএনএ স্ট্র্যান্ডের আকারে একটি ক্রোমোজোম থাকে, যা একটি কম্প্যাক্ট কাঠামোতে প্যাক করা হয় - একটি নিউক্লিয়েড। প্রোক্যারিওটিক ক্রোমোজোম Escherichia coli, যার জিনোম সম্পূর্ণরূপে ডিকোড করা হয়, এটি একটি বৃত্তাকার ডিএনএ অণু (আসলে, এটি নয় ডান বৃত্ত, বরং শুরু বা শেষ ছাড়া একটি লুপ), যার মধ্যে 4,639,675 b.p. এই ক্রমটিতে প্রায় 4300টি প্রোটিন জিন এবং স্থিতিশীল আরএনএ অণুর জন্য আরও 157টি জিন রয়েছে। AT মানুষের জিনোমআনুমানিক 3.1 বিলিয়ন বেস জোড়া 24টি ভিন্ন ক্রোমোসোমে অবস্থিত প্রায় 29,000 জিনের সাথে সম্পর্কিত।
আপনি জানেন যে, ব্যাকটেরিয়া কোষগুলির একটি ডিএনএ স্ট্র্যান্ডের আকারে একটি ক্রোমোজোম থাকে, যা একটি কম্প্যাক্ট কাঠামোতে প্যাক করা হয় - একটি নিউক্লিয়েড। প্রোক্যারিওটিক ক্রোমোজোম Escherichia coli, যার জিনোম সম্পূর্ণরূপে ডিকোড করা হয়, এটি একটি বৃত্তাকার ডিএনএ অণু (আসলে, এটি নয় ডান বৃত্ত, বরং শুরু বা শেষ ছাড়া একটি লুপ), যার মধ্যে 4,639,675 b.p. এই ক্রমটিতে প্রায় 4300টি প্রোটিন জিন এবং স্থিতিশীল আরএনএ অণুর জন্য আরও 157টি জিন রয়েছে। AT মানুষের জিনোমআনুমানিক 3.1 বিলিয়ন বেস জোড়া 24টি ভিন্ন ক্রোমোসোমে অবস্থিত প্রায় 29,000 জিনের সাথে সম্পর্কিত।
প্রোক্যারিওটস (ব্যাকটেরিয়া)।
 ব্যাকটেরিয়াম ই কোলাইএকটি ডাবল-স্ট্র্যান্ডেড বৃত্তাকার ডিএনএ অণু রয়েছে। এটি 4,639,675 b.p নিয়ে গঠিত। এবং প্রায় 1.7 মিমি দৈর্ঘ্যে পৌঁছায়, যা কোষের দৈর্ঘ্যকে অতিক্রম করে ই কোলাইপ্রায় 850 বার। নিউক্লিয়েডের অংশ হিসাবে বৃহৎ বৃত্তাকার ক্রোমোজোম ছাড়াও, অনেক ব্যাকটেরিয়ায় এক বা একাধিক ছোট বৃত্তাকার ডিএনএ অণু থাকে যা অবাধে সাইটোসোলে অবস্থিত। এই extrachromosom উপাদান বলা হয় প্লাজমিড(চিত্র 16)।
ব্যাকটেরিয়াম ই কোলাইএকটি ডাবল-স্ট্র্যান্ডেড বৃত্তাকার ডিএনএ অণু রয়েছে। এটি 4,639,675 b.p নিয়ে গঠিত। এবং প্রায় 1.7 মিমি দৈর্ঘ্যে পৌঁছায়, যা কোষের দৈর্ঘ্যকে অতিক্রম করে ই কোলাইপ্রায় 850 বার। নিউক্লিয়েডের অংশ হিসাবে বৃহৎ বৃত্তাকার ক্রোমোজোম ছাড়াও, অনেক ব্যাকটেরিয়ায় এক বা একাধিক ছোট বৃত্তাকার ডিএনএ অণু থাকে যা অবাধে সাইটোসোলে অবস্থিত। এই extrachromosom উপাদান বলা হয় প্লাজমিড(চিত্র 16)।
বেশিরভাগ প্লাজমিড মাত্র কয়েক হাজার বেস জোড়া নিয়ে গঠিত, কিছুতে 10,000 bp এর বেশি থাকে। তারা জেনেটিক তথ্য বহন করে এবং কন্যা প্লাজমিড গঠনের জন্য প্রতিলিপি তৈরি করে, যা পিতামাতার কোষের বিভাজনের সময় কন্যা কোষে প্রবেশ করে। প্লাজমিডগুলি কেবল ব্যাকটেরিয়াতেই নয়, খামির এবং অন্যান্য ছত্রাকেও পাওয়া যায়। অনেক ক্ষেত্রে, প্লাজমিড হোস্ট কোষের জন্য কোন সুবিধা দেয় না এবং তাদের একমাত্র কাজ হল স্বাধীনভাবে পুনরুৎপাদন করা। যাইহোক, কিছু প্লাজমিড হোস্টের জন্য দরকারী জিন বহন করে। উদাহরণস্বরূপ, প্লাজমিডে থাকা জিনগুলি ব্যাকটেরিয়া কোষে অ্যান্টিব্যাকটেরিয়াল এজেন্টদের প্রতিরোধ করতে পারে। β-ল্যাকটামেস জিন বহনকারী প্লাজমিডগুলি পেনিসিলিন এবং অ্যামোক্সিসিলিনের মতো β-ল্যাকটাম অ্যান্টিবায়োটিকের প্রতিরোধ করে। প্লাজমিড অ্যান্টিবায়োটিক-প্রতিরোধী কোষ থেকে একই বা ভিন্ন ব্যাকটেরিয়া প্রজাতির অন্যান্য কোষে যেতে পারে, যার ফলে সেই কোষগুলিও প্রতিরোধী হয়ে ওঠে। অ্যান্টিবায়োটিকের নিবিড় ব্যবহার একটি শক্তিশালী নির্বাচনী ফ্যাক্টর যা প্যাথোজেনিক ব্যাকটেরিয়াগুলির মধ্যে অ্যান্টিবায়োটিক প্রতিরোধের এনকোডিং প্লাজমিডের বিস্তারকে উৎসাহিত করে (পাশাপাশি ট্রান্সপোসন যা অনুরূপ জিনগুলিকে এনকোড করে) এবং বিভিন্ন অ্যান্টিবায়োটিকের প্রতিরোধের সাথে ব্যাকটেরিয়াল স্ট্রেনের উত্থানের দিকে পরিচালিত করে। চিকিত্সকরা অ্যান্টিবায়োটিকের ব্যাপক ব্যবহারের বিপদগুলি বুঝতে শুরু করেছেন এবং যখনই প্রয়োজন তখনই এগুলি লিখে দেন। অনুরূপ কারণে, খামারের পশুদের চিকিত্সার জন্য অ্যান্টিবায়োটিকের ব্যাপক ব্যবহার সীমিত।
আরো দেখুন: রাভিন N.V., Shestakov S.V. প্রোক্যারিওটসের জিনোম // ভ্যাভিলভ জার্নাল অফ জেনেটিক্স অ্যান্ড ব্রিডিং, 2013. ভি. 17. নং 4/2। পৃষ্ঠা 972-984।
ইউক্যারিওটস।
সারণী 2. কিছু জীবের ডিএনএ, জিন এবং ক্রোমোজোম
|
ভাগ করা ডিএনএ, b.s |
ক্রোমোজোমের সংখ্যা* |
জিনের আনুমানিক সংখ্যা |
|
|
Escherichia coli(ব্যাকটেরিয়া) |
4 639 675 |
4 435 |
|
|
স্যাকারোমাইসিস সেরাভিসি(খামির) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(নেমাটোড) |
90 269 800 |
12*** |
23 000 |
|
আরবিডোপসিস থালিয়ানা(উদ্ভিদ) |
119 186 200 |
33 000 |
|
|
ড্রোসোফিলা মেলানোজেস্টার(ফলের মাছি) |
120 367 260 |
20 000 |
|
|
ওরিজা স্যাটিভা(ভাত) |
480 000 000 |
57 000 |
|
|
Mus পেশী(মাউস) |
2 634 266 500 |
27 000 |
|
|
হোমো সেপিয়েন্স(মানুষ) |
3 070 128 600 |
29 000 |
বিঃদ্রঃ.তথ্য ক্রমাগত আপডেট করা হয়; আরও আপ-টু-ডেট তথ্যের জন্য, পৃথক জিনোমিক প্রকল্পের ওয়েবসাইটগুলি পড়ুন।
* সমস্ত ইউক্যারিওটের জন্য, খামির ছাড়া, ক্রোমোজোমের ডিপ্লয়েড সেট দেওয়া হয়। ডিপ্লয়েডকিট ক্রোমোজোম (গ্রীক ডিপ্লোস থেকে - ডবল এবং ইডোস - ভিউ) - ক্রোমোজোমের ডবল সেট(2n), যার প্রত্যেকটিরই নিজস্ব একটি সমতা রয়েছে।
** হ্যাপ্লয়েড সেট। খামিরের বন্য স্ট্রেইনে সাধারণত এই ক্রোমোজোমের আটটি (অক্টাপ্লয়েড) বা তার বেশি সেট থাকে।
*** দুটি X ক্রোমোজোম সহ মহিলাদের জন্য। পুরুষদের একটি X ক্রোমোজোম আছে, কিন্তু Y নেই, অর্থাৎ শুধুমাত্র 11টি ক্রোমোজোম।
একটি খামির কোষ, ক্ষুদ্রতম ইউক্যারিওটগুলির মধ্যে একটি, একটি কোষের চেয়ে 2.6 গুণ বেশি ডিএনএ রয়েছে ই কোলাই(টেবিল ২). ফলের মাছি কোষ ড্রোসোফিলা, জেনেটিক গবেষণার একটি ক্লাসিক বস্তু, 35 গুণ বেশি ডিএনএ ধারণ করে এবং মানুষের কোষে কোষের তুলনায় প্রায় 700 গুণ বেশি ডিএনএ থাকে ই কোলাই.অনেক গাছপালা এবং উভচরের মধ্যে আরও বেশি ডিএনএ থাকে। ইউক্যারিওটিক কোষের জেনেটিক উপাদান ক্রোমোজোম আকারে সংগঠিত হয়। ক্রোমোজোমের ডিপ্লয়েড সেট (2 n) জীবের ধরণের উপর নির্ভর করে (সারণী 2)।
উদাহরণস্বরূপ, একটি মানুষের সোম্যাটিক কোষে 46টি ক্রোমোজোম থাকে ( চাল 17) একটি ইউক্যারিওটিক কোষের প্রতিটি ক্রোমোজোম, যেমন চিত্রে দেখানো হয়েছে। 17, ক, একটি খুব বড় ডবল-স্ট্র্যান্ডেড ডিএনএ অণু রয়েছে। চব্বিশটি মানব ক্রোমোজোম (22 জোড়া ক্রোমোজোম এবং দুটি সেক্স ক্রোমোজোম X এবং Y) 25 গুণেরও বেশি দৈর্ঘ্যে পৃথক। প্রতিটি ইউক্যারিওটিক ক্রোমোজোমে জিনের একটি নির্দিষ্ট সেট থাকে।

ভাত। 17। ইউক্যারিওটিক ক্রোমোজোম।ক- মানব ক্রোমোজোম থেকে সংযুক্ত এবং ঘনীভূত বোন ক্রোমাটিডের একটি জোড়া। এই ফর্মে, ইউক্যারিওটিক ক্রোমোজোমগুলি প্রতিলিপির পরে এবং মাইটোসিসের সময় মেটাফেজে থাকে। খ- বইয়ের লেখকদের একজনের লিউকোসাইট থেকে ক্রোমোজোমের একটি সম্পূর্ণ সেট। প্রতিটি স্বাভাবিক মানুষের সোম্যাটিক কোষে 46টি ক্রোমোজোম থাকে।

বংশগত উপাদান সংরক্ষণ এবং প্রেরণের জন্য একটি ম্যাট্রিক্স হিসাবে ডিএনএর আকার এবং কার্যকারিতা এই অণুর সংগঠনে বিশেষ কাঠামোগত উপাদানগুলির উপস্থিতি ব্যাখ্যা করে। উচ্চতর জীবগুলিতে, ডিএনএ ক্রোমোজোমের মধ্যে বিতরণ করা হয়।
একটি জীবের ডিএনএ (ক্রোমোজোম) এর সেটকে জিনোম বলা হয়। ক্রোমোজোম কোষের নিউক্লিয়াসে অবস্থিত এবং ক্রোমাটিন নামে একটি গঠন গঠন করে। ক্রোমাটিন হল 1:1 অনুপাতে ডিএনএ এবং মৌলিক প্রোটিন (হিস্টোন) এর একটি জটিল। ডিএনএর দৈর্ঘ্য সাধারণত পরিপূরক নিউক্লিওটাইডের জোড়ার সংখ্যা (বিপি) দ্বারা পরিমাপ করা হয়। উদাহরণস্বরূপ, 3য় মানব ক্রোমোজোমশতাব্দী হল একটি ডিএনএ অণু যার আকার 160 মিলিয়ন bp। এর দৈর্ঘ্য প্রায় 1 মিমি, তাই, তৃতীয় মানব ক্রোমোজোমের একটি রৈখিক অণুর দৈর্ঘ্য হবে 5 মিমি, এবং একটি হ্যাপ্লয়েডের সমস্ত 23টি ক্রোমোজোমের ডিএনএ (~ 3 * 10 9 bp, MR = 1.8 * 10 12) কোষ - ডিম বা শুক্রাণু কোষ - একটি রৈখিক আকারে 1 মিটার হবে। জীবাণু কোষ ব্যতীত, মানবদেহের সমস্ত কোষে (এগুলির মধ্যে প্রায় 1013টি রয়েছে) ক্রোমোজোমের একটি দ্বিগুণ সেট ধারণ করে। কোষ বিভাজনের সময়, সমস্ত 46টি ডিএনএ অণু প্রতিলিপি করে এবং 46টি ক্রোমোজোমে পুনর্গঠিত হয়।
আপনি যদি মানব জিনোমের ডিএনএ অণুগুলিকে (22 ক্রোমোজোম এবং ক্রোমোজোম X এবং Y বা X এবং X) একে অপরের সাথে সংযুক্ত করেন তবে আপনি প্রায় এক মিটার দীর্ঘ একটি ক্রম পাবেন। দ্রষ্টব্য: সমস্ত স্তন্যপায়ী এবং অন্যান্য হেটেরোগ্যামেটিক পুরুষ জীবের মধ্যে, মহিলাদের দুটি X ক্রোমোজোম (XX) এবং পুরুষদের একটি X ক্রোমোজোম এবং একটি Y ক্রোমোজোম (XY) থাকে।
বেশিরভাগ মানব কোষ, তাই এই ধরনের কোষের মোট ডিএনএ দৈর্ঘ্য প্রায় 2 মি। একজন প্রাপ্তবয়স্ক মানুষের প্রায় 10 14 কোষ থাকে, তাই সমস্ত DNA অণুর মোট দৈর্ঘ্য 2・10 11 কিমি। তুলনা করার জন্য, পৃথিবীর পরিধি হল 4・10 4 কিমি, এবং পৃথিবী থেকে সূর্যের দূরত্ব হল 1.5・10 8 কিমি। আমাদের কোষে কতটা আশ্চর্যজনকভাবে কম্প্যাক্টলি প্যাকেজ করা ডিএনএ!
ইউক্যারিওটিক কোষে, ডিএনএ ধারণকারী অন্যান্য অর্গানেল রয়েছে - এগুলি হল মাইটোকন্ড্রিয়া এবং ক্লোরোপ্লাস্ট। মাইটোকন্ড্রিয়াল এবং ক্লোরোপ্লাস্ট ডিএনএ এর উৎপত্তি সম্পর্কে অনেক অনুমান সামনে রাখা হয়েছে। আজ সাধারণভাবে গৃহীত দৃষ্টিভঙ্গি হল যে তারা প্রাচীন ব্যাকটেরিয়ার ক্রোমোজোমের মূল উপাদান যা হোস্ট কোষের সাইটোপ্লাজমে প্রবেশ করেছিল এবং এই অর্গানেলগুলির অগ্রদূত হয়ে উঠেছে। মাইটোকন্ড্রিয়াল টিআরএনএ এবং আরআরএনএর জন্য মাইটোকন্ড্রিয়াল ডিএনএ কোড, পাশাপাশি বেশ কয়েকটি মাইটোকন্ড্রিয়াল প্রোটিন। 95% এর বেশি মাইটোকন্ড্রিয়াল প্রোটিন পারমাণবিক ডিএনএ দ্বারা এনকোড করা হয়।
জিনের গঠন
প্রোক্যারিওটস এবং ইউক্যারিওটে জিনের গঠন, তাদের মিল এবং পার্থক্য বিবেচনা করুন। একটি জিন হল ডিএনএ-এর একটি অংশ যা শুধুমাত্র একটি প্রোটিন বা আরএনএ এনকোডিং করে, সরাসরি কোডিং অংশ ছাড়াও, এতে নিয়ন্ত্রক এবং অন্যান্য কাঠামোগত উপাদান রয়েছে যেগুলির প্রোক্যারিওটস এবং ইউক্যারিওটে আলাদা কাঠামো রয়েছে।
কোডিং ক্রম- জিনের প্রধান কাঠামোগত এবং কার্যকরী একক, এটিতে নিউক্লিওটাইডের ট্রিপলেট এনকোডিংঅ্যামিনো অ্যাসিড ক্রম। এটি একটি স্টার্ট কোডন দিয়ে শুরু হয় এবং একটি স্টপ কোডন দিয়ে শেষ হয়।
আগে এবং পরে কোডিং সিকোয়েন্স হয় অনূদিত 5' এবং 3' ক্রম. তারা নিয়ন্ত্রক এবং সহায়ক ফাংশন সম্পাদন করে, উদাহরণস্বরূপ, এমআরএনএ-তে রাইবোসোমের অবতরণ নিশ্চিত করে।
অঅনুবাদিত এবং কোডিং ক্রমগুলি ট্রান্সক্রিপশনের একক তৈরি করে - প্রতিলিপিকৃত ডিএনএ অঞ্চল, অর্থাৎ, ডিএনএ অঞ্চল যেখান থেকে এমআরএনএ সংশ্লেষিত হয়।
টার্মিনেটরএকটি জিনের শেষে ডিএনএ-এর একটি নন-লিপিকৃত অঞ্চল যেখানে আরএনএ সংশ্লেষণ বন্ধ হয়ে যায়।
শুরুতে জিন থাকে নিয়ন্ত্রক এলাকা, যা রয়েছে প্রচারকএবং অপারেটর.
প্রচারক- ট্রান্সক্রিপশন দীক্ষার সময় যে ক্রমটির সাথে পলিমারেজ আবদ্ধ হয়। অপারেটর- এটি সেই ক্ষেত্র যেখানে বিশেষ প্রোটিন আবদ্ধ হতে পারে - দমনকারী, যা এই জিন থেকে আরএনএ সংশ্লেষণের কার্যকলাপ কমাতে পারে - অন্য কথায়, এটি কমাতে পারে অভিব্যক্তি.
প্রোক্যারিওটে জিনের গঠন
প্রোক্যারিওটস এবং ইউক্যারিওটে জিনের গঠনের জন্য সাধারণ পরিকল্পনা আলাদা নয় - উভয়টিতে একটি প্রবর্তক এবং অপারেটর সহ একটি নিয়ন্ত্রক অঞ্চল, কোডিং এবং অ-অনুবাদিত সিকোয়েন্স সহ একটি ট্রান্সক্রিপশন ইউনিট এবং একটি টার্মিনেটর রয়েছে। যাইহোক, প্রোক্যারিওট এবং ইউক্যারিওটে জিনের সংগঠন আলাদা।

ভাত। 18. প্রোক্যারিওটে (ব্যাকটেরিয়া) জিনের গঠনের স্কিম -ইমেজ বড় করা হয়
অপারোনের শুরুতে এবং শেষে, বেশ কয়েকটি কাঠামোগত জিনের জন্য সাধারণ নিয়ন্ত্রক অঞ্চল রয়েছে। অপেরনের প্রতিলিপিকৃত অঞ্চল থেকে, একটি mRNA অণু পঠিত হয়, এতে বেশ কয়েকটি কোডিং সিকোয়েন্স রয়েছে, যার প্রতিটির নিজস্ব স্টার্ট এবং স্টপ কোডন রয়েছে। এই প্রতিটি এলাকা থেকেএকটি প্রোটিন সংশ্লেষিত হয়। এইভাবে, একটি আই-আরএনএ অণু থেকে বেশ কয়েকটি প্রোটিন অণু সংশ্লেষিত হয়।
প্রোক্যারিওটগুলি একটি একক কার্যকরী ইউনিটে একাধিক জিনের সংমিশ্রণ দ্বারা চিহ্নিত করা হয় - অপেরন. অপেরনের কাজটি অন্যান্য জিন দ্বারা নিয়ন্ত্রিত হতে পারে, যা লক্ষণীয়ভাবে অপেরন থেকেই সরানো যেতে পারে - নিয়ন্ত্রক. এই জিন থেকে অনুবাদ করা প্রোটিন বলা হয় দমনকারী. এটি অপারোনের অপারেটরের সাথে আবদ্ধ হয়, এটিতে থাকা সমস্ত জিনের অভিব্যক্তিকে একযোগে নিয়ন্ত্রণ করে।
Prokaryotes এছাড়াও ঘটনা দ্বারা চিহ্নিত করা হয় ট্রান্সক্রিপশন এবং অনুবাদ কনজুগেশন.
![]()
ভাত। 19 প্রোক্যারিওটে প্রতিলিপি এবং অনুবাদের সংমিশ্রণের ঘটনা - ইমেজ বড় করা হয়
সাইটোপ্লাজমকে আলাদা করে এমন একটি পারমাণবিক ঝিল্লির উপস্থিতির কারণে ইউক্যারিওটে এই জোড়াটি ঘটে না, যেখানে অনুবাদ ঘটে, জেনেটিক উপাদান থেকে, যার উপর ট্রান্সক্রিপশন ঘটে। প্রোক্যারিওটে, ডিএনএ টেমপ্লেটে আরএনএ সংশ্লেষণের সময়, একটি রাইবোসোম অবিলম্বে সংশ্লেষিত আরএনএ অণুর সাথে আবদ্ধ হতে পারে। এইভাবে, ট্রান্সক্রিপশন সম্পূর্ণ হওয়ার আগেই অনুবাদ শুরু হয়। তদুপরি, বেশ কয়েকটি রাইবোসোম একই সাথে একটি আরএনএ অণুর সাথে আবদ্ধ হতে পারে, একযোগে একটি প্রোটিনের বেশ কয়েকটি অণু সংশ্লেষিত করে।
ইউক্যারিওটে জিনের গঠন
ইউক্যারিওটের জিন এবং ক্রোমোজোম খুব জটিলভাবে সংগঠিত।
অনেক প্রজাতির ব্যাকটেরিয়ায় একটি মাত্র ক্রোমোজোম থাকে এবং প্রায় সব ক্ষেত্রেই প্রতিটি ক্রোমোজোমে প্রতিটি জিনের একটি করে অনুলিপি থাকে। শুধুমাত্র কয়েকটি জিন, যেমন rRNA জিন, একাধিক কপিতে থাকে। জিন এবং নিয়ন্ত্রক ক্রম প্রোক্যারিওটসের প্রায় পুরো জিনোম তৈরি করে। অধিকন্তু, প্রায় প্রতিটি জিনই এনকোড করা অ্যামিনো অ্যাসিড সিকোয়েন্স (বা RNA ক্রম) এর সাথে কঠোরভাবে মিলে যায় (চিত্র 14)।
ইউক্যারিওটিক জিনের গঠনগত ও কার্যকরী সংগঠন অনেক বেশি জটিল। ইউক্যারিওটিক ক্রোমোজোমের অধ্যয়ন, এবং পরে সম্পূর্ণ ইউক্যারিওটিক জিনোম সিকোয়েন্সের সিকোয়েন্সিং অনেক বিস্ময় নিয়ে এসেছে। অনেকের, বেশির ভাগ না হলেও, ইউক্যারিওটিক জিনের একটি আকর্ষণীয় বৈশিষ্ট্য রয়েছে: তাদের নিউক্লিওটাইড ক্রমগুলিতে এক বা একাধিক ডিএনএ অঞ্চল থাকে যা পলিপেপটাইড পণ্যের অ্যামিনো অ্যাসিড ক্রমকে এনকোড করে না। এই ধরনের অ-অনুবাদিত সন্নিবেশগুলি জিনের নিউক্লিওটাইড ক্রম এবং এনকোডেড পলিপেপটাইডের অ্যামিনো অ্যাসিড ক্রমগুলির মধ্যে সরাসরি সঙ্গতিকে ব্যাহত করে। জিনের এই অ-অনুবাদিত অংশগুলোকে বলা হয় introns, বা অন্তর্নির্মিত ক্রম, এবং কোডিং সেগমেন্ট হয় exons. প্রোক্যারিওটে, শুধুমাত্র কয়েকটি জিনে অন্তর্নিহিত থাকে।
সুতরাং, ইউক্যারিওটে, অপারনগুলিতে জিনগুলির কার্যত কোন সংমিশ্রণ নেই এবং ইউক্যারিওটিক জিনের কোডিং ক্রমটি প্রায়শই অনুবাদিত অঞ্চলে বিভক্ত হয়। - exons, এবং অনূদিত বিভাগ - introns
বেশিরভাগ ক্ষেত্রে, ইন্ট্রোনের ফাংশন প্রতিষ্ঠিত হয়নি। সাধারণভাবে, মানুষের ডিএনএর মাত্র 1.5% "কোডিং" হয়, অর্থাৎ এটি প্রোটিন বা আরএনএ সম্পর্কে তথ্য বহন করে। যাইহোক, বড় অন্তঃকরণগুলিকে বিবেচনায় নিয়ে, এটি দেখা যাচ্ছে যে মানুষের ডিএনএর 30% জিন নিয়ে গঠিত। যেহেতু জিনগুলি মানব জিনোমের একটি অপেক্ষাকৃত ছোট অনুপাত তৈরি করে, তাই ডিএনএর একটি উল্লেখযোগ্য পরিমাণ হিসাবহীন থেকে যায়।

ভাত। 16. ইউক্যারিওটে জিনের গঠনের স্কিম - ইমেজ বড় করা হয়
প্রতিটি জিন থেকে, একটি অপরিণত, বা প্রাক-আরএনএ, প্রথমে সংশ্লেষিত হয়, যার মধ্যে ইন্ট্রোন এবং এক্সন উভয়ই থাকে।
এর পরে, স্প্লিসিং প্রক্রিয়াটি সঞ্চালিত হয়, যার ফলস্বরূপ ইন্ট্রন অঞ্চলগুলি কেটে ফেলা হয় এবং একটি পরিপক্ক এমআরএনএ গঠিত হয়, যা থেকে একটি প্রোটিন সংশ্লেষিত হতে পারে।

ভাত। 20. বিকল্প স্প্লিসিং প্রক্রিয়া - ইমেজ বড় করা হয়
জিনের এই ধরনের সংগঠন অনুমতি দেয়, উদাহরণস্বরূপ, যখন একটি জিন থেকে প্রোটিনের বিভিন্ন রূপকে সংশ্লেষিত করা যেতে পারে, এই কারণে যে বিভক্ত করার সময় এক্সনগুলি বিভিন্ন ক্রম অনুসারে মিশ্রিত হতে পারে।

ভাত। 21. প্রোক্যারিওটস এবং ইউক্যারিওটসের জিনের গঠনে পার্থক্য - ইমেজ বড় করা হয়
মিউটেশন এবং মিউটেজেনেসিস
মিউটেশনযাকে বলা হয় জিনোটাইপের অবিরাম পরিবর্তন, অর্থাৎ নিউক্লিওটাইড অনুক্রমের পরিবর্তন।
যে প্রক্রিয়াটি মিউটেশনের দিকে নিয়ে যায় তাকে বলা হয় mutagenesis, এবং জীব সবযার কোষ একই মিউটেশন বহন করে মিউট্যান্ট.
মিউটেশন তত্ত্বপ্রথম 1903 সালে Hugh de Vries দ্বারা প্রণয়ন করা হয়েছিল। এর আধুনিক সংস্করণে নিম্নলিখিত বিধানগুলি অন্তর্ভুক্ত রয়েছে:
1. মিউটেশন ঘটে হঠাৎ করে, আকস্মিকভাবে।
2. মিউটেশন প্রজন্ম থেকে প্রজন্মে চলে যায়।
3. মিউটেশন উপকারী, ক্ষতিকারক বা নিরপেক্ষ, প্রভাবশালী বা পশ্চাদপসরণকারী হতে পারে।
4. মিউটেশন সনাক্ত করার সম্ভাবনা অধ্যয়ন করা ব্যক্তির সংখ্যার উপর নির্ভর করে।
5. অনুরূপ মিউটেশন বারবার ঘটতে পারে।
6. মিউটেশন নির্দেশিত হয় না.
বিভিন্ন কারণের প্রভাবে মিউটেশন ঘটতে পারে। দ্বারা সৃষ্ট মিউটেশনের মধ্যে পার্থক্য করুন mutagenic প্রভাব: শারীরিক (যেমন অতিবেগুনি বা বিকিরণ), রাসায়নিক (যেমন কোলচিসিন বা প্রতিক্রিয়াশীল অক্সিজেন প্রজাতি) এবং জৈবিক (যেমন ভাইরাস)। মিউটেশনও হতে পারে প্রতিলিপি ত্রুটি.
মিউটেশন চেহারা জন্য শর্ত উপর নির্ভর করে বিভক্ত করা হয় স্বতঃস্ফূর্ত- অর্থাৎ, মিউটেশন যা স্বাভাবিক অবস্থায় উদ্ভূত হয়েছে, এবং প্ররোচিত- অর্থাৎ, বিশেষ অবস্থার অধীনে উদ্ভূত মিউটেশন।
মিউটেশনগুলি কেবল পারমাণবিক ডিএনএতেই নয়, উদাহরণস্বরূপ, মাইটোকন্ড্রিয়া বা প্লাস্টিডের ডিএনএতেও ঘটতে পারে। তদনুসারে, আমরা পার্থক্য করতে পারি পারমাণবিকএবং সাইটোপ্লাজমিকমিউটেশন
মিউটেশনের সংঘটনের ফলস্বরূপ, নতুন অ্যালিলগুলি প্রায়শই উপস্থিত হতে পারে। যদি মিউট্যান্ট অ্যালিল স্বাভাবিক অ্যালিলকে ওভাররাইড করে তবে মিউটেশন বলা হয় প্রভাবশালী. যদি স্বাভাবিক অ্যালিল পরিবর্তিতকে দমন করে, তবে তাকে মিউটেশন বলা হয় পতনশীল. নতুন অ্যালিলের জন্ম দেয় এমন বেশিরভাগ মিউটেশন রিসেসিভ।
মিউটেশনগুলি প্রভাব দ্বারা আলাদা করা হয় অভিযোজিত, পরিবেশের সাথে জীবের অভিযোজন ক্ষমতা বৃদ্ধির দিকে পরিচালিত করে, নিরপেক্ষযা বেঁচে থাকাকে প্রভাবিত করে না ক্ষতিকরযা পরিবেশগত অবস্থার সাথে জীবের অভিযোজনযোগ্যতা হ্রাস করে এবং প্রাণঘাতীবিকাশের প্রাথমিক পর্যায়ে জীবের মৃত্যুর দিকে নিয়ে যায়।
পরিণতি অনুসারে, মিউটেশনগুলিকে আলাদা করা হয়, যা নেতৃত্ব দেয় প্রোটিন ফাংশন ক্ষতি, মিউটেশনের দিকে পরিচালিত করে উত্থান প্রোটিন একটি নতুন ফাংশন আছে, সেইসাথে মিউটেশন যে একটি জিনের ডোজ পরিবর্তন করুন, এবং, সেই অনুযায়ী, এটি থেকে সংশ্লেষিত প্রোটিনের ডোজ।
শরীরের যে কোনো কোষে মিউটেশন ঘটতে পারে। জীবাণু কোষে মিউটেশন ঘটলে তাকে বলা হয় জীবাণুসংক্রান্ত(জীবাণু, বা উৎপন্ন)। এই ধরনের মিউটেশনগুলি জীবের মধ্যে উপস্থিত হয় না যেখানে তারা আবির্ভূত হয়, তবে বংশধরদের মধ্যে মিউট্যান্টদের উপস্থিতির দিকে পরিচালিত করে এবং উত্তরাধিকারসূত্রে প্রাপ্ত হয়, তাই তারা জেনেটিক্স এবং বিবর্তনের জন্য গুরুত্বপূর্ণ। অন্য কোন কোষে মিউটেশন ঘটলে তাকে বলা হয় সোমাটিক. এই ধরনের মিউটেশন কিছু পরিমাণে জীবের মধ্যে নিজেকে প্রকাশ করতে পারে যেখানে এটি উদ্ভূত হয়, উদাহরণস্বরূপ, ক্যান্সারের টিউমার গঠনের দিকে পরিচালিত করে। যাইহোক, এই ধরনের মিউটেশন উত্তরাধিকারসূত্রে পাওয়া যায় না এবং বংশকে প্রভাবিত করে না।
মিউটেশন বিভিন্ন আকারের জিনোমের অংশগুলিকে প্রভাবিত করতে পারে। বরাদ্দ জেনেটিক, ক্রোমোসোমালএবং জিনোমিকমিউটেশন
জিন মিউটেশন
একটি জিনের চেয়ে ছোট স্কেলে যে মিউটেশনগুলি ঘটে তাকে বলা হয় জেনেটিক, বা ডটেড (ডটেড). এই ধরনের মিউটেশন ক্রমানুসারে এক বা একাধিক নিউক্লিওটাইডের পরিবর্তন ঘটায়। জিন মিউটেশন অন্তর্ভুক্তপ্রতিস্থাপন, একটি নিউক্লিওটাইড অন্য দ্বারা প্রতিস্থাপনের দিকে পরিচালিত করে,মুছে ফেলানিউক্লিওটাইডগুলির একটির ক্ষতির দিকে পরিচালিত করে,সন্নিবেশ, ক্রমটিতে একটি অতিরিক্ত নিউক্লিওটাইড যোগ করে।

ভাত। 23. জিন (বিন্দু) মিউটেশন
প্রোটিনের উপর ক্রিয়া করার পদ্ধতি অনুসারে, জিন মিউটেশনগুলিকে ভাগ করা হয়:সমার্থক, যা (জেনেটিক কোডের অবক্ষয়ের ফলে) প্রোটিন পণ্যের অ্যামিনো অ্যাসিড গঠনে পরিবর্তন আনে না,ভুল মিউটেশন, যা একটি অ্যামিনো অ্যাসিড অন্য দ্বারা প্রতিস্থাপনের দিকে পরিচালিত করে এবং সংশ্লেষিত প্রোটিনের গঠনকে প্রভাবিত করতে পারে, যদিও প্রায়শই তারা নগণ্য,আজেবাজে মিউটেশন, একটি স্টপ কোডন দিয়ে কোডিং কোডন প্রতিস্থাপনের দিকে পরিচালিত করে,মিউটেশনের দিকে পরিচালিত করে স্প্লিসিং ব্যাধি:

ভাত। 24. মিউটেশন স্কিম
এছাড়াও, প্রোটিনের উপর ক্রিয়া করার পদ্ধতি অনুসারে, মিউটেশনগুলি বিচ্ছিন্ন হয়, যার ফলে ফ্রেম শিফট পড়াযেমন সন্নিবেশ এবং মুছে ফেলা। এই ধরনের মিউটেশনগুলি, অর্থহীন মিউটেশনের মতো, যদিও সেগুলি জিনের এক পর্যায়ে ঘটে, প্রায়শই প্রোটিনের সম্পূর্ণ কাঠামোকে প্রভাবিত করে, যা হতে পারে সম্পূর্ণ পরিবর্তনএর কাঠামো। যখন একটি ক্রোমোজোমের একটি অংশ 180 ডিগ্রি ঘোরে ভাত। 28. স্থানান্তর
যখন একটি ক্রোমোজোমের একটি অংশ 180 ডিগ্রি ঘোরে ভাত। 28. স্থানান্তর
ভাত। 29. নকলের আগে এবং পরে ক্রোমোজোম
জিনোমিক মিউটেশন
অবশেষে, জিনোমিক মিউটেশনপুরো জিনোমকে প্রভাবিত করে, অর্থাৎ ক্রোমোজোমের সংখ্যা পরিবর্তন করে। পলিপ্লয়েডি আলাদা করা হয় - কোষের চালচলন বৃদ্ধি, এবং অ্যানিউপ্লয়েডি, অর্থাৎ, ক্রোমোজোমের সংখ্যার পরিবর্তন, উদাহরণস্বরূপ, ট্রাইসোমি (ক্রোমোজোমের একটিতে একটি অতিরিক্ত হোমোলগের উপস্থিতি) এবং মনোসোমি (এর অনুপস্থিতি) ক্রোমোজোমে একটি হোমোলজ)।
ডিএনএ সম্পর্কিত ভিডিও
ডিএনএ প্রতিলিপি, আরএনএ কোডিং, প্রোটিন সংশ্লেষণ
(যদি ভিডিওটি প্রদর্শিত না হয় তবে এটি উপলব্ধ
ডিএনএর রাসায়নিক গঠন অনুসারে ( ডিঅক্সিরাইবোনিউক্লিয়িক এসিড) হয় বায়োপলিমার, যার monomers হয় নিউক্লিওটাইড. অর্থাৎ ডিএনএ হলো পলিনিউক্লিওটাইড. অধিকন্তু, একটি ডিএনএ অণু সাধারণত একটি হেলিকাল রেখা বরাবর একে অপরের সাথে পেঁচানো দুটি চেইন নিয়ে গঠিত (যাকে প্রায়ই "সর্পিল পাকানো" বলা হয়) এবং হাইড্রোজেন বন্ড দ্বারা আন্তঃসংযুক্ত।
চেইনগুলি বাম এবং ডানদিকে (বেশিরভাগ সময়) উভয় দিকেই পাকানো যেতে পারে।
কিছু ভাইরাসের একক স্ট্র্যান্ড ডিএনএ থাকে।
প্রতিটি ডিএনএ নিউক্লিওটাইডে থাকে ১) নাইট্রোজেনাস বেস, ২) ডিঅক্সিরাইবোজ, ৩) ফসফরিক অ্যাসিডের অবশিষ্টাংশ।
ডাবল ডান হাতের ডিএনএ হেলিক্স
ডিএনএতে নিম্নলিখিতগুলি রয়েছে: adenine, গুয়ানিন, থাইমিনএবং সাইটোসিন. এডেনাইন ও গুয়ানিন হয় পিউরিন, এবং থাইমিন এবং সাইটোসিন - থেকে pyrimidines. কখনও কখনও ডিএনএতে ইউরাসিল থাকে, যা সাধারণত আরএনএর বৈশিষ্ট্য, যেখানে এটি থাইমিনকে প্রতিস্থাপন করে।
একটি ডিএনএ অণুর একটি শৃঙ্খলের নাইট্রোজেনাস ঘাঁটিগুলি অন্যটির নাইট্রোজেনাস ঘাঁটির সাথে কঠোরভাবে সম্পূরকতার নীতি অনুসারে সংযুক্ত থাকে: অ্যাডেনিন শুধুমাত্র থাইমিনের সাথে (তারা নিজেদের মধ্যে দুটি হাইড্রোজেন বন্ধন তৈরি করে), এবং গুয়ানিন শুধুমাত্র সাইটোসিনের সাথে (তিনটি বন্ধন)।
নিউক্লিওটাইডে নাইট্রোজেনাস বেস নিজেই সাইক্লিক ফর্মের প্রথম কার্বন পরমাণুর সাথে সংযুক্ত থাকে ডিঅক্সিরিবোজ, যা একটি পেন্টোজ (পাঁচটি কার্বন পরমাণু সহ কার্বোহাইড্রেট)। বন্ধনটি সমযোজী, গ্লাইকোসিডিক (C-N)। রাইবোসের বিপরীতে, ডিঅক্সিরাইবোজে এর একটি হাইড্রক্সিল গ্রুপের অভাব রয়েছে। ডিঅক্সিরাইবোজের বলয়টি চারটি কার্বন পরমাণু এবং একটি অক্সিজেন পরমাণু দ্বারা গঠিত হয়। পঞ্চম কার্বন পরমাণুটি বলয়ের বাইরে থাকে এবং একটি অক্সিজেন পরমাণুর মাধ্যমে একটি ফসফরিক অ্যাসিড অবশিষ্টাংশের সাথে সংযুক্ত থাকে। এছাড়াও, তৃতীয় কার্বন পরমাণুতে অক্সিজেন পরমাণুর মাধ্যমে, প্রতিবেশী নিউক্লিওটাইডের ফসফরিক অ্যাসিড অবশিষ্টাংশ সংযুক্ত থাকে।
এইভাবে, ডিএনএর একটি স্ট্র্যান্ডে, সংলগ্ন নিউক্লিওটাইডগুলি পরস্পর সংযুক্ত থাকে সমযোজী বন্ধনেরডিঅক্সিরিবোজ এবং ফসফরিক অ্যাসিড (ফসফোডিস্টার বন্ড) এর মধ্যে। একটি ফসফেট-ডিঅক্সিরিবোজ ব্যাকবোন গঠিত হয়। এটির লম্বভাবে, ডিএনএর অন্য স্ট্র্যান্ডের দিকে, নাইট্রোজেনাস ঘাঁটিগুলি নির্দেশিত হয়, যা হাইড্রোজেন বন্ড দ্বারা দ্বিতীয় স্ট্র্যান্ডের ঘাঁটির সাথে সংযুক্ত থাকে।
ডিএনএ-র গঠন এমন যে হাইড্রোজেন বন্ড দ্বারা সংযুক্ত চেইনের মেরুদণ্ড বিভিন্ন দিকে পরিচালিত হয় (তারা বলে "মাল্টিডাইরেশনাল", "অ্যান্টিপ্যারালাল")। যে দিকে একটি ফসফরিক অ্যাসিড দিয়ে শেষ হয় ডিঅক্সিরিবোজের পঞ্চম কার্বন পরমাণুর সাথে যুক্ত, অন্যটি "মুক্ত" তৃতীয় কার্বন পরমাণুর সাথে শেষ হয়। অর্থাৎ, একটি চেইনের কঙ্কাল অন্যটির সাথে আপেক্ষিক হিসাবে উল্টে গেছে। এইভাবে, ডিএনএ চেইনের গঠনে, 5 "প্রান্ত এবং 3" প্রান্ত আলাদা করা হয়।
ডিএনএ-র প্রতিলিপি (দ্বিগুণ) করার সময়, নতুন চেইনগুলির সংশ্লেষণ সর্বদা তাদের 5 তম প্রান্ত থেকে তৃতীয় প্রান্তে অগ্রসর হয়, যেহেতু নতুন নিউক্লিওটাইডগুলি কেবলমাত্র বিনামূল্যে তৃতীয় প্রান্তে সংযুক্ত হতে পারে।
শেষ পর্যন্ত (পরোক্ষভাবে আরএনএর মাধ্যমে), প্রতিটি পরপর তিনটি নিউক্লিওটাইড ডিএনএ চেইন কোডে প্রোটিনের একটি অ্যামিনো অ্যাসিডের জন্য।
ডিএনএ অণুর গঠন আবিষ্কার 1953 সালে এফ. ক্রিক এবং ডি. ওয়াটসনের কাজের (যা অন্যান্য বিজ্ঞানীদের প্রাথমিক কাজ দ্বারা সহজতর হয়েছিল) এর কাজের জন্য ঘটেছিল। যদিও কিভাবে রাসায়নিক পদার্থডিএনএ 19 শতক থেকে পরিচিত। 1940 এর দশকে, এটি স্পষ্ট হয়ে ওঠে যে ডিএনএ জেনেটিক তথ্যের বাহক।
ডাবল হেলিক্সকে ডিএনএ অণুর গৌণ গঠন হিসাবে বিবেচনা করা হয়। ইউক্যারিওটিক কোষে, ডিএনএর সিংহভাগই ক্রোমোজোমে অবস্থিত, যেখানে এটি প্রোটিন এবং অন্যান্য পদার্থের সাথে যুক্ত এবং ঘন প্যাকেজিংয়ের মধ্য দিয়ে যায়।
ডিঅক্সিরাইবোনিউক্লিয়িক এসিড বা ডিএনএ জেনেটিক তথ্যের বাহক। কোষের বেশিরভাগ ডিএনএ নিউক্লিয়াসে অবস্থিত। এটি ক্রোমোজোমের প্রধান উপাদান। ইউক্যারিওটে, ডিএনএ মাইটোকন্ড্রিয়া এবং প্লাস্টিডেও পাওয়া যায়। ডিএনএ একটি দীর্ঘ শাখাবিহীন পলিমারের প্রতিনিধিত্ব করে একে অপরের সাথে সমযোজীভাবে সংযুক্ত মনোনিউক্লিওটাইড নিয়ে গঠিত। ডিএনএ তৈরি করা মনোনিউক্লিওটাইডগুলি ডিঅক্সিরাইবোজ নিয়ে গঠিত, 4টি নাইট্রোজেনাস ঘাঁটির মধ্যে একটি (অ্যাডেনাইন, গুয়ানিন, সাইটোসিন এবং থাইমিন), এবং একটি ফসফরিক অ্যাসিড অবশিষ্টাংশ। এই মনোনিউক্লিওটাইডের সংখ্যা অনেক বেশি। উদাহরণস্বরূপ, একটি একক ক্রোমোজোম ধারণকারী প্রোক্যারিওটিক কোষে, ডিএনএ হল একটি একক ম্যাক্রোমোলিকুল যার আণবিক ওজন 2 x 10 9 এর বেশি।
ডিএনএর এক স্ট্র্যান্ডের মনোনিউক্লিওটাইডগুলি গঠনের কারণে একে অপরের সাথে সিরিজে সংযুক্ত থাকে সমযোজী ফসফোডিস্টার বন্ড একটি মনোনিউক্লিওটাইডের ডিঅক্সিরাইবোজ ওএইচ গ্রুপ এবং অন্যটির ফসফরিক অ্যাসিড অবশিষ্টাংশের মধ্যে। ডিএনএর এক স্ট্র্যান্ডের গঠিত মেরুদণ্ডের একপাশে নাইট্রোজেনাস বেস রয়েছে। তারা একটি থ্রেড উপর করা চারটি ভিন্ন জপমালা সঙ্গে তুলনা করা যেতে পারে, কারণ. তারা, যেমন ছিল, একটি চিনি-ফসফেট শৃঙ্খলে আটকে আছে।
প্রশ্ন জাগে, কিভাবে এই দীর্ঘ পলিনিউক্লিওটাইড চেইন একটি কোষ বা এমনকি একটি সম্পূর্ণ জীবের বিকাশের জন্য প্রোগ্রামকে এনকোড করতে পারে? ডিএনএর স্থানিক গঠন কীভাবে গঠিত হয় তা বোঝার মাধ্যমে এই প্রশ্নের উত্তর পাওয়া যেতে পারে। এই অণুর গঠনটি 1953 সালে জে. ওয়াটসন এবং এফ. ক্রিক দ্বারা পাঠোদ্ধার এবং বর্ণনা করা হয়েছিল।
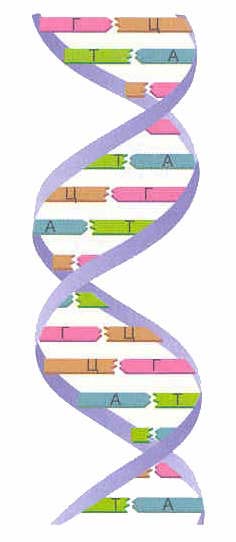
ডিএনএ অণু দুটি স্ট্র্যান্ড যা একে অপরের সমান্তরাল এবং গঠন করে ডান হাতের হেলিক্স
. এই সর্পিলটির প্রস্থ প্রায় 2 এনএম, তবে এর দৈর্ঘ্য কয়েক হাজার ন্যানোমিটারে পৌঁছাতে পারে। ওয়াটসন এবং ক্রিক ডিএনএ-র একটি মডেল প্রস্তাব করেছিলেন, যে অনুসারে সমস্ত ডিএনএ বেস হেলিক্সের ভিতরে অবস্থিত, চিনি-ফসফেট ব্যাকবোন বাইরে। সুতরাং, একটি চেইনের ঘাঁটিগুলি অন্যটির ঘাঁটির সাথে যতটা সম্ভব কাছাকাছি,
তাই তাদের মধ্যে হাইড্রোজেন বন্ধন তৈরি হয়। ডিএনএ হেলিক্সের গঠনটি এমন যে এটি তৈরি করা পলিনিউক্লিওটাইড চেইনগুলিকে ক্ষতবিক্ষত করার পরেই আলাদা করা যায়।
ডিএনএর দুটি স্ট্র্যান্ডের সর্বাধিক নৈকট্যের কারণে, এর রচনায় এক ধরণের (অ্যাডেনাইন এবং গুয়ানিন) একই পরিমাণ নাইট্রোজেনাস ঘাঁটি এবং অন্য ধরণের (থাইমিন এবং সাইটোসিন) নাইট্রোজেনাস ঘাঁটি রয়েছে, অর্থাৎ, সূত্রটি বৈধ: A+G=T+C. এটি নাইট্রোজেনাস ঘাঁটির আকারের কারণে, যথা, অ্যাডেনিন-থাইমিন এবং গুয়ানাইন-সাইটোসিন জোড়ার মধ্যে হাইড্রোজেন বন্ধনের কারণে যে কাঠামো তৈরি হয় তার দৈর্ঘ্য প্রায় 1.1 এনএম। এই জোড়ার মোট মাত্রা ডিএনএ হেলিক্সের ভেতরের অংশের মাত্রার সাথে মিলে যায়। একটি সর্পিল গঠন দম্পতি C-Tখুব ছোট হবে দম্পতি এ-জি, বিপরীতভাবে, খুব বড়. অর্থাৎ, ডিএনএর প্রথম স্ট্র্যান্ডের নাইট্রোজেনাস বেস, সেই বেসটিকে নির্দিষ্ট করে যা ডিএনএর অন্য স্ট্র্যান্ডের একই জায়গায় অবস্থিত। একে অপরের সমান্তরালে জোড়া শৃঙ্খলে ডিএনএ অণুতে অবস্থিত নিউক্লিওটাইডগুলির কঠোর সঙ্গতিকে বলা হয় পরিপূরকতা (ঐচ্ছিক)। সঠিক প্রজনন বা প্রতিলিপি ডিএনএ অণুর এই বৈশিষ্ট্যের কারণে জেনেটিক তথ্য সঠিকভাবে সম্ভব।
ডিএনএ-তে, জৈবিক তথ্য এমনভাবে রেকর্ড করা হয় যাতে তা হুবহু কপি করে বংশধর কোষে প্রেরণ করা যায়। কোষ বিভাজনের আগে, প্রতিলিপি (স্ব-দ্বিগুণ ) ডিএনএ। যেহেতু প্রতিটি চেইনে একটি নিউক্লিওটাইড ক্রম থাকে যা অংশীদার চেইনের অনুক্রমের পরিপূরক, তারা আসলে একই জেনেটিক তথ্য বহন করে। আপনি যদি স্ট্র্যান্ডগুলিকে আলাদা করেন এবং একটি দ্বিতীয় স্ট্র্যান্ড তৈরি করতে তাদের প্রতিটিকে একটি টেমপ্লেট (ম্যাট্রিক্স) হিসাবে ব্যবহার করেন, আপনি দুটি নতুন অভিন্ন ডিএনএ স্ট্র্যান্ড পাবেন। এভাবেই কোষে DNA সদৃশ হয়।


